H2O2-Based Method for Rapid Detection of Transgene-Free Rice Plants from Segregating CRISPR/Cas9 Genome-Edited Progenies

 , and
, and 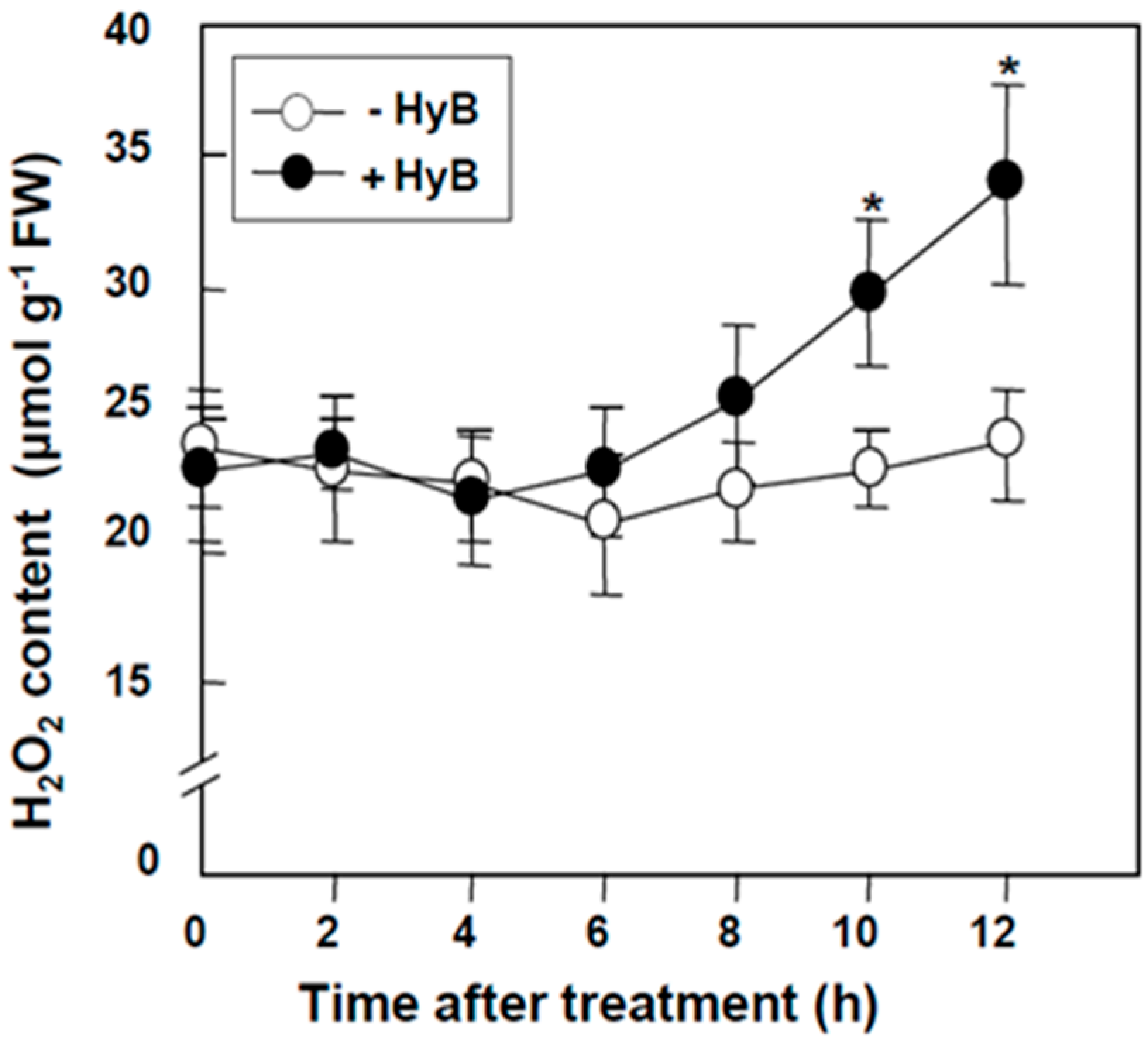{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Hygromycin (HyB) Significantly Increased the Production of H2O2 in Leaves of Rice Seedlings
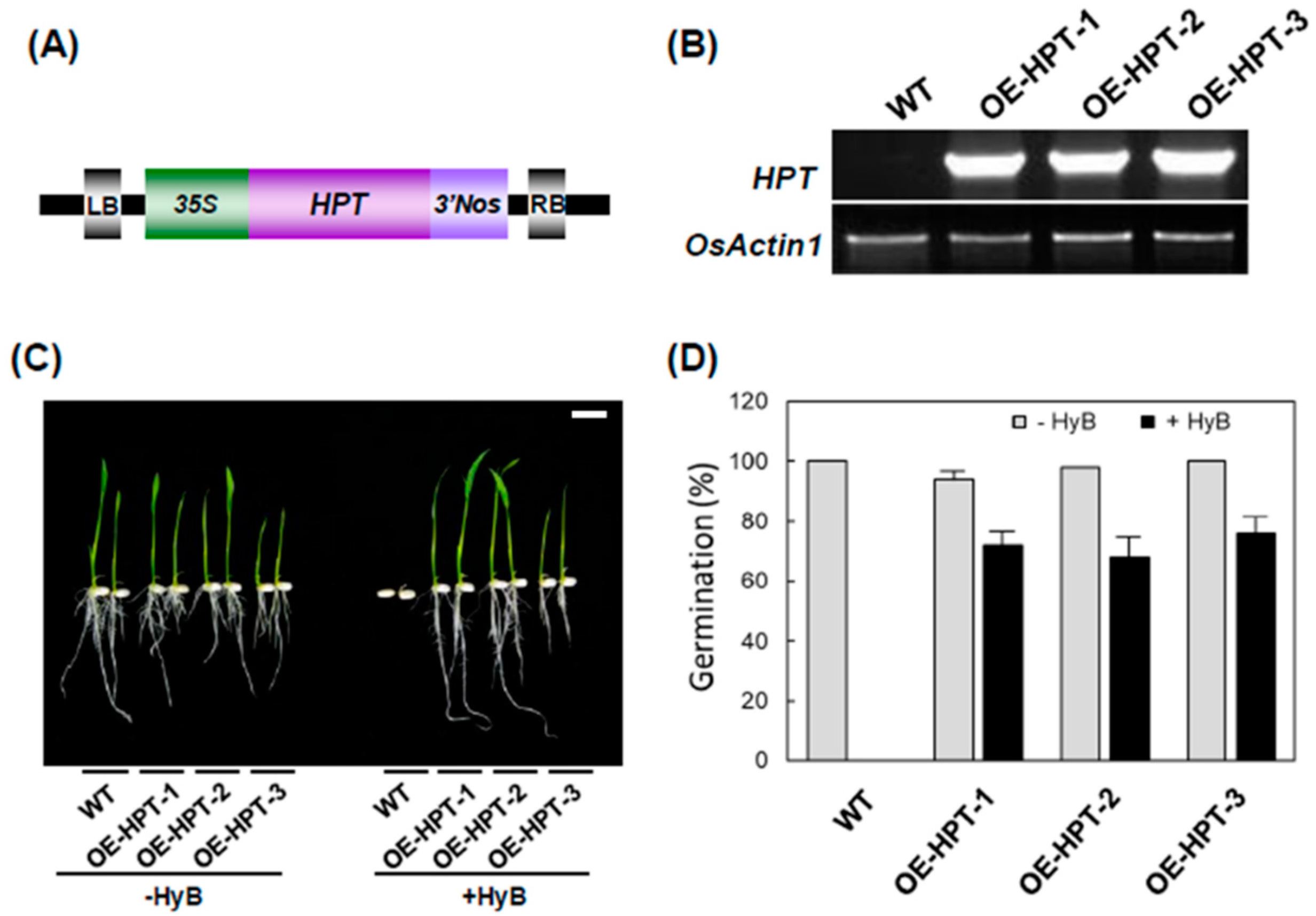
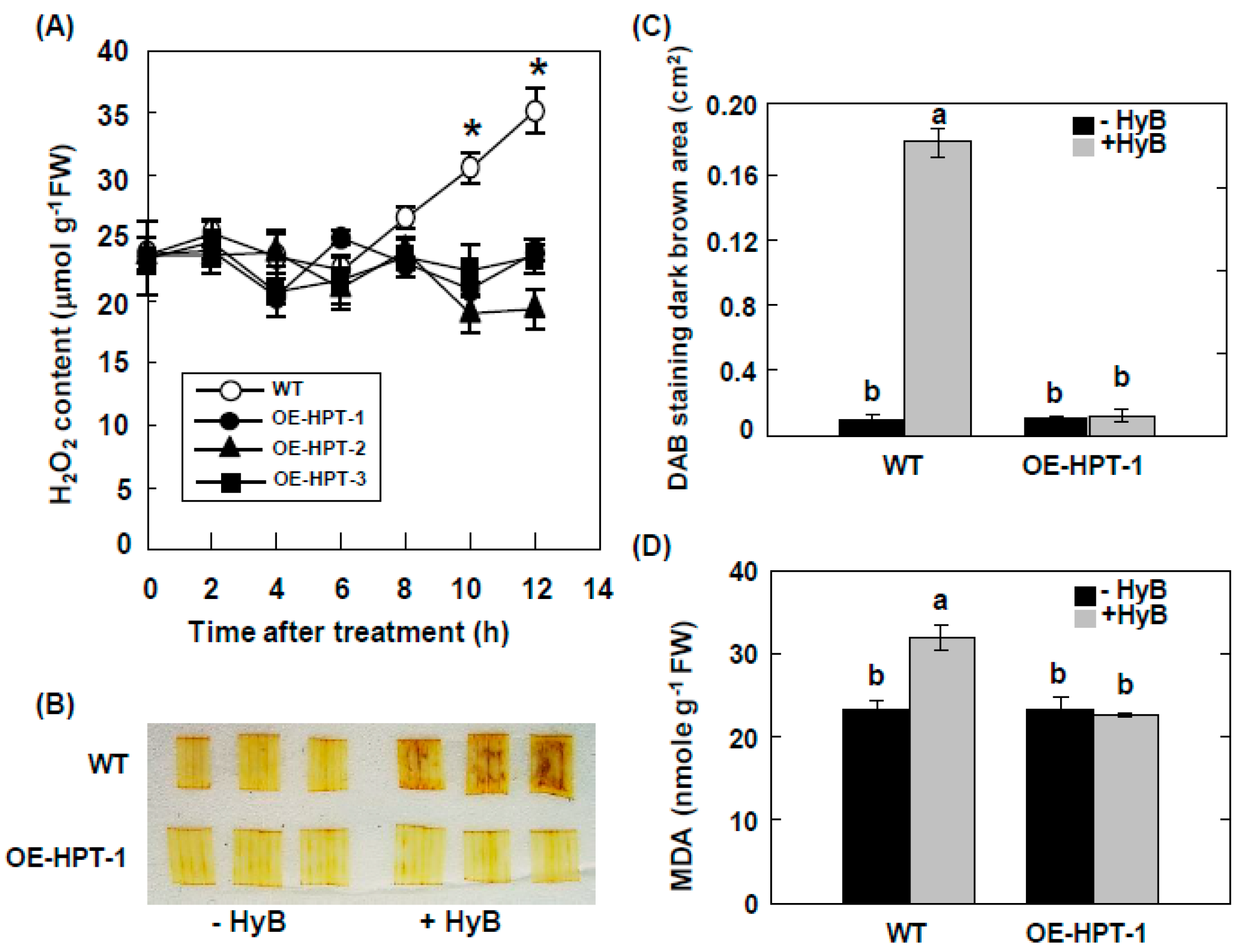
2.2. Accumulation of H2O2 in Transgenic Rice Overexpressing HPT
2.3. DAB–H2O2 Method is Efficient for Detecting HPT-Containing Transgenic Rice
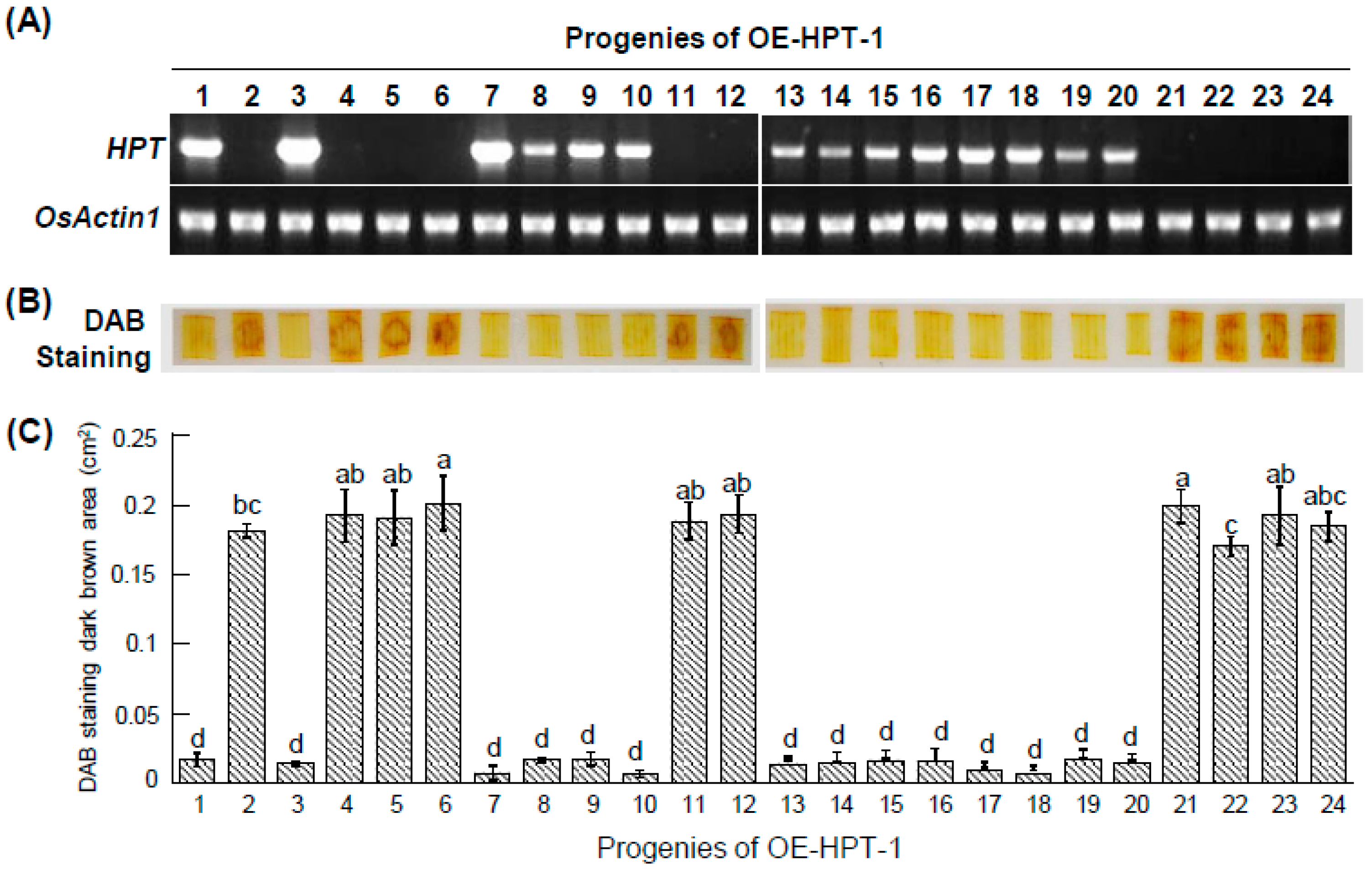
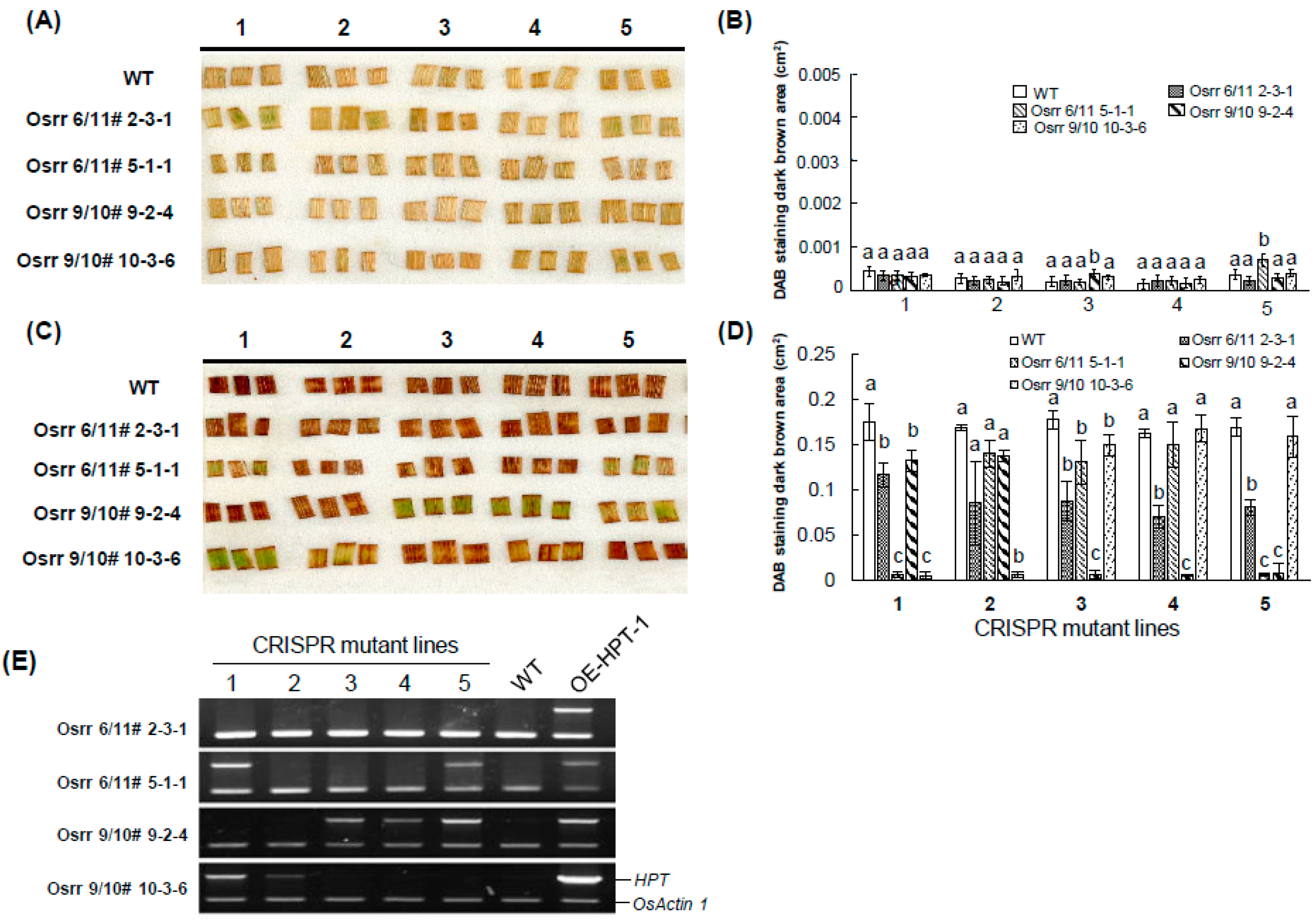
2.4. Detection of Transgene-Free Genome-Edited Plants by DAB Method
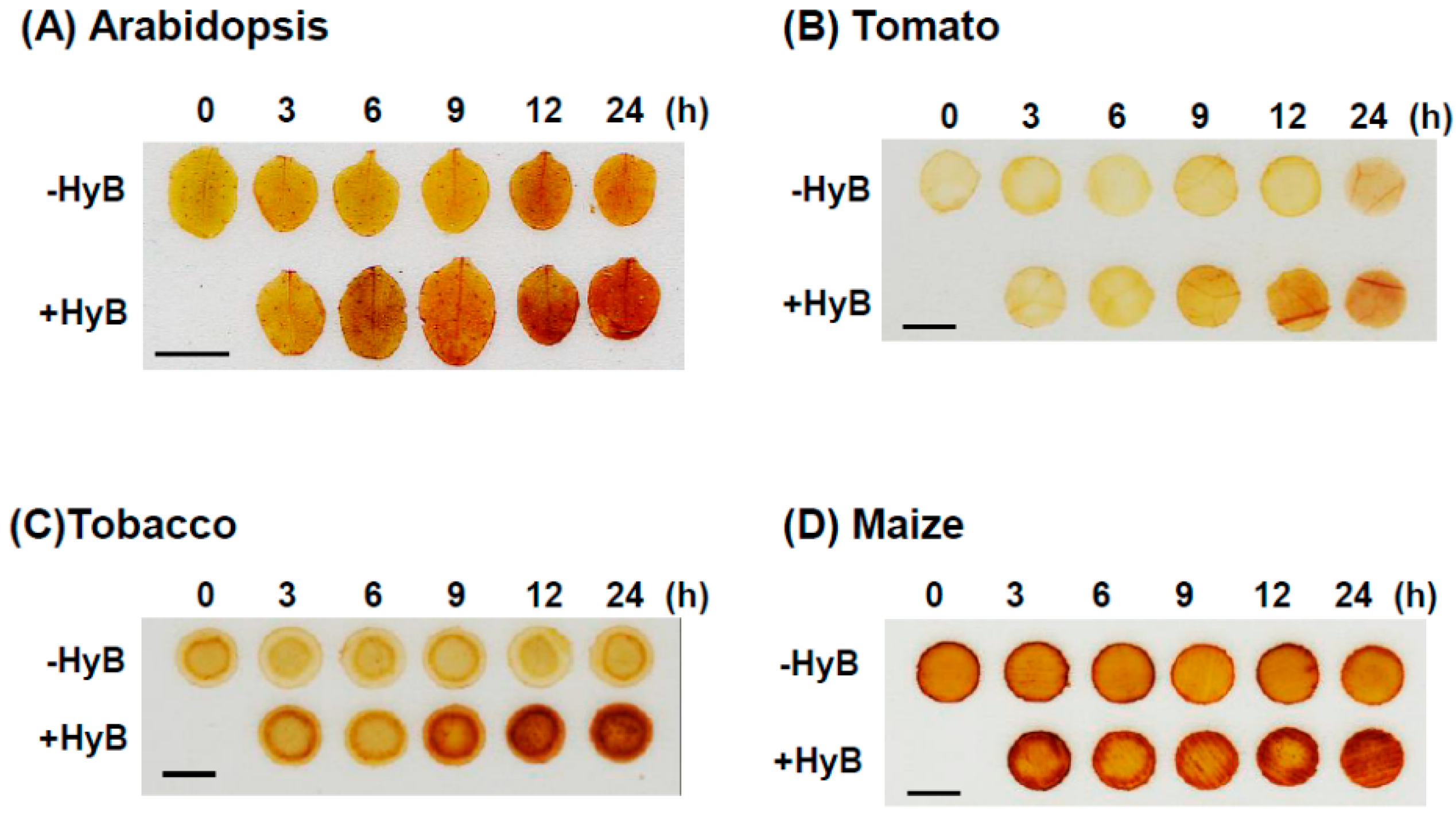
2.5. HyB-Induced Overproduction of H2O2 Observed in Monocot and Dicot Plants
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Generation of Hygromycinb (HyB)-Resistant Transgenic Rice
4.3. HyB Treatment
4.4. Visual Detection of H2O2
4.5. Quantification of H2O2, DAB–H2O2, Malondialdehyde, and Protein Content
4.6. RT-PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| CRISPR | Clustered Regularly Interspaced Palindromic Repeat |
| DAB | 3,3′-diaminobenzidine |
| HPT | Hygromycin phosphotransferase |
| HyB | Hygromycin B |
| WT | Wild-type |
References
