RUNX3 Promotes the Tumorigenic Phenotype in KGN, a Human Granulosa Cell Tumor-Derived Cell Line

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Expression of RUNX Proteins in GCT and Immortalized Granulosa Cell Lines
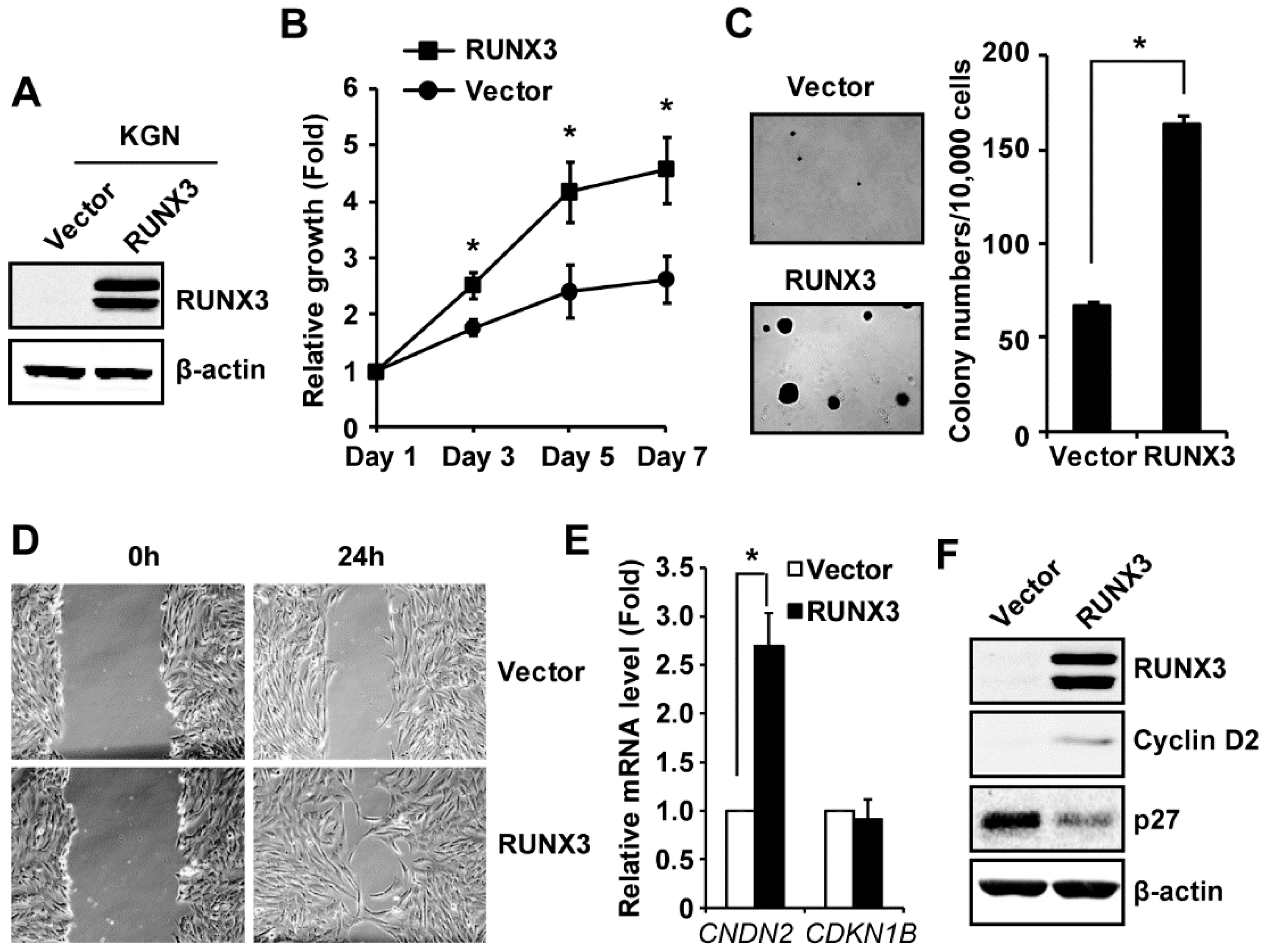
2.2. RUNX3 Promotes Cell Growth and Migration in KGN Cells
2.3. RUNX3 Regulates the Expression of Cyclin D2 and CDK Inhibitor p27Kip1 in KGN Cells
2.4. RUNX3 Enables KGN Cells to Form Subcutaneous Tumors
2.5. Overexpression of dnRUNX3 Decreases Growth of COV434 Cells
2.6. Expression of RUNX Proteins in Human GCT Tissues
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Gene Transfer
4.3. Neutral Red Uptake Assay of Cell Proliferation
4.4. Soft Agar Assay
4.5. Cell Migration Assay
4.6. Preparation of Whole Cell Lysates and Cytosolic/Nuclear Fraction and Immunoblotting
4.7. RNA Isolation and Quantitative Reverse Transcription-PCR (qRT-PCR)
4.8. Xenograft Model of KGN Cells
4.9. Expression of RUNX Proteins in GCT Tissues
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colombo, N.; Parma, G.; Zanagnolo, V.; Insinga, A. Management of ovarian stromal cell tumors. J. Clin. Oncol. 2007, 25, 2944–2951. [Google Scholar] [CrossRef] [PubMed]
- Sekkate, S.; Kairouani, M.; Serji, B.; M’Rabti, H.; El Ghissassi, I.; Errihani, H. Granulosa cell tumors of the ovary. Bull. Cancer 2014, 101, 93–101. [Google Scholar] [PubMed]
- Levin, G.; Zigron, R.; Haj-Yahya, R.; Matan, L.S.; Rottenstreich, A. Granulosa cell tumor of ovary: A systematic review of recent evidence. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 225, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, S.; Fuller, P.J. Molecular pathogenesis of granulosa cell tumors of the ovary. Endocr. Rev. 2012, 33, 109–144. [Google Scholar] [CrossRef] [PubMed]
- Mangili, G.; Ottolina, J.; Gadducci, A.; Giorda, G.; Breda, E.; Savarese, A.; Candiani, M.; Frigerio, L.; Scarfone, G.; Pignata, S.; et al. Long-term follow-up is crucial after treatment for granulosa cell tumours of the ovary. Br. J. Cancer 2013, 109, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkkila, A.; Haltia, U.M.; Tapper, J.; McConechy, M.K.; Huntsman, D.G.; Heikinheimo, M. Pathogenesis and treatment of adult-type granulosa cell tumor of the ovary. Ann. Med. 2017, 49, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.P.; Kobel, M.; Senz, J.; Morin, R.D.; Clarke, B.A.; Wiegand, K.C.; Leung, G.; Zayed, A.; Mehl, E.; Kalloger, S.E.; et al. Mutation of foxl2 in granulosa-cell tumors of the ovary. N. Engl. J. Med. 2009, 360, 2719–2729. [Google Scholar] [CrossRef] [PubMed]
