Circulating miRNAs as Potential Biomarkers Associated with Cardiac Remodeling and Fibrosis in Chagas Disease Cardiomyopathy

 ,
,
Abstract
:1. Introduction
2. Results
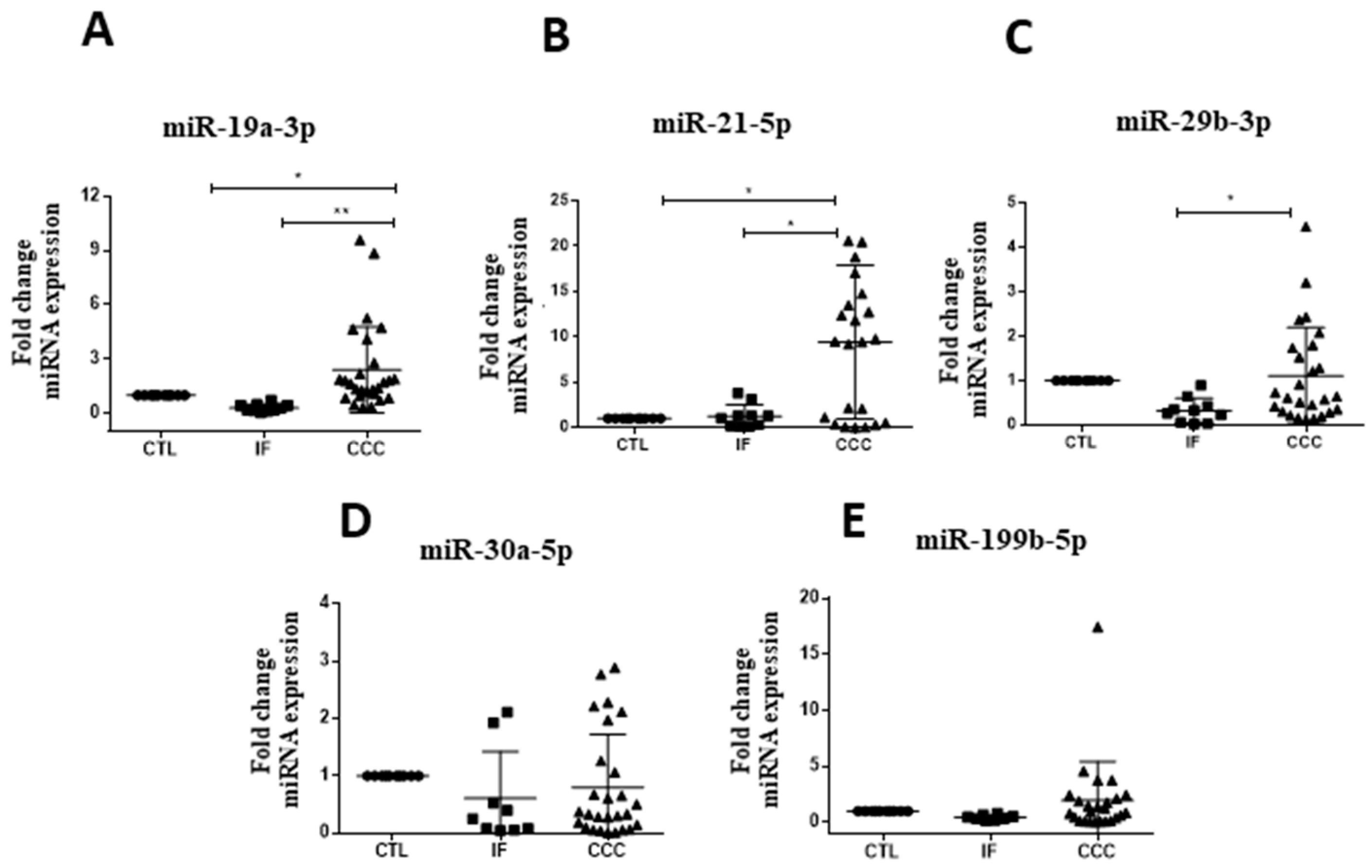
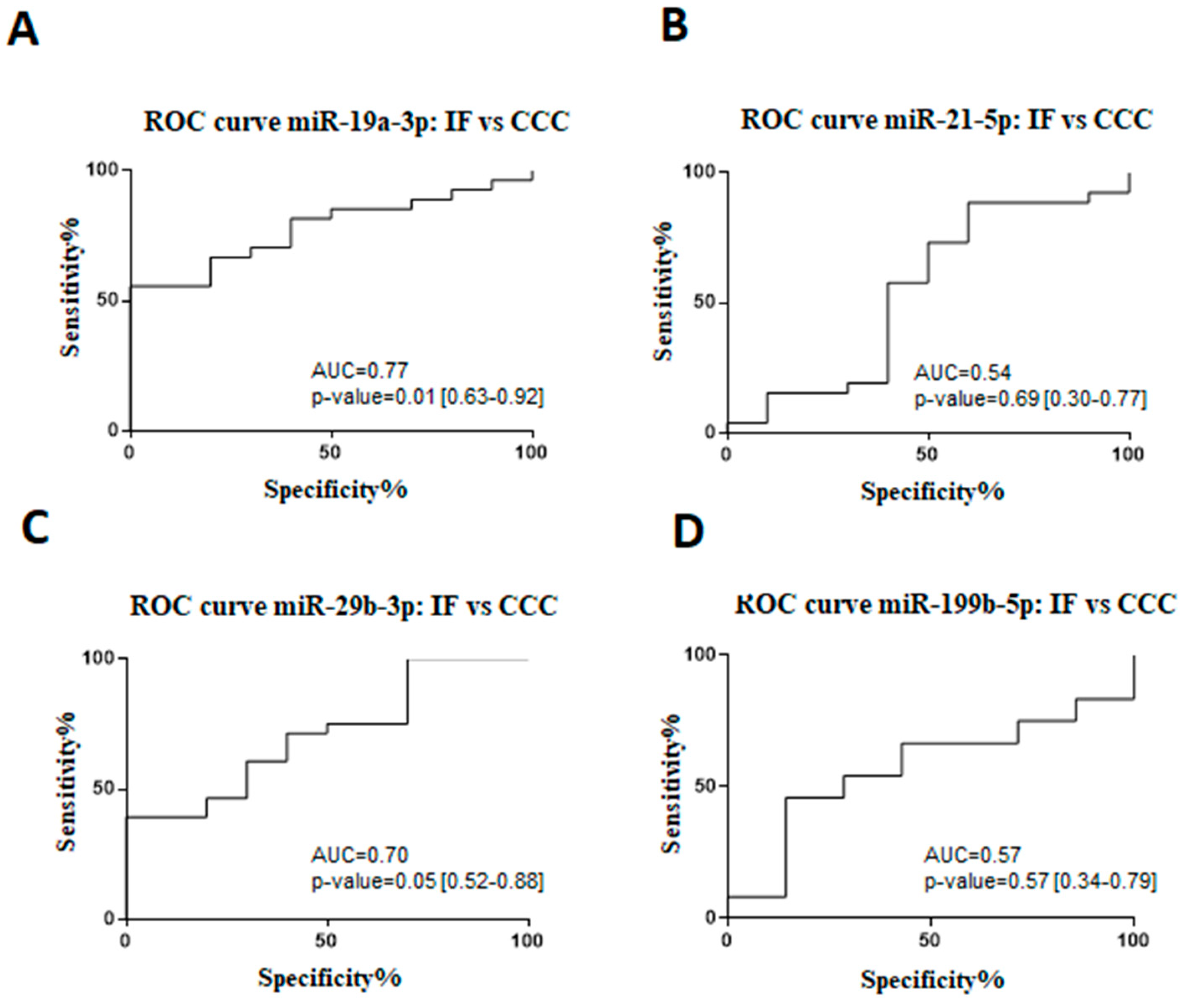
2.1. Expression Profile of Circulating microRNAs in Subjects with Chronic Chagas Disease
2.2. MicroRNAs in Cardiac Tissue Samples of Subjects with End-Stage CCC
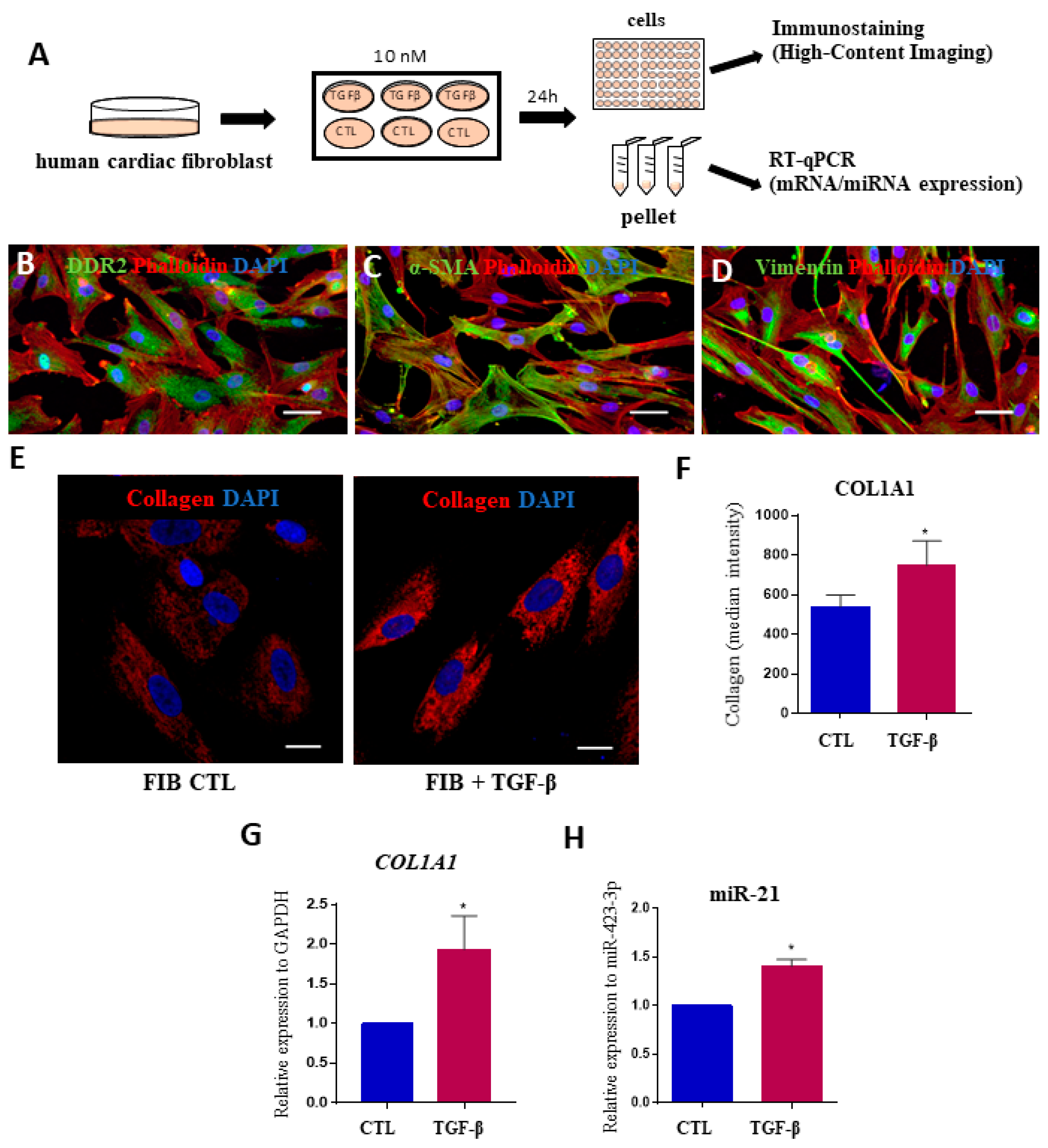
2.3. MicroRNAs Expression Levels in Fibrosis Using Human Cardiac Fibroblasts
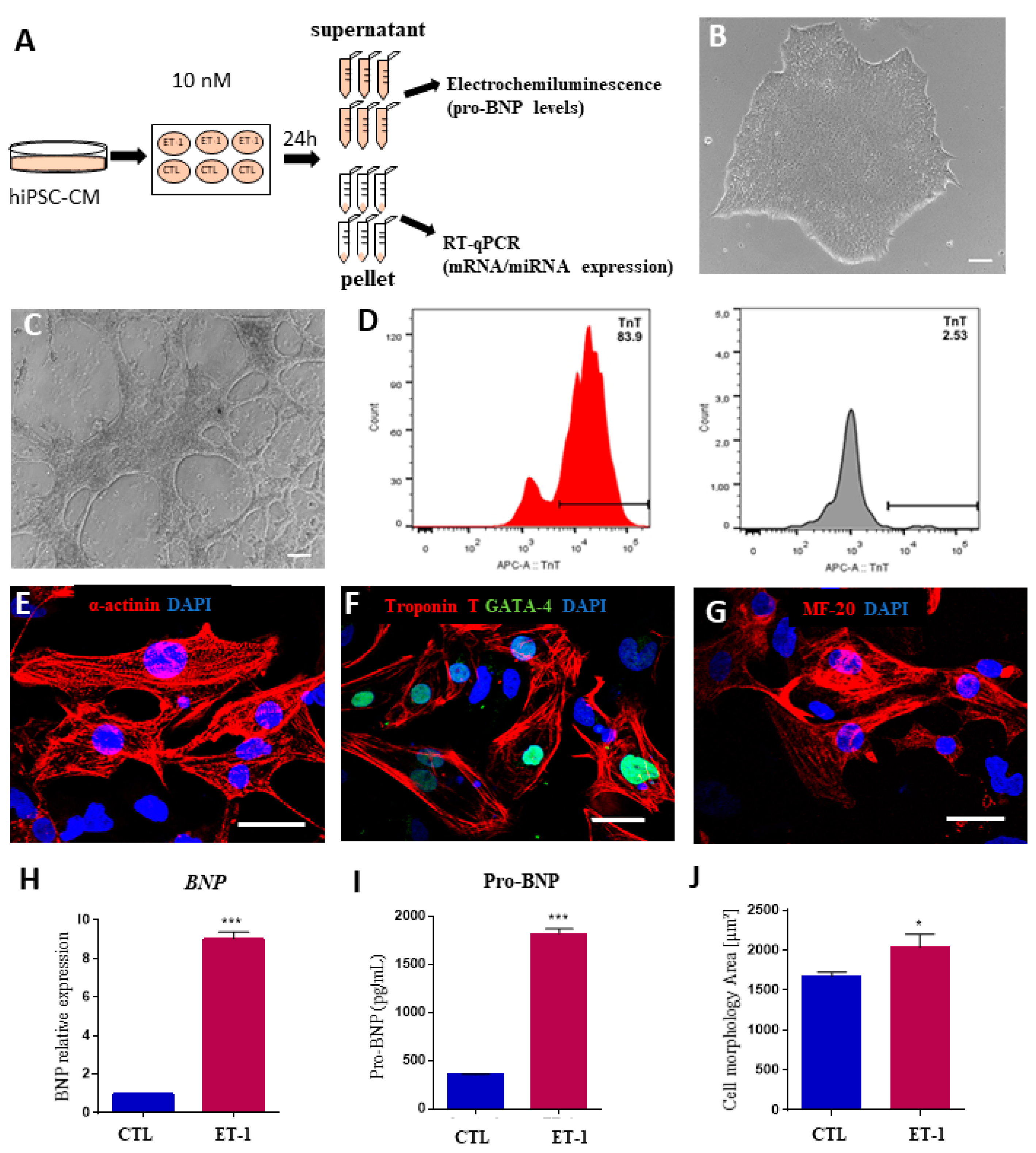
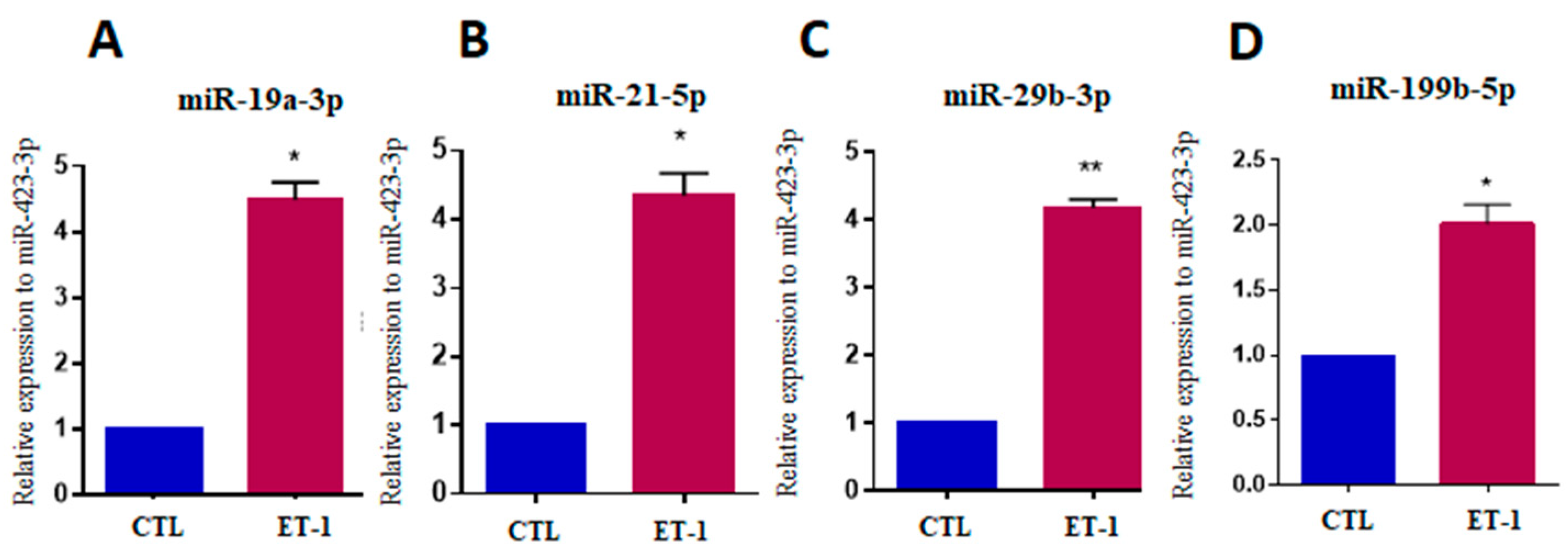
2.4. Expression of MicroRNAs in a Hypertrophy Model Using hiPSC-Derived Cardiomyocytes
3. Discussion
4. Materials and Methods
4.1. Ethics and Study Design
4.2. Study Population
4.3. MiRNA Selection
4.4. Total RNA, MicroRNA Extraction and cDNA Synthesis
4.5. Quantitative Real-time PCR (RT-qPCR)
4.6. Functional in vitro Assays with Human Cardiac Fibroblasts
4.7. Modeling Human Cardiac Hypertrophy in hiPSC-CM
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- WHO. Epidemiology; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- Bern, C. Chagas’ Disease. N. Engl. J. Med. 2015, 373, 1881–1882. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.D.A., Jr.; Andrade, Z.A. Identificação do Trypanosoma cruzi nos tecidos extracardíacos de portadores de miocardite crônica chagásica. Rev. Soc. Bras. Med. Trop. 1984, 17, 123–126. [Google Scholar] [CrossRef]
- Soares, M.B.; Pontes-de-Carvalho, L.; Ribeiro-dos-Santos, R. The pathogenesis of Chagas’ disease: When autoimmune and parasite-specific immune responses meet. An. Acad. Bras Cienc. 2001, 73, 547–559. [Google Scholar] [CrossRef] [PubMed]
- De Lourdes Higuchi, M.; De Brito, T.; Reis, M.M.; Barbosa, A.; Bellotti, G.; Pereira-Barreto, A.C.; Pileggi, F. Correlation between Trypanosoma cruzi parasitism and myocardial inflammatory infiltrate in human chronic chagasic myocarditis: Light microscopy and immunohistochemical findings. Cardiovasc. Pathol. 1993, 2, 101–106. [Google Scholar] [CrossRef]
- Rossi, M.A. The pattern of myocardial fibrosis in chronic Chagas’ heart disease. Int. J. Cardiol. 1991, 30, 335–340. [Google Scholar] [CrossRef]