- Van den Elzen, P.J.; Townsend, J.; Lee, K.Y.; Bedbrook, J.R. A chimaeric hygromycin resistance gene as a selectable marker in plant cells. Plant Mol. Biol. 1985, 5, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Pardo, J.M.; Malpartida, F.; Rico, M.; Jimenez, A. Biochemical basis of resistance to hygromycin B in Streptomyces hygroscopicus—The producing organism. J. Gen. Microbiol. 1985, 131, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Aldemita, R.R.; Hodges, T.K. Agrobacterium tumefaciens—Mediated transformation of japonica and indica rice varieties. Planta 1996, 199, 612–617. [Google Scholar] [CrossRef]
- Christou, P.; Ford, T.L. The impact of selection parameters on the phenotype and genotype of transgenic rice callus and plants. Transgenic Res. 1995, 4, 44–51. [Google Scholar] [CrossRef]
- Miki, B.; McHugh, S. Selectable marker genes in transgenic plants: Applications, alternatives and biosafety. J. Biotechnol. 2004, 107, 193–232. [Google Scholar] [CrossRef] [PubMed]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.P.A.; Reggiardo, M.I.; Ravizzini, R.A.; Altabe, S.G.; Cervigni, G.D.L.; Spitteler, M.A.; Morata, M.M.; Elias, F.E.; Vallejos, R.H. Hygromycin resistance as an efficient selectable marker for wheat stable transformation. Plant Cell Rep. 1996, 15, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Shavrukov, Y.; Bazanova, N.; Chirkova, L.; Borisjuk, N.; Kovalchuk, N.; Ismagul, A.; Parent, B.; Langridge, P.; Hrmova, M.; et al. Constitutive overexpression of the TaNF-YB4 gene in transgenic wheat significantly improves grain yield. J. Exp. Bot. 2015, 66, 6635–6650. [Google Scholar] [CrossRef]