- Rosario, R.; Cohen, P.A.; Shelling, A.N. The role of foxl2 in the pathogenesis of adult ovarian granulosa cell tumours. Gynecol. Oncol. 2014, 133, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Mancari, R.; Portuesi, R.; Colombo, N. Adult granulosa cell tumours of the ovary. Curr. Opin. Oncol. 2014, 26, 536–541. [Google Scholar] [CrossRef]
- Yanagida, S.; Anglesio, M.S.; Nazeran, T.M.; Lum, A.; Inoue, M.; Iida, Y.; Takano, H.; Nikaido, T.; Okamoto, A.; Huntsman, D.G. Clinical and genetic analysis of recurrent adult-type granulosa cell tumor of the ovary: Persistent preservation of heterozygous c.402c>g foxl2 mutation. PLoS ONE 2017, 12, e0178989. [Google Scholar] [CrossRef]
- Goyama, S.; Schibler, J.; Cunningham, L.; Zhang, Y.; Rao, Y.; Nishimoto, N.; Nakagawa, M.; Olsson, A.; Wunderlich, M.; Link, K.A.; et al. Transcription factor runx1 promotes survival of acute myeloid leukemia cells. J. Clin. Investig. 2013, 123, 3876–3888. [Google Scholar] [CrossRef] [PubMed]
- Akech, J.; Wixted, J.J.; Bedard, K.; van der Deen, M.; Hussain, S.; Guise, T.A.; van Wijnen, A.J.; Stein, J.L.; Languino, L.R.; Altieri, D.C.; et al. Runx2 association with progression of prostate cancer in patients: Mechanisms mediating bone osteolysis and osteoblastic metastatic lesions. Oncogene 2010, 29, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Qu, Z.; Ong, C.W.; Tsang, Y.H.; Xiao, G.; Shapiro, D.; Salto-Tellez, M.; Ito, K.; Ito, Y.; Chen, L.F. Runx3 acts as a tumor suppressor in breast cancer by targeting estrogen receptor alpha. Oncogene 2012, 31, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, E.; Inuzuka, M.; Maruyama, M.; Satake, M.; Naito-Fujimoto, M.; Ito, Y.; Shigesada, K. Molecular cloning and characterization of pebp2 beta, the heterodimeric partner of a novel drosophila runt-related DNA binding protein pebp2 alpha. Virology 1993, 194, 314–331. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Curry, T.E., Jr. Luteinizing hormone-induced runx1 regulates the expression of genes in granulosa cells of rat periovulatory follicles. Mol. Endocrinol. 2006, 20, 2156–2172. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Park, E.S.; Jo, M. Runt-related transcription factor 1 regulates luteinized hormone-induced prostaglandin-endoperoxide synthase 2 expression in rat periovulatory granulosa cells. Endocrinology 2009, 150, 3291–3300. [Google Scholar] [CrossRef] [PubMed]
- Park, E.S.; Lind, A.K.; Dahm-Kahler, P.; Brannstrom, M.; Carletti, M.Z.; Christenson, L.K.; Curry, T.E., Jr.; Jo, M. Runx2 transcription factor regulates gene expression in luteinizing granulosa cells of rat ovaries. Mol. Endocrinol. 2010, 24, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Park, E.S.; Park, J.; Franceschi, R.T.; Jo, M. The role for runt related transcription factor 2 (runx2) as a transcriptional repressor in luteinizing granulosa cells. Mol. Cell. Endocrinol. 2012, 362, 165–175. [Google Scholar] [CrossRef]