- Rassi, A., Jr.; Rassi, A.; Marin-Neto, J.A. Chagas heart disease: Pathophysiologic mechanisms, prognostic factors and risk stratification. Mem. Inst. Oswaldo Cruz 2009, 104, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Rassi, A.; Rassi, A.; Rassi, S.G. Predictors of Mortality in Chronic Chagas Disease: A Systematic Review of Observational Studies. Circulation 2007, 115, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Coura, J.R.; Borges-Pereira, J. Chronic phase of Chagas disease: Why should it be treated? A comprehensive review. Mem. Inst. Oswaldo Cruz 2011, 106, 641–645. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Kroh, E.M.; Parkin, R.K.; Mitchell, P.S.; Tewari, M. Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 2010, 50, 298–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, N.A.; Dehlendorff, C.; Jensen, B.V.; Bjerregaard, J.K.; Nielsen, K.R.; Bojesen, S.E.; Andersen, K.K. MicroRNA biomarkers in whole blood for detection of pancreatic cancer. JAMA 2014, 311, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Scrutinio, D.; Conserva, F.; Passantino, A.; Iacoviello, M.; Lagioia, R.; Gesualdo, L. Circulating microRNA-150-5p as a novel biomarker for advanced heart failure: A genome-wide prospective study. J. Heart Lung Transplant. 2017, 36, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Nana-Sinkam, S.P. Detection of microRNA Expression in Human Peripheral Blood Microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Li, Q. Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Widera, C.; Gupta, S.K.; Lorenzen, J.M.; Bang, C.; Bauersachs, J.; Bethmann, K.; Thum, T. Diagnostic and prognostic impact of six circulating microRNAs in acute coronary syndrome. J. Mol. Cell. Cardiol. 2011, 51, 872–875. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, J.; Evan, G.; Xiao, C.; Cheng, Y.; Xiao, J. Circulating microRNAs: Novel biomarkers for cardiovascular diseases. J. Mol. Med. 2012, 90, 865–875. [Google Scholar] [CrossRef]
- Miao, Y.; Chen, H.; Li, M. MiR-19a overexpression contributes to heart failure through targeting ADRB1. Int. J. Clin. Exp. Med. 2015, 8, 642–649. [Google Scholar]
- Villar, A.V.; García, R.; Merino, D.; Llano, M.; Cobo, M.; Montalvo, C.; Nistal, J.F. Myocardial and circulating levels of microRNA-21 reflect left ventricular fibrosis in aortic stenosis patients. Int. J. Cardiol. 2013, 167, 2875–2881. [Google Scholar] [CrossRef] [Green Version]
