The Highly Conserved Barley Powdery Mildew Effector BEC1019 Confers Susceptibility to Biotrophic and Necrotrophic Pathogens in Wheat


Abstract
:1. Introduction
2. Results
2.1. BEC1019 is Broadly Conserved in Plant Fungal Pathogens
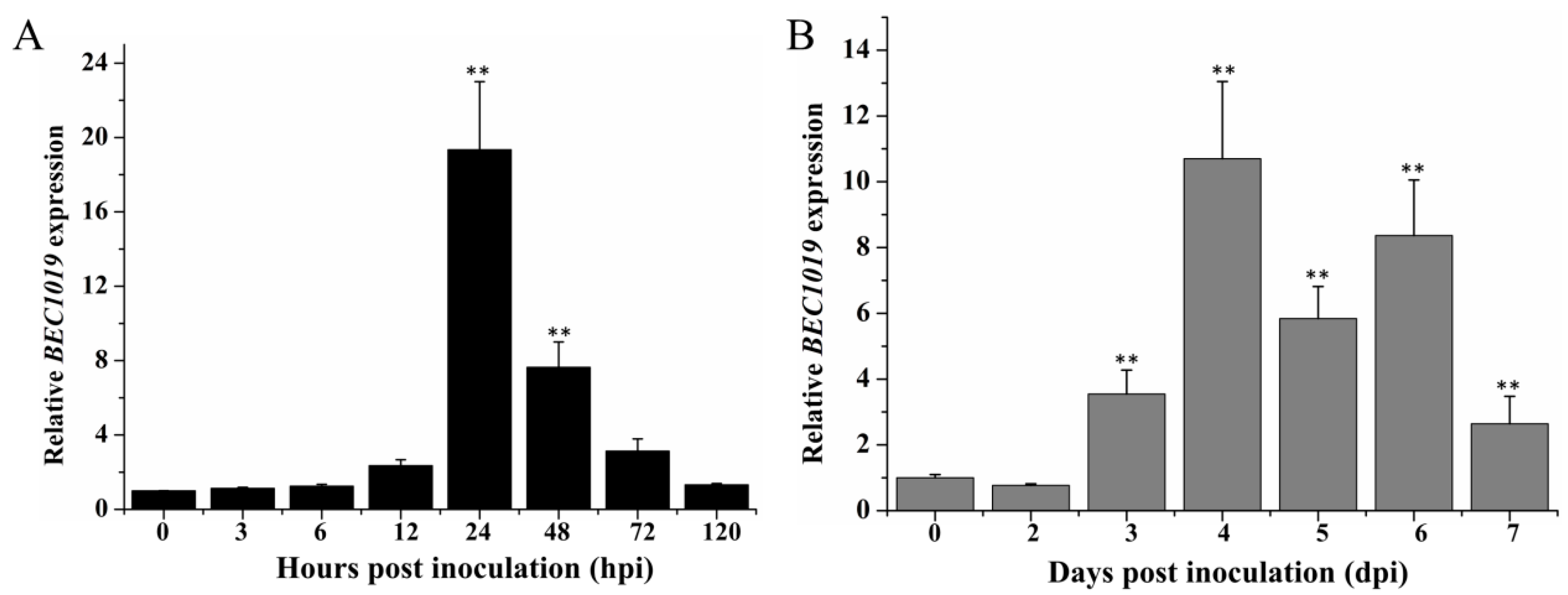
2.2. BEC1019 Is Highly Expressed during Haustorium Formation by Bgt and Concomitant with the Development of Necrotic Symptoms in Ggt
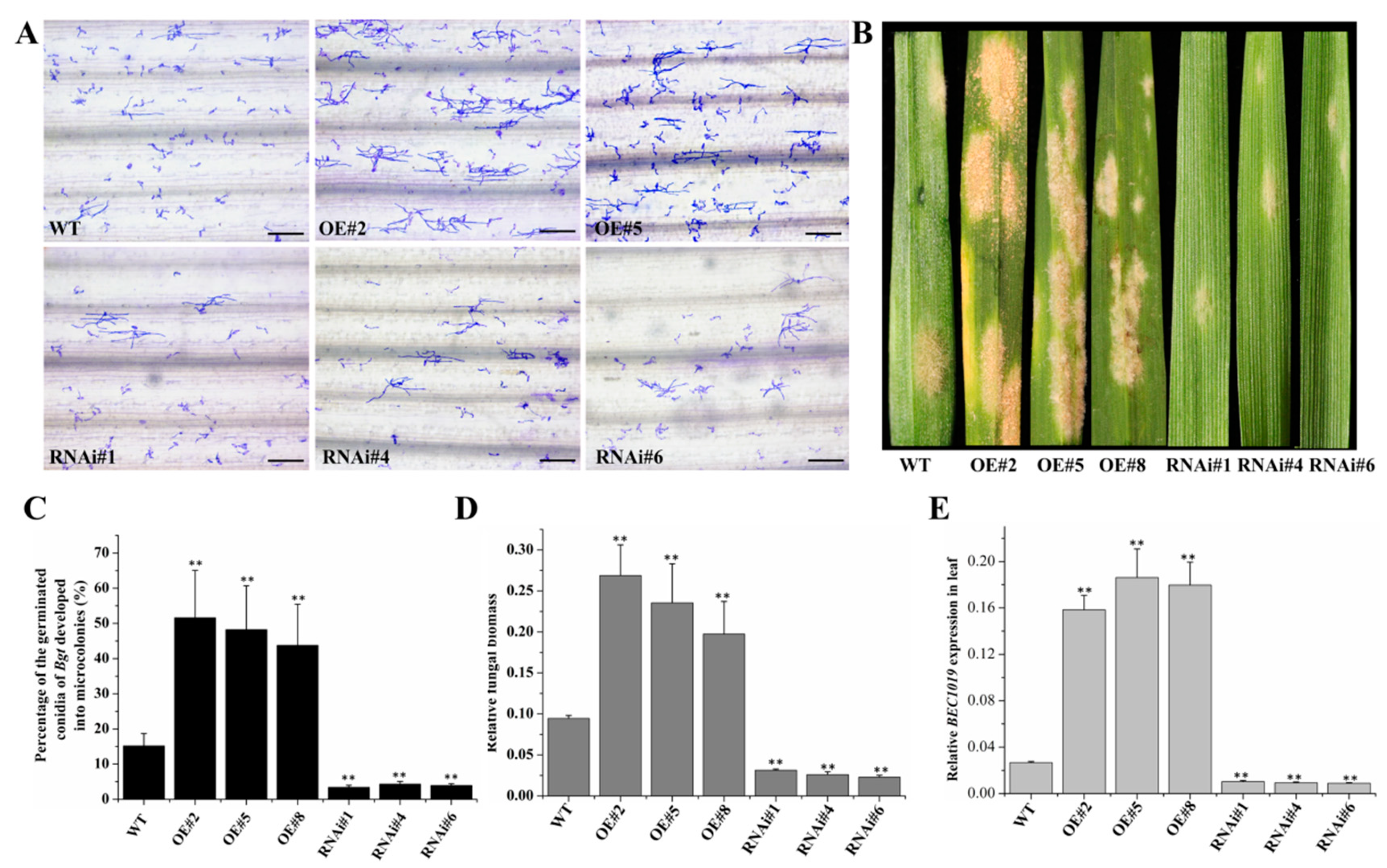
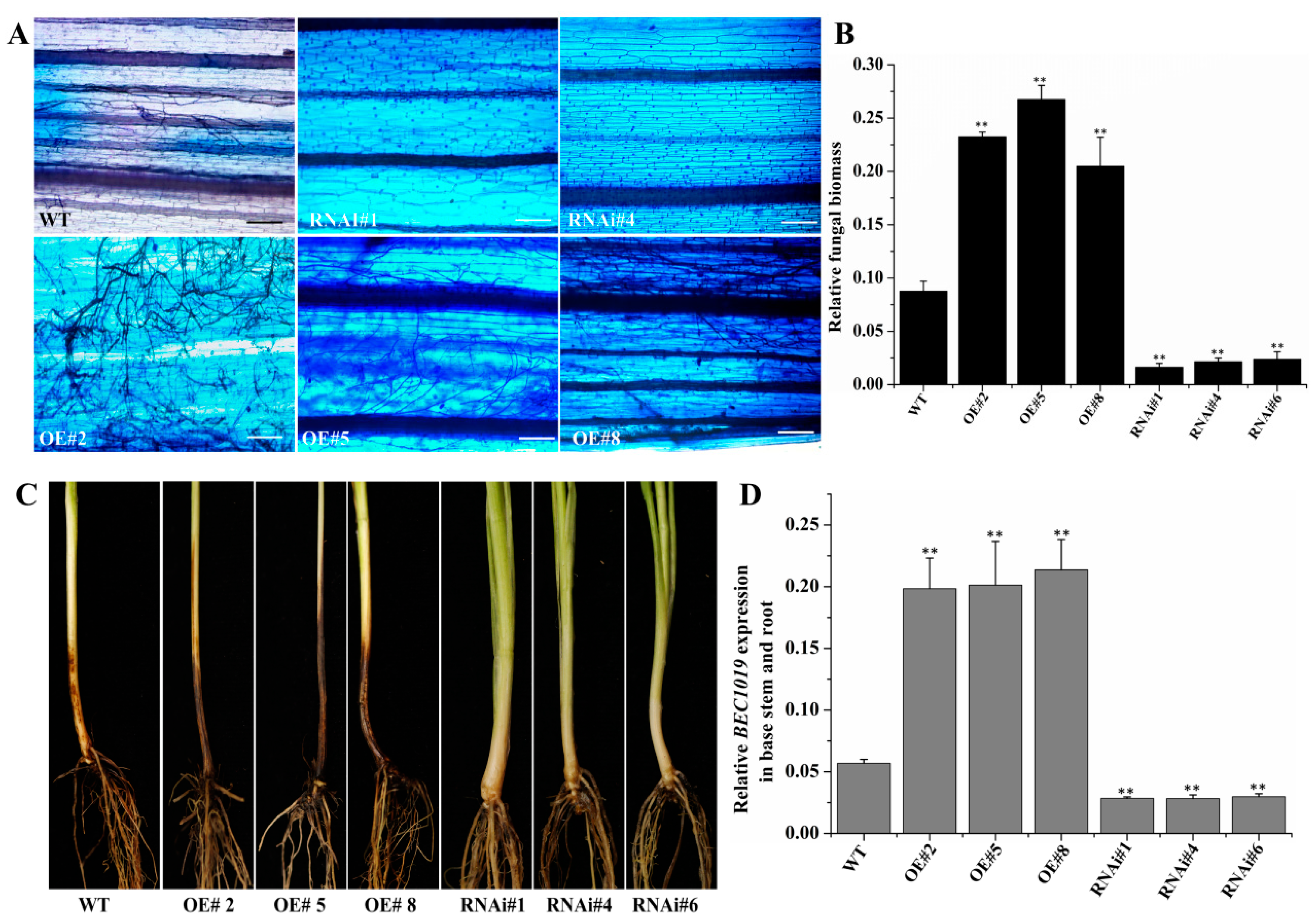
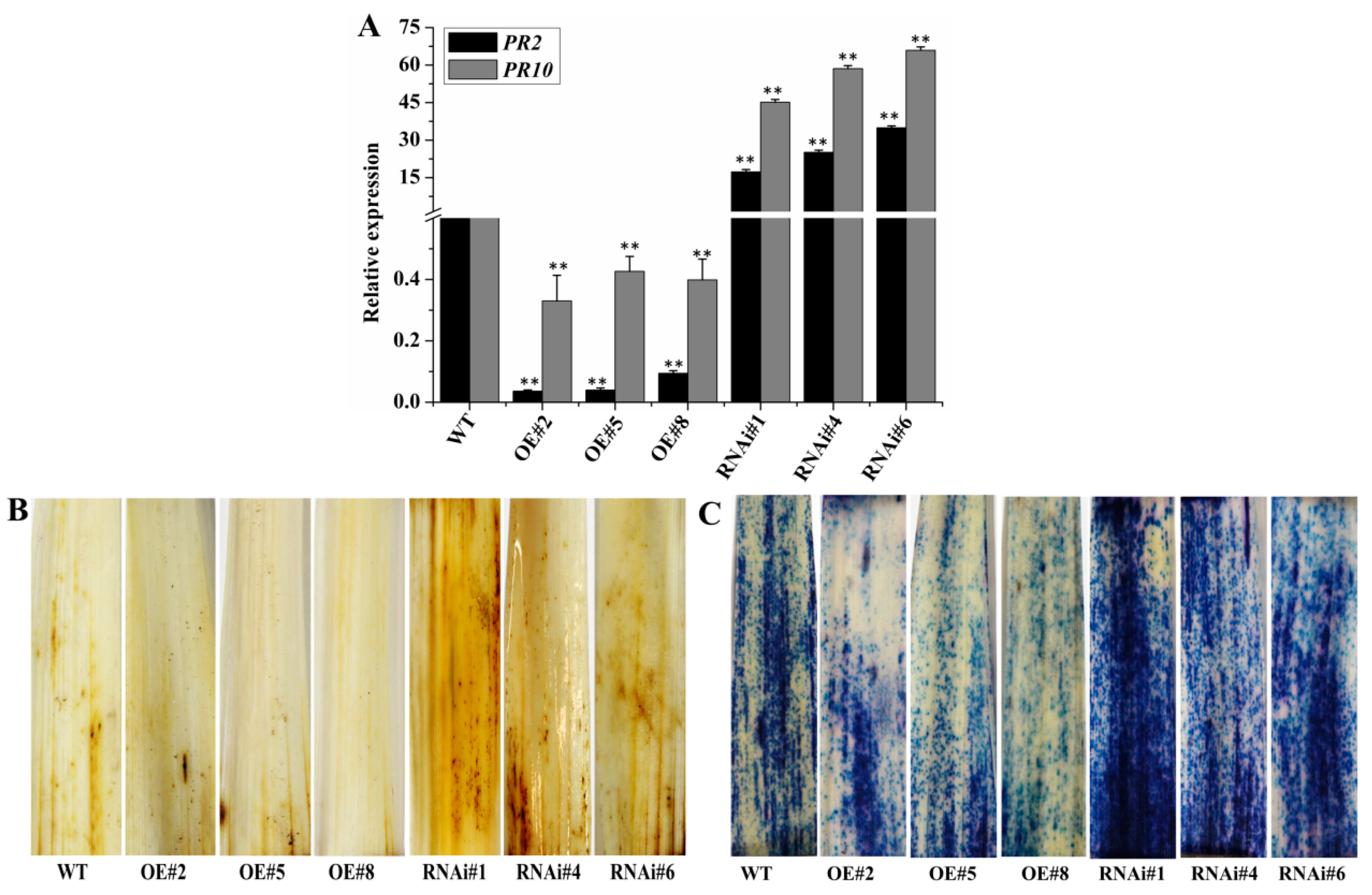
2.3. Overexpression of BEC1019 Increases Wheat Susceptibility to Bgt and Ggt, whereas Silencing BEC1019 Enhances Resistance to both Biotrophic and Necrotrophic Pathogens
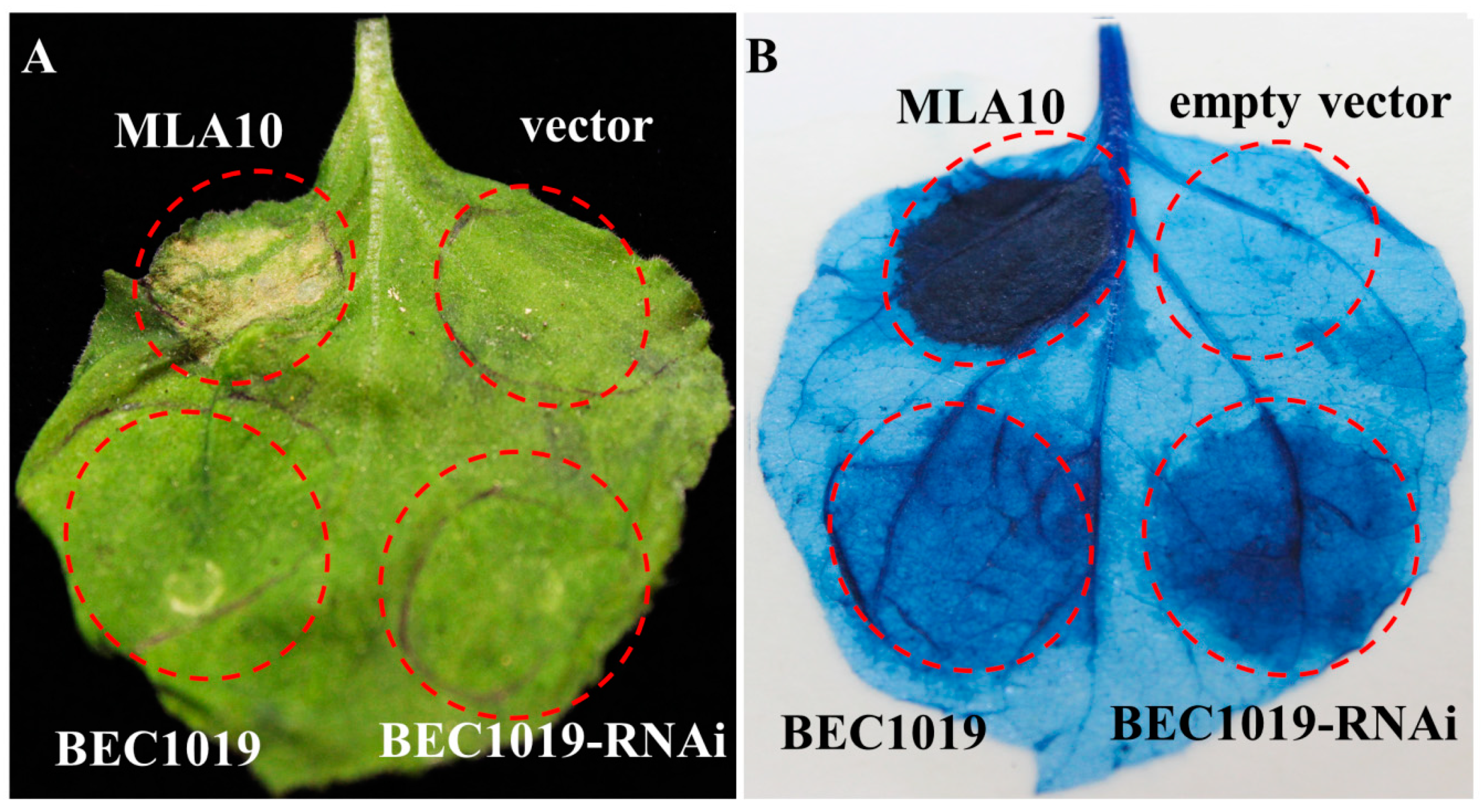
2.4. Transient Expression of BEC1019 Protein Triggers Cell Death in Nicotiana benthamiana
3. Discussion
4. Materials and Methods
4.1. Plants and Fungal Materials
4.2. Cloning of BEC1019 Homologous Genes
4.3. Construction of BEC1019 Overexpression and RNAi Vectors
4.4. DNA and RNA Extraction and Quantitative Real-Time PCR
4.5. Detection of H2O2 and Cell Death
4.6. Responses of Transgenic Wheat Plants to Bgt and Ggt