- Trabucco, G.M.; Matos, D.A.; Lee, S.J.; Saathoff, A.J.; Priest, H.D.; Mockler, T.C.; Sarath, G.; Hazen, S.P. Functional characterization of cinnamyl alcohol dehydrogenase and caffeic acid O-methyltransferase in Brachypodium distachyon. BMC Biotechnol. 2013, 13, 61. [Google Scholar] [CrossRef]
- Vogel, J.; Hill, T. High-efficiency Agrobacterium-mediated transformation of Brachypodium distachyon inbred line Bd21-3. Plant Cell Rep. 2008, 27, 471–478. [Google Scholar] [CrossRef]
- Bartlett, J.G.; Alves, S.C.; Smedley, M.; Snape, J.W.; Harwood, W.A. High-throughput Agrobacterium-mediated barley transformation. Plant Methods 2008, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Hagio, T.; Hirabayashi, T.; Machii, H.; Tomotsune, H. Production of fertile transgenic barley (Hordeum vulgare L.) plant using the hygromycin-resistance marker. Plant Cell Rep. 1995, 14, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Soltesz, A.; Smedley, M.; Vashegyi, I.; Galiba, G.; Harwood, W.; Vagujfalvi, A. Transgenic barley lines prove the involvement of TaCBF14 and TaCBF15 in the cold acclimation process and in frost tolerance. J. Exp. Bot. 2013, 64, 1849–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettinger, R.C.; Wolfe, E.N.; Hoehn, M.M.; Marks, P.N.; Daily, W.A.; McGuire, J.M. Hygromycin. I. Preliminary studies on the production and biologic activity of a new antibiotic. Antibiot. Chemother. 1953, 3, 1268–1278. [Google Scholar]