- Roncarati, R.; Anselmi, C.V.; Losi, M.A.; Papa, L.; Cavarretta, E.; Martins, P.D.C.; Latronico, M.V. Circulating miR-29a, among other up-regulated microRNAs, is the only biomarker for both hypertrophy and fibrosis in patients with hypertrophic cardiomyopathy. J. Am. Coll. Cardiol. 2014, 63, 920–927. [Google Scholar] [CrossRef]
- Shi, J.Y.; Chen, C.; Xu, X.; Lu, Q. miR-29a promotes pathological cardiac hypertrophy by targeting the PTEN/AKT/mTOR signalling pathway and suppressing autophagy. Acta Physiol. (Oxf). 2019, e13323. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, L.; Wang, H.; Chen, J.; Li, W.; Liu, W.; Fu, X. Long noncoding RNA CNALPTC1 promotes cell proliferation and migration of papillary thyroid cancer via sponging miR-30 family. Am. J. Cancer Res. 2018, 8, 192. [Google Scholar]
- Duygu, B.; Poels, E.M.; Juni, R.; Bitsch, N.; Ottaviani, L.; Olieslagers, S.; Da Costa Martins, P.A. miR-199b-5p is a regulator of left ventricular remodeling following myocardial infarction. Non-Coding RNA Res. 2017, 2, 18–26. [Google Scholar] [CrossRef]
- Ferreira, L.R.P.; Frade, A.F.; Santos, R.H.B.; Teixeira, P.C.; Baron, M.A.; Navarro, I.C.; Chevillard, C. MicroRNAs miR-1, miR-133a, miR-133b, miR-208a and miR-208b are dysregulated in Chronic Chagas disease Cardiomyopathy. Int. J. Cardiol. 2014, 175, 409–417. [Google Scholar] [CrossRef]
- Linhares-Lacerda, L.; Granato, A.; Gomes-Neto, J.F.; Conde, L.; Freire-de-Lima, L.; De Freitas, E.O.; Savino, W. Circulating Plasma MicroRNA-208a as Potential Biomarker of Chronic Indeterminate Phase of Chagas Disease. Front. Microbiol. 2018, 9, 269. [Google Scholar] [CrossRef]
- Wang, K.; Yuan, Y.; Cho, J.H.; McClarty, S.; Baxter, D.; Galas, D.J. Comparing the MicroRNA spectrum between serum and plasma. PLoS ONE 2012, 7, e41561. [Google Scholar] [CrossRef]
- Araújo-Jorge, T.C.; Waghabi, M.C.; Hasslocher-Moreno, A.M.; Xavier, S.S.; Higuchi Mde, L.; Keramidas, M.; Bailly, S.; Feige, J.J. Implication of transforming growth factor-beta1 in Chagas disease myocardiopathy. J. Infect. Dis. 2002, 186, 1823–1828. [Google Scholar] [CrossRef]
- Cunha-Neto, E.; Chevillard, C. Chagas disease cardiomyopathy: Immunopathology and genetics. Mediat. Inflamm. 2014, 2014, 683230. [Google Scholar] [CrossRef]
- Ferreira, L.R.P.; Ferreira, F.M.; Nakaya, H.I.; Deng, X.; Cândido, D.D.S.; De Oliveira, L.C.; Kalil, J. Blood Gene Signatures of Chagas disease Cardiomyopathy with or without ventricular dysfunction. J. Infect. Dis. 2016, 215, 387–395. [Google Scholar] [CrossRef]