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Godfray, H.C.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Liu, Y.; Naqvi, N.I. Fungal effectors at the crossroads of phytohormone signaling. Curr. Opin. Microbiol. 2018, 46, 1–6. [Google Scholar] [CrossRef]
- Macho, A.P.; Cyril, Z. Plant PRRs and the activation of innate immune signaling. Mol. Cell 2014, 54, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Thomas, B.; Yang, H.S. Innate immunity in plants: An arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–744. [Google Scholar]
- Zhang, J.; Zhou, J.M. Plant immunity triggered by microbial molecular signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef]
- Jacob, F.; Vernaldi, S.; Maekawa, T. Evolution and conservation of plant NLR functions. Front. Immuno. 2013, 4, 297. [Google Scholar] [CrossRef]
- Stella, C.; Maud, B.; Philippe, M.; Thomas, K.; Dodds, P.N. A novel conserved mechanism for plant NLR protein pairs: The “integrated decoy” hypothesis. Front. Plant Sci. 2014, 5, 606. [Google Scholar]
- Stergiopoulos, I.; Wit, P.J.G.M.D. Fungal effector proteins. Annu. Rev. Phytopathol. 2009, 47, 233. [Google Scholar] [CrossRef] [PubMed]
- Bithell, S.L.; McKay, A.C.; Butler, R.C.; Cromey, M.G. Consecutive wheat sequences: Effects of contrasting growing seasons on concentrations of Gaeumannomyces graminis var. tritici DNA in soil and take-all disease across different cropping sequences. J. Agric. Sci. 2016, 154, 472–486. [Google Scholar]
- Gutteridge, R.J.; Bateman, G.L.; Todd, A.D. Variation in the effects of take-all disease on grain yield and quality of winter cereals in field experiments. Pest Manag. Sci. 2003, 59, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Guilleroux, M.; Osbourn, A. Gene expression during infection of wheat roots by the ‘take-all’ fungus Gaeumannomyces graminis. Mol. Plant Pathol. 2004, 5, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Huang, L.; Kang, Z.; Buchenauer, H. Evaluation of endophytic bacterial strains as antagonists of take-all in wheat caused by Gaeumannomyces graminis var. tritici in greenhouse and field. J. Pest Sci. 2011, 84, 257–264. [Google Scholar]
- Barret, M.; Frey-Klett, P.; Guillerm-Erckelboudt, A.Y.; Boutin, M.; Guernec, G.; Sarniguet, A. Effect of wheat roots infected with the pathogenic fungus Gaeumannomyces graminis var. tritici on gene expression of the biocontrol bacterium Pseudomonas fluorescens Pf29Arp. Mol. Plant-Microbe Interact. 2009, 22, 1611–1623. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Zhang, W.; Cai, X.; Zhu, T.; Xue, Y.; Liu, C. Bacillus velezensis CC09: A potential ‘vaccine’ for controlling wheat diseases. Mol. Plant-Microbe Interact. 2018, 31, 623–632. [Google Scholar] [CrossRef]
- Wang, M.; Xing, Y.; Wang, J.; Xu, Y.; Wang, G. The role of the chi1 gene from the endophytic bacteria Serratia proteamaculans 336x in the biological control of wheat take-all. Can. J. Microbiol. 2014, 60, 533–540. [Google Scholar] [CrossRef]
- Zhang, D.D.; Guo, X.J.; Wang, Y.J.; Gao, T.G.; Zhu, B.C. Novel screening strategy reveals a potent Bacillus antagonist capable of mitigating wheat take-all disease caused by Gaeumannomyces graminis var. tritici. Lett. Appl. Microbiol. 2017, 65, 512–519. [Google Scholar] [CrossRef]
- Panstruga, R. Establishing compatibility between plants and obligate biotrophic pathogens. Curr. Opin. Plant Biol. 2003, 6, 320–326. [Google Scholar] [CrossRef]
- Kamoun, S. A catalogue of the effector secretome of plant pathogenic oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; Thorsten, N.; Joosten, M.H.A.J. Of PAMPs and effectors: The blurred PTI-ETI dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Qian-Hua, S.; Yusuke, S.; Stefan, M.; Christoph, B.; Stéphane, B.; Beat, K.; Hikaru, S.; Bekir, U.; Somssich, I.E.; Paul, S.L. Nuclear