- Cabañas, M.J.; Vázquez, D.; Modolell, J. Dual interference of hygromycin B with ribosomal translocation and with aminoacyl-tRNA recognition. Eur. J. Biochem. 1978, 87, 21–27. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Jiménez, A.; Vázquez, D.; Davies, J.E.; Schindler, D. Studies on the mode of action of hygromycin B, an inhibitor of translocation in eukaryotes. Biochim. Biophys. Acta 1978, 521, 459–469. [Google Scholar] [CrossRef]
- Dekeyser, R.; Claes, B.; Marichal, M.; Van Montagu, M.; Caplan, A. Evaluation of selectable markers for rice transformation. Plant Physiol. 1989, 90, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Song, W.J.; Ding, X. Effects of hygromycin on growth and development of Arabidopsis seedlings roots. Pak. J. Bot. 2007, 39, 2167–2173. [Google Scholar]
- Walters, D.A.; Vetsch, C.S.; Potts, D.E.; Lundquist, R.C. Transformation and inheritance of a hygromycin phosphotransferase gene in maize plants. Plant Mol. Biol. 1992, 18, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.-T.; Chang, H.-H.; Ho, S.-L.; Tong, W.-F.; Yu, S.-M. Agrobacterium-mediated production of transgenic rice plants expressing a chimeric α-amylase promoter/β-glucuronidase gene. Plant Mol. Biol. 1993, 22, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.Y.; Cheng, K.J.; Tseng, T.H.; Wang, C.S.; Liu, L.F.; Yu, S.M. Production of two highly active bacterial phytases with broad pH optima in germinated transgenic rice seeds. Transgenic Res. 2004, 13, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.H.; Do Choi, Y.; Kim, M.; Reuzeau, C.; Kim, J.K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Schaffrath, U.; Mauch, F.; Freydl, E.; Schweizer, P.; Dudler, R. Constitutive expression of the defense-related Rir1b gene in transgenic rice plants confers enhanced resistance to the rice blast fungus Magnaporthe grisea. Plant Mol. Biol. 2000, 43, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Y.; Wang, A.M.; Li, W.; Wang, Z.Y.; Cai, X.L. Establishing a gene trap system mediated by T-DNA(GUS) in rice. J. Integr. Plant Biol. 2008, 50, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Xu, Q.; Liu, Y.; Zhang, J.; Ren, D.; Wang, G.; Liu, Y. Generation of transgene-free maize male sterile lines using the CRISPR/Cas9 system. Front. Plant Sci. 2018, 9, 1180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An effective strategy for reliably isolating heritable and Cas9-free Arabidopsis mutants generated by CRISPR/Cas9-mediated genome editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef]