- Song, D.W.; Ryu, J.Y.; Kim, J.O.; Kwon, E.J.; Kim, D.H. The miR-19a/b family positively regulates cardiomyocyte hypertrophy by targeting atrogin-1 and MuRF-1. Biochem. J. 2014, 457, 151–162. [Google Scholar] [CrossRef]
- Lai, Y.; He, S.; Ma, L.; Lin, H.; Ren, B.; Ma, J.; Zhuang, S. HOTAIR functions as a competing endogenous RNA to regulate PTEN expression by inhibiting miR-19 in cardiac hypertrophy. Mol. Cell. Biochem. 2017, 432, 179–187. [Google Scholar] [CrossRef]
- Zhong, J.; He, Y.; Chen, W.; Shui, X.; Chen, C.; Lei, W. Circulating microRNA-19a as a potential novel biomarker for diagnosis of acute myocardial infarction. Int. J. Mol. Sci. 2014, 15, 20355–20364. [Google Scholar] [CrossRef]
- Machado, F.S.; Jelicks, L.A.; Kirchhoff, L.V.; Shirani, J.; Nagajyothi, F.; Mukherjee, S.; Guan, F. Chagas heart disease: Report on recent developments. Cardiol. Rev. 2012, 20, 53–65. [Google Scholar] [CrossRef]
- Sassi, Y.; Avramopoulos, P.; Ramanujam, D.; Grüter, L.; Werfel, S.; Giosele, S.; Hübner, N. Cardiac myocyte miR-29 promotes pathological remodeling of the heart by activating Wnt signaling. Nat. Commun. 2017, 8, 1614. [Google Scholar] [CrossRef]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Castoldi, M. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Adam, O.; Löhfelm, B.; Thum, T.; Gupta, S.K.; Puhl, S.L.; Schäfers, H.J.; Laufs, U. Role of miR-21 in the pathogenesis of atrial fibrosis. Basic Res. Cardiol. 2012, 107, 278. [Google Scholar] [CrossRef]
- Dong, S.; Ma, W.; Hao, B.; Hu, F.; Yan, L.; Yan, X.; Wang, Z. microRNA-21 promotes cardiac fibrosis and development of heart failure with preserved left ventricular ejection fraction by up-regulating Bcl-2. Int. J. Clin. Exp. Pathol. 2014, 7, 565–574. [Google Scholar]
- Tostes, S.; Bertulucci Rocha-Rodrigues, D.; De Araujo Pereira, G.; Rodrigues, V. Myocardiocyte apoptosis in heart failure in chronic Chagas’ disease. Int. J. Cardiol. 2005, 99, 233–237. [Google Scholar] [CrossRef]
- Liu, G.; Friggeri, A.; Yang, Y.; Milosevic, J.; Ding, Q.; Thannickal, V.J.; Abraham, E. miR-21 mediates fibrogenic activation of pulmonary fibroblasts and lung fibrosis. J. Exp. Med. 2010, 207, 1589–1597. [Google Scholar] [CrossRef]