- Ojima, F.; Saito, Y.; Tsuchiya, Y.; Ogoshi, M.; Fukamachi, H.; Inagaki, K.; Otsuka, F.; Takeuchi, S.; Takahashi, S. Runx3 regulates folliculogenesis and steroidogenesis in granulosa cells of immature mice. Cell Tissue Res. 2019, 375, 743–754. [Google Scholar] [CrossRef]
- Ojima, F.; Saito, Y.; Tsuchiya, Y.; Kayo, D.; Taniuchi, S.; Ogoshi, M.; Fukamachi, H.; Takeuchi, S.; Takahashi, S. Runx3 transcription factor regulates ovarian functions and ovulation in female mice. J. Reprod. Dev. 2016, 62, 479–486. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.; Park, J.; Curry, T.E., Jr.; Mishra, B.; Gossen, J.; Taniuchi, I.; Jo, M. Core binding factor-beta knockdown alters ovarian gene expression and function in the mouse. Mol. Endocrinol. 2016, 30, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Lee-Thacker, S.; Choi, Y.; Taniuchi, I.; Takarada, T.; Yoneda, Y.; Ko, C.; Jo, M. Core binding factor beta expression in ovarian granulosa cells is essential for female fertility. Endocrinology 2018, 159, 2094–2109. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Bae, S.C.; Chuang, L.S. The runx family: Developmental regulators in cancer. Nat. Rev. Cancer 2015, 15, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Chuang, L.S.; Kimura, S.; Lai, S.K.; Ong, C.W.; Yan, B.; Salto-Tellez, M.; Choolani, M.; Ito, Y. Runx3 functions as an oncogene in ovarian cancer. Gynecol. Oncol. 2011, 122, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Keita, M.; Bachvarova, M.; Morin, C.; Plante, M.; Gregoire, J.; Renaud, M.C.; Sebastianelli, A.; Trinh, X.B.; Bachvarov, D. The runx1 transcription factor is expressed in serous epithelial ovarian carcinoma and contributes to cell proliferation, migration and invasion. Cell Cycle 2013, 12, 972–986. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Keita, M.; Bachvarova, M.; Gobeil, S.; Morin, C.; Plante, M.; Gregoire, J.; Renaud, M.C.; Sebastianelli, A.; Trinh, X.B.; et al. Inhibition of runx2 transcriptional activity blocks the proliferation, migration and invasion of epithelial ovarian carcinoma cells. PLoS ONE 2013, 8, e74384. [Google Scholar] [CrossRef] [PubMed]
- Nevadunsky, N.S.; Barbieri, J.S.; Kwong, J.; Merritt, M.A.; Welch, W.R.; Berkowitz, R.S.; Mok, S.C. Runx3 protein is overexpressed in human epithelial ovarian cancer. Gynecol. Oncol. 2009, 112, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Greer, A.H.; Yong, T.; Fennell, K.; Moustafa, Y.W.; Fowler, M.; Galiano, F.; Ng, S.W.; Berkowitz, R.S.; Cardelli, J.; Meyers, S.; et al. Knockdown of core binding factorbeta alters sphingolipid metabolism. J. Cell. Physiol. 2013, 228, 2350–2364. [Google Scholar] [CrossRef] [PubMed]