activity of MLA immune receptors links isolate-specific and basal disease-resistance responses. Science 2007, 315, 1098–1103. [Google Scholar]
- Ridout, C.; Skamnioti, P.; Porritt, O.; Sacristan, S.; Jones, J.D.G.; Brown, J. Multiple avirulence paralogues in cereal powdery mildew fungi may contribute to parasite fitness and defeat of plant resistance. Plant Cell 2006, 18, 2402–2414. [Google Scholar] [CrossRef]
- Lu, X.; Kracher, B.; Saur, I.M.L.; Bauer, S.; Ellwood, S.R.; Wise, R.; Yaeno, T.; Maekawa, T.; Schulze-Lefert, P. Allelic barley MLA immune receptors recognize sequence-unrelated avirulence effectors of the powdery mildew pathogen. Proc. Natl. Acad. Sci. USA 2016, 113, E6486–E6495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spanu, P.D.; Abbott, J.C.; Amselem, J.; Burgis, T.A.; Soanes, D.M.; Stuber, K.; Ver Loren van Themaat, E.; Brown, J.K.; Butcher, S.A.; Gurr, S.J. Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism. Science 2010, 330, 1543–1546. [Google Scholar] [CrossRef]
- Wicker, T.; Oberhaensli, S.; Parlange, F.; Buchmann, J.P.; Shatalina, M.; Roffler, S.; Ben-David, R.; Dolezel, J.; Simkova, H.; Schulze-Lefert, P. The wheat powdery mildew genome shows the unique evolution of an obligate biotroph. Nat. Genet. 2013, 45, 1092–1096. [Google Scholar] [CrossRef] [Green Version]
- Ali Abdurehim, A.; Carsten, P.; Torsten, S.L.; Mark, K.; JoRgensen, H.J.L.; Hans, T.C. The barley powdery mildew candidate secreted effector protein CSEP0105 inhibits the chaperone activity of a small heat shock protein. Plant Physiol. 2015, 168, 321–333. [Google Scholar]
- Zhang, W.J.; Pedersen, C.; Kwaaitaal, M.; Gregersen, P.L.; Mørch, S.M.; Hanisch, S.; Kristensen, A.; Fuglsang, A.T.; Collinge, D.B.; Thordal-Christensen, H. Interaction of barley powdery mildew effector candidate CSEP0055 with the defence protein PR17c. Mol. Plant Pathol. 2012, 13, 1110–1119. [Google Scholar] [CrossRef]
- Pliego, C.; Nowara, D.; Bonciani, G.; Gheorghe, D.M.; Xu, R.; Surana, P.; Whigham, E.; Nettleton, D.; Bogdanove, A.J.; Wise, R.P. Host-induced gene silencing in barley powdery mildew reveals a class of ribonuclease-like effectors. Mol. Plant Microbe Interact. 2013, 26, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef] [PubMed]