- Zhong, X.; Chung, A.C.; Chen, H.Y.; Meng, X.M.; Lan, H.Y. Smad3-mediated upregulation of miR-21 promotes renal fibrosis. J. Am. Soc. Nephrol. 2011, 22, 1668–1681. [Google Scholar] [CrossRef]
- Reddy, S.; Hu, D.Q.; Zhao, M.; Blay, E., Jr.; Sandeep, N.; Ong, S.G.; Bernstein, D. miR-21 is associated with fibrosis and right ventricular failure. JCI Insight 2017, 2, e91625. [Google Scholar] [CrossRef]
- Duygu, B.; Da Costa Martins, P.A. miR-21: A star player in cardiac hypertrophy. Cardiovasc. Res. 2015, 105, 235–237. [Google Scholar] [CrossRef]
- Navarro, I.C.; Ferreira, F.M.; Nakaya, H.I.; Baron, M.A.; Vilar-Pereira, G.; Pereira, I.R.; Lannes-Vieira, J. MicroRNA Transcriptome Profiling in Heart of Trypanosoma cruzi-Infected Mice: Parasitological and Cardiological Outcomes. PLoS Negl. Trop. Dis. 2015, 9, e0003828. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.H.; Yang, R.; Yang, B.J.; Gao, Z.Y. Association between circulating microRNA-208a and severity of coronary heart disease. Scand. J. Clin. Lab. Investig. 2017, 77, 379–384. [Google Scholar] [CrossRef]
- Cao, W.; Shi, P.; Ge, J.J. miR-21 enhances cardiac fibrotic remodeling and fibroblast proliferation via CADM1/STAT3 pathway. BMC Cardiovasc. Disord. 2017, 17, 88. [Google Scholar] [CrossRef]
- Aggarwal, P.; Turner, A.; Matter, A.; Kattman, S.J.; Stoddard, A.; Lorier, R.; Broeckel, U. RNA Expression Profiling of Human iPSC-Derived Cardiomyocytes in a Cardiac Hypertrophy Model. PLoS ONE 2014, 9, e1080cc51. [Google Scholar] [CrossRef]
- Wijnen, W.J.; Van der Made, I.; Van den Oever, S.; Hiller, M.; De Boer, B.A.; Picavet, D.I.; Van Veen, H. Cardiomyocyte-specific miRNA-30c over-expression causes dilated cardiomyopathy. PLoS ONE 2014, 9, e96290. [Google Scholar] [CrossRef]
- Thum, T. Noncoding RNAs and myocardial fibrosis. Nat. Rev. Cardiol. 2014, 11, 655. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, H.Y.; Li, Y.; Guo, S.H.; Zhang, L.; Cai, J.H. Peripheral blood miRNAs as a biomarker for chronic cardiovascular diseases. Sci. Rep. 2015, 4, 5026. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Diehl, P.; Fricke, A.; Sander, L.; Stamm, J.; Bassler, N.; Htun, N.; Ziemann, M.; Helbing, T.; El-Osta, A.; Jowett, J.B.; et al. Microparticles: Major transport vehicles for distinct microRNAs in circulation. Cardiovasc. Res. 2012, 93, 633–644. [Google Scholar] [CrossRef]