- Barghout, S.H.; Zepeda, N.; Vincent, K.; Azad, A.K.; Xu, Z.; Yang, C.; Steed, H.; Postovit, L.M.; Fu, Y. Runx3 contributes to carboplatin resistance in epithelial ovarian cancer cells. Gynecol. Oncol. 2015, 138, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, V.S.; Shahid, M.; Husain, S.A. Cpg methylation of the fhit, fancf, cyclin-d2, brca2 and runx3 genes in granulosa cell tumors (gcts) of ovarian origin. Mol. Cancer 2004, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Salto-Tellez, M.; Peh, B.K.; Ito, K.; Tan, S.H.; Chong, P.Y.; Han, H.C.; Tada, K.; Ong, W.Y.; Soong, R.; Voon, D.C.; et al. Runx3 protein is overexpressed in human basal cell carcinomas. Oncogene 2006, 25, 7646–7649. [Google Scholar] [CrossRef] [PubMed]
- Anttonen, M.; Pihlajoki, M.; Andersson, N.; Georges, A.; L’Hote, D.; Vattulainen, S.; Farkkila, A.; Unkila-Kallio, L.; Veitia, R.A.; Heikinheimo, M. Foxl2, gata4, and smad3 co-operatively modulate gene expression, cell viability and apoptosis in ovarian granulosa cell tumor cells. PLoS ONE 2014, 9, e85545. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Chang, H.M.; Qiu, X.; Fang, L.; Leung, P.C. Foxl2-induced follistatin attenuates activin a-stimulated cell proliferation in human granulosa cell tumors. Biochem. Biophys. Res. Commun. 2014, 443, 537–542. [Google Scholar] [CrossRef]
- Wang, C.; Lv, X.; Jiang, C.; Cordes, C.M.; Fu, L.; Lele, S.M.; Davis, J.S. Transforming growth factor alpha (tgfalpha) regulates granulosa cell tumor (gct) cell proliferation and migration through activation of multiple pathways. PLoS ONE 2012, 7, e48299. [Google Scholar]
- Robker, R.L.; Richards, J.S. Hormone-induced proliferation and differentiation of granulosa cells: A coordinated balance of the cell cycle regulators cyclin d2 and p27kip1. Mol. Endocrinol. 1998, 12, 924–940. [Google Scholar] [CrossRef] [PubMed]
- Imai, M.; Muraki, M.; Takamatsu, K.; Saito, H.; Seiki, M.; Takahashi, Y. Spontaneous transformation of human granulosa cell tumours into an aggressive phenotype: A metastasis model cell line. BMC Cancer 2008, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, Y.H.; Kim, H.M.; Park, H.O.; Ha, N.C.; Kim, T.H.; Park, M.; Lee, K.; Bae, J. Foxl2 posttranslational modifications mediated by gsk3beta determine the growth of granulosa cell tumours. Nat. Commun. 2014, 5, 2936. [Google Scholar] [CrossRef]
- Bilandzic, M.; Wang, Y.; Ahmed, N.; Luwor, R.B.; Zhu, H.J.; Findlay, J.K.; Stenvers, K.L. Betaglycan blocks metastatic behaviors in human granulosa cell tumors by suppressing nfkappab-mediated induction of mmp2. Cancer Lett. 2014, 354, 107–114. [Google Scholar] [CrossRef]