- Bibi, N.; Fan, K.; Yuan, S.; Ni, M.; Ahmed, I.M.; Malik, W.; Wang, X. An efficient and highly reproducible approach for the selection of upland transgenic cotton produced by pollen tube pathway method. Aust. J. Crop Sci. 2013, 7, 1714–1722. [Google Scholar]
- Kim, J.K.; Duan, X.; Wu, R.; Seok, S.J.; Boston, R.S.; Jang, I.C.; Eun, M.Y.; Nahm, B.H. Molecular and genetic analysis of transgenic rice plants expressing the ribosome-inactivating protein b-32 gene and the herbicide resistance bar gene. Mol. Breed. 1999, 5, 85–94. [Google Scholar] [CrossRef]
- Rajasekaran, K.; Majumdar, R.; Sickler, C.; Wei, Q.; Cary, J.; Bhatnagar, D. Fidelity of a simple Liberty leaf-painting assay to validate transgenic maize plants expressing the selectable marker gene, bar. J. Crop Improv. 2017, 31, 628–636. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Oung, H.-M.; Lin, K.-C.; Wu, T.-M.; Chandrika, N.N.P.; Hong, C.-Y. Hygromycin B-induced cell death is partly mediated by reactive oxygen species in rice (Oryza sativa L.). Plant Mol. Biol. 2015, 89, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-C.; Hou, Y.-W.; Wu, H.-Y.; Tsai, Y.-C. Efficient CRISPR/Cas9-mediated genome editing in rice using an early flowering cultivar Kitaake combined with simple mutant screening approaches. Crop Environ. Bioinform. 2017, 14, 175–186. [Google Scholar]