- Long, G.; Wang, F.; Duan, Q.; Yang, S.; Chen, F.; Gong, W.; Yang, X.; Wang, Y.; Chen, C.; Wang, D.W. Circulating miR-30a, miR-195 and let-7b associated with acute myocardial infarction. PLoS ONE 2012, 7, e50926. [Google Scholar]
- Long, G.; Wang, F.; Li, H.; Yin, Z.; Sandip, C.; Lou, Y.; Wang, Y.; Chen, C.; Wang, D.W. Circulating miR-30a, miR-126 and let-7b as biomarker for ischemic stroke in humans. BMC Neurol. 2013, 13, 178. [Google Scholar] [CrossRef]
- Zhao, D.S.; Chen, Y.; Jiang, H.; Lu, J.P.; Zhang, G.; Geng, J.; Zhang, Q.; Shen, J.H.; Zhou, X.; Zhu, W.; et al. Serum miR-210 and miR-30a expressions tend to revert to fetal levels in Chinese adult patients with chronic heart failure. Cardiovasc. Pathol. 2013, 22, 444–450. [Google Scholar] [CrossRef]
- De Rosa, S.; Fichtlscherer, S.; Lehmann, R.; Assmus, B.; Dimmeler, S.; Zeiher, A.M. Transcoronary concentration gradients of circulating microRNAs. Circulation 2011, 124, 1936–1944. [Google Scholar] [CrossRef]
- Liebetrau, C.; Möllmann, H.; Dörr, O.; Szardien, S.; Troidl, C.; Willmer, M.; Voss, S.; Gaede, L.; Rixe, J.; Rolf, A.; et al. Release kinetics of circulating muscle-enriched microRNAs in patients undergoing transcoronary ablation of septal hypertrophy. J. Am. College Cardiol. 2013, 62, 992–998. [Google Scholar] [CrossRef]
- Li, C.; Fang, Z.; Jiang, T.; Zhang, Q.; Liu, C.; Zhang, C.; Xiang, Y. Serum microRNAs profile from genome-wide serves as a fingerprint for diagnosis of acute myocardial infarction and angina pectoris. BMC Med. Genomics 2013, 6, 16. [Google Scholar]
- Barry, S.E.; Chan, B.; Ellis, M.; Yang, Y.; Plit, M.L.; Guan, G.; Saunders, B.M. Identification of miR-93 as a suitable miR for normalizing miRNA in plasma of tuberculosis patients. J. Cell. Mol. Med. 2015, 19, 1606–1613. [Google Scholar] [CrossRef]
- Lange, T.; Stracke, S.; Rettig, R.; Lendeckel, U.; Kuhn, J.; Schlüter, R.; Endlich, N. Identification of miR-16 as an endogenous reference gene for the normalization of urinary exosomal miRNA expression data from CKD patients. PLoS ONE 2017, 12, e0183435. [Google Scholar] [CrossRef]
- Smith, W.H.; Nair, R.U.; Adamson, D.; Kearney, M.T.; Ball, S.G.; Balmforth, A.J. Somatostatin receptor subtype expression in the human heart: Differential expression by myocytes and fibroblasts. J. Endocrinol. 2005, 187, 379–386. [Google Scholar] [CrossRef]