- Goh, Y.M.; Cinghu, S.; Hong, E.T.; Lee, Y.S.; Kim, J.H.; Jang, J.W.; Li, Y.H.; Chi, X.Z.; Lee, K.S.; Wee, H.; et al. Src kinase phosphorylates RUNX3 at tyrosine residues and localizes the protein in the cytoplasm. J. Biol. Chem. 2010, 285, 10122–10129. [Google Scholar] [CrossRef]
- Chi, X.Z.; Kim, J.; Lee, Y.H.; Lee, J.W.; Lee, K.S.; Wee, H.; Kim, W.J.; Park, W.Y.; Oh, B.C.; Stein, G.S.; et al. Runt-related transcription factor RUNX3 is a target of MDM2-mediated ubiquitination. Cancer Res. 2009, 69, 8111–8119. [Google Scholar] [CrossRef]
- Jin, Y.H.; Jeon, E.J.; Li, Q.L.; Lee, Y.H.; Choi, J.K.; Kim, W.J.; Lee, K.Y.; Bae, S.C. Transforming growth factor-beta stimulates p300-dependent RUNX3 acetylation, which inhibits ubiquitination-mediated degradation. J. Biol. Chem. 2004, 279, 29409–29417. [Google Scholar] [CrossRef] [PubMed]
- Hnit, S.S.; Xie, C.; Yao, M.; Holst, J.; Bensoussan, A.; De Souza, P.; Li, Z.; Dong, Q. P27(kip1) signaling: Transcriptional and post-translational regulation. Int. J. Biochem. Cell Biol. 2015, 68, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Haltia, U.M.; Andersson, N.; Yadav, B.; Farkkila, A.; Kulesskiy, E.; Kankainen, M.; Tang, J.; Butzow, R.; Riska, A.; Leminen, A.; et al. Systematic drug sensitivity testing reveals synergistic growth inhibition by dasatinib or mtor inhibitors with paclitaxel in ovarian granulosa cell tumor cells. Gynecol. Oncol. 2017, 144, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Chan-Penebre, E.; Armstrong, K.; Drew, A.; Grassian, A.R.; Feldman, I.; Knutson, S.K.; Kuplast-Barr, K.; Roche, M.; Campbell, J.; Ho, P.; et al. Selective killing of smarca2- and smarca4-deficient small cell carcinoma of the ovary, hypercalcemic type cells by inhibition of ezh2: In vitro and in vivo preclinical models. Mol. Cancer Therapeutics 2017, 16, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Carlton, A.L.; Illendula, A.; Gao, Y.; Llaneza, D.C.; Boulton, A.; Shah, A.; Rajewski, R.A.; Landen, C.N.; Wotton, D.; Bushweller, J.H. Small molecule inhibition of the cbfbeta/runx interaction decreases ovarian cancer growth and migration through alterations in genes related to epithelial-to-mesenchymal transition. Gynecol. Oncol. 2018, 149, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Nishi, Y.; Yanase, T.; Mu, Y.; Oba, K.; Ichino, I.; Saito, M.; Nomura, M.; Mukasa, C.; Okabe, T.; Goto, K.; et al. Establishment and characterization of a steroidogenic human granulosa-like tumor cell line, kgn, that expresses functional follicle-stimulating hormone receptor. Endocrinology 2001, 142, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Vollmer, M.; De Geyter, M.; Litzistorf, Y.; Ladewig, A.; Durrenberger, M.; Guggenheim, R.; Miny, P.; Holzgreve, W.; De Geyter, C. Characterization of an immortalized human granulosa cell line (cov434). Mol. Hum. Reproduct. 2000, 6, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Liu, Q.; Salto-Tellez, M.; Yano, T.; Tada, K.; Ida, H.; Huang, C.; Shah, N.; Inoue, M.; Rajnakova, A.; et al. Runx3, a novel tumor suppressor, is frequently inactivated in gastric cancer by protein mislocalization. Cancer Res. 2005, 65, 7743–7750. [Google Scholar] [CrossRef]