- Lee, H.Y.; Chen, Z.; Zhang, C.; Yoon, G.M. Editing of the OsACS locus alters phosphate deficiency-induced adaptive responses in rice seedlings. J. Exp. Bot. 2019, 70, 1927–1940. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-C.; Lin, T.-C.; Kieber, J.J.; Tsai, Y.-C. Response Regulator 9 and 10 negatively regulate salinity tolerance in rice. Plant Cell Physiol. 2019. accepted. [Google Scholar] [CrossRef]
- Zhu, X.; Xu, Y.; Yu, S.; Lu, L.; Ding, M.; Cheng, J.; Song, G.; Gao, X.; Yao, L.; Fan, D.; et al. An efficient genotyping method for genome-modified animals and human cells generated with CRISPR/Cas9 system. Sci. Rep. 2014, 4, 6420. [Google Scholar] [CrossRef]
- Dahlem, T.J.; Hoshijima, K.; Jurynec, M.J.; Gunther, D.; Starker, C.G.; Locke, A.S.; Weis, A.M.; Voytas, D.F.; Grunwald, D.J. Simple methods for generating and detecting locus-specific mutations induced with TALENs in the zebrafish genome. PLoS Genet. 2012, 8, e1002861. [Google Scholar] [CrossRef]
- Sanger, F.; Coulson, A.R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol. 1975, 94, 441–448. [Google Scholar] [CrossRef]
- Orozco-Cardenas, M.; Ryan, C.A. Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6553–6557. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.-C.; Hong, C.-Y.; Liu, L.-F.; Kao, C.H. Relative importance of Na+ and Cl− in NaCl− induced antioxidant systems in roots of rice seedlings. Physiol. Plant. 2004, 122, 86–94. [Google Scholar] [CrossRef]