- Okita, K.; Matsumura, Y.; Sato, Y.; Okada, A.; Morizane, A.; Okamoto, S.; Shibata, T. A more efficient method to generate integration-free human iPS cells. Nat. Methods 2011, 8, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Indeterminate Form (n = 10) | CCC (n = 28) |
|---|---|---|
| Age (years) | 59 ± 8 | 60 ± 7 |
| Male gender | 7 (70%) | 18 (65%) |
| % fibrosis (CMR) | 3.1 ± 5 | 19 ± 15 |
| EF (CMR) | 77.5 ± 13 | 32 ± 10 |
| Strain (LV) | 22.5 ± 6 | 10 ± 4 |
| Ventricular arrhythmia | 8 (80%) | 28 (100%) |
| SV arrythmia | 8 (80%) | 23 (82%) |
| HR variability | 123 ± 22 | 120 ± 39 |
| miRNAs | Cardiac Dysfunction | Age | NYHA (II-IV) | %Fibrosis | EF (CMR) | Strain (LV) | Ventricular Arrhythmia | SV Arrhythmia | HR Variability |
|---|---|---|---|---|---|---|---|---|---|
| miR-19a-3p | r = 0.47 | r = 0.20 | r = 0.47 | r = 0.41 | r = −0.40 | r = −0.36 | r = 0.15 | r = 0.09 | r = −0.11 |
| p = 0.003 ** | p = 0.22 | p = 0.003 ** | p = 0.01 * | p = 0.01 * | p = 0.02 * | p = 0.37 | p = 0.59 | p = 0.54 | |
| miR-21-5p | r = 0.42 | r = 0.06 | r = 0.42 | r = 0.38 | r = −0.40 | r = −0.34 | r = 0.16 | r = 0.02 | r = −0.21 |
| p = 0.008 ** | p = 0.70 | p = 0.008 ** | p = 0.03 * | p = 0.01 * | p = 0.04 * | p = 0.32 | p = 0.89 | p = 0.24 | |
| miR-29b-3p | r = 0.47 | r = 0.04 | r = 0.47 | r = 0.36 | r = −0.34 | r = −0.32 | r = 0.24 | r = 0.27 | r = −0.34 |
| p = 0.02 * | p = 0.79 | p = 0.02 * | p = 0.03 * | p = 0.03 * | p = 0.05 | p = 0.13 | p = 0.09 | p = 0.05 | |
| miR-199b-3p | r = 0.24 | r = −0.27 | r = 0.24 | r = 0.01 | r = −0.28 | r = −0.18 | r = 0.03 | r = −0.07 | r = −0.11 |
| p = 0.16 | p = 0.12 | p = 0.16 | p = 0.96 | p = 0.10 | p = 0.31 | p = 0.85 | p = 0.66 | p = 0.54 |
| microRNA | Disease | Sample | Regulation | References |
|---|---|---|---|---|
| miR-19a | Heart failure | plasma | upregulated | [18] |
| miR-21 | Aortic stenosis | plasma | upregulated | [19] |
| miR-21 | Acute coronary syndrome | plasma | upregulated | [51] |
| miR-21 | Chronic cardiovascular disease (atherosclerosis) | peripheral blood | upregulated | [49] |
| miR-21 | Congestive heart failure | serum | upregulated | [17] |
| miR-29 | Hypertrophy cardiomyopathy | plasma | upregulated | [20] |
| miR-30a | Acute myocardial infarction | plasma | upregulated | [52] |
| miR-30a | Ischemic stroke | plasma | upregulated | [53] |