- Yano, T.; Ito, K.; Fukamachi, H.; Chi, X.Z.; Wee, H.J.; Inoue, K.; Ida, H.; Bouillet, P.; Strasser, A.; Bae, S.C.; et al. The runx3 tumor suppressor upregulates bim in gastric epithelial cells undergoing transforming growth factor beta-induced apoptosis. Mol. Cell. Biol. 2006, 26, 4474–4488. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Steed, H.; Davidge, S.; Fu, Y. Tgfbeta and egf synergistically induce a more invasive phenotype of epithelial ovarian cancer cells. Biochem. Biophys. Res. Commun. 2010, 401, 376–381. [Google Scholar] [CrossRef]
- Fu, Y.; Sies, H.; Lei, X.G. Opposite roles of selenium-dependent glutathione peroxidase-1 in superoxide generator diquat- and peroxynitrite-induced apoptosis and signaling. J. Biol. Chem. 2001, 276, 43004–43009. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.K.; Chakrabarti, S.; Xu, Z.; Davidge, S.T.; Fu, Y. Coiled-coil domain containing 3 (ccdc3) represses tumor necrosis factor-alpha/nuclear factor kappab-induced endothelial inflammation. Cell. Signall. 2014, 26, 2793–2800. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Crosley, P.; Azad, A.K.; Gupta, N.; Gokul, N.; Xu, Z.; Weinfeld, M.; Postovit, L.-M.; Pangas, S.A.; Hitt, M.M.; et al. RUNX3 Promotes the Tumorigenic Phenotype in KGN, a Human Granulosa Cell Tumor-Derived Cell Line. Int. J. Mol. Sci. 2019, 20, 3471. https://doi.org/10.3390/ijms20143471
Chen H, Crosley P, Azad AK, Gupta N, Gokul N, Xu Z, Weinfeld M, Postovit L-M, Pangas SA, Hitt MM, et al. RUNX3 Promotes the Tumorigenic Phenotype in KGN, a Human Granulosa Cell Tumor-Derived Cell Line. International Journal of Molecular Sciences. 2019; 20(14):3471. https://doi.org/10.3390/ijms20143471
Chicago/Turabian StyleChen, Huachen, Powel Crosley, Abul K. Azad, Nidhi Gupta, Nisha Gokul, Zhihua Xu, Michael Weinfeld, Lynne-Marie Postovit, Stephanie A. Pangas, Mary M. Hitt, and et al. 2019. "RUNX3 Promotes the Tumorigenic Phenotype in KGN, a Human Granulosa Cell Tumor-Derived Cell Line" International Journal of Molecular Sciences 20, no. 14: 3471. https://doi.org/10.3390/ijms20143471
APA StyleChen, H., Crosley, P., Azad, A. K., Gupta, N., Gokul, N., Xu, Z., Weinfeld, M., Postovit, L. -M., Pangas, S. A., Hitt, M. M., & Fu, Y. (2019). RUNX3 Promotes the Tumorigenic Phenotype in KGN, a Human Granulosa Cell Tumor-Derived Cell Line. International Journal of Molecular Sciences, 20(14), 3471. https://doi.org/10.3390/ijms20143471