- Graham, R.C., Jr.; Karnovsky, M.J. The early stages of absorption of injected horseradish peroxidase in the proximal tubules of mouse kidney: Ultrastructural cytochemistry by a new technique. J. Histochem. Cytochem. 1966, 14, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.-M.; Huang, J.-Z.; Oung, H.-M.; Hsu, Y.-T.; Tsai, Y.-C.; Hong, C.-Y. H2O2-Based Method for Rapid Detection of Transgene-Free Rice Plants from Segregating CRISPR/Cas9 Genome-Edited Progenies. Int. J. Mol. Sci. 2019, 20, 3885. https://doi.org/10.3390/ijms20163885
Wu T-M, Huang J-Z, Oung H-M, Hsu Y-T, Tsai Y-C, Hong C-Y. H2O2-Based Method for Rapid Detection of Transgene-Free Rice Plants from Segregating CRISPR/Cas9 Genome-Edited Progenies. International Journal of Molecular Sciences. 2019; 20(16):3885. https://doi.org/10.3390/ijms20163885
Chicago/Turabian StyleWu, Tsung-Meng, Jian-Zhi Huang, Hui-Min Oung, Yi-Ting Hsu, Yu-Chang Tsai, and Chwan-Yang Hong. 2019. "H2O2-Based Method for Rapid Detection of Transgene-Free Rice Plants from Segregating CRISPR/Cas9 Genome-Edited Progenies" International Journal of Molecular Sciences 20, no. 16: 3885. https://doi.org/10.3390/ijms20163885
APA StyleWu, T. -M., Huang, J. -Z., Oung, H. -M., Hsu, Y. -T., Tsai, Y. -C., & Hong, C. -Y. (2019). H2O2-Based Method for Rapid Detection of Transgene-Free Rice Plants from Segregating CRISPR/Cas9 Genome-Edited Progenies. International Journal of Molecular Sciences, 20(16), 3885. https://doi.org/10.3390/ijms20163885