| miR-30a | Chronic heart failure | serum | upregulated | [54] |
| miR-30a | Hypertrophy cardiomyopathy | plasma | upregulated | [20] |
| miR-199 | Hypertrophy cardiomyopathy | plasma | upregulated | [20] |
| miR-208a | Coronary artery disease | plasma | upregulated | [55] |
| miR-208a | Acute myocardial infarction (TASH: transcoronary ablation of septal hypertrophy) | serum | upregulated | [56] |
| miR-208 | Acute myocardial infarction | plasma | upregulated | [57] |
| miR-208a | Chronic Chagas disease | plasma | upregulated | [25] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nonaka, C.K.V.; Macêdo, C.T.; Cavalcante, B.R.R.; Alcântara, A.C.d.; Silva, D.N.; Bezerra, M.d.R.; Caria, A.C.I.; Tavora, F.R.F.; Neto, J.D.d.S.; Noya-Rabelo, M.M.; et al. Circulating miRNAs as Potential Biomarkers Associated with Cardiac Remodeling and Fibrosis in Chagas Disease Cardiomyopathy. Int. J. Mol. Sci. 2019, 20, 4064. https://doi.org/10.3390/ijms20164064
Nonaka CKV, Macêdo CT, Cavalcante BRR, Alcântara ACd, Silva DN, Bezerra MdR, Caria ACI, Tavora FRF, Neto JDdS, Noya-Rabelo MM, et al. Circulating miRNAs as Potential Biomarkers Associated with Cardiac Remodeling and Fibrosis in Chagas Disease Cardiomyopathy. International Journal of Molecular Sciences. 2019; 20(16):4064. https://doi.org/10.3390/ijms20164064
Chicago/Turabian StyleNonaka, Carolina Kymie Vasques, Carolina Thé Macêdo, Bruno Raphael Ribeiro Cavalcante, Adriano Costa de Alcântara, Daniela Nascimento Silva, Milena da Rocha Bezerra, Alex Cleber Improta Caria, Fábio Rocha Fernandes Tavora, João David de Souza Neto, Márcia Maria Noya-Rabelo, and et al. 2019. "Circulating miRNAs as Potential Biomarkers Associated with Cardiac Remodeling and Fibrosis in Chagas Disease Cardiomyopathy" International Journal of Molecular Sciences 20, no. 16: 4064. https://doi.org/10.3390/ijms20164064
APA StyleNonaka, C. K. V., Macêdo, C. T., Cavalcante, B. R. R., Alcântara, A. C. d., Silva, D. N., Bezerra, M. d. R., Caria, A. C. I., Tavora, F. R. F., Neto, J. D. d. S., Noya-Rabelo, M. M., Rogatto, S. R., Ribeiro dos Santos, R., Souza, B. S. d. F., & Soares, M. B. P. (2019). Circulating miRNAs as Potential Biomarkers Associated with Cardiac Remodeling and Fibrosis in Chagas Disease Cardiomyopathy. International Journal of Molecular Sciences, 20(16), 4064. https://doi.org/10.3390/ijms20164064