- Whigham, E.; Qi, S.; Mistry, D.; Surana, P.; Xu, R.; Fuerst, G.; Pliego, C.; Bindschedler, L.V.; Spanu, P.D.; Dickerson, J.A. Broadly conserved fungal effector BEC1019 suppresses host cell Death and enhances pathogen virulence in powdery mildew of barley (Hordeum vulgare L.). Mol. Plant Microbe Interact. 2015, 28, 968–983. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Bogdanove, A.J.; Roger, I.W.; Roger, W.P. A confounding effect of bacterial titer in a type III delivery-based assay of eukaryotic effector function. Mol. Plant-Microbe Interact. 2018, 31, 1115–1116. [Google Scholar] [CrossRef] [PubMed]
- Both, M.; Csukai, M.; Stumpf, M.P.; Spanu, P.D. Gene expression profiles of Blumeria graminis indicate dynamic changes to primary metabolism during development of an obligate biotrophic pathogen. Plant Cell 2005, 17, 2107–2122. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Liu, J.; Chang, C.; Zhang, L.; Maekawa, T.; Wang, Q.; Xiao, W.; Liu, Y.; Chai, J.; Takken, F.L. Structure-function analysis of barley NLR immune receptor MLA10 reveals its cell compartment specific activity in cell death and disease resistance. PLoS Pathog. 2012, 8, e1002752. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, G.B.; Pedersen, C.; Thordal-Christensen, H. Identification of eight effector candidate genes involved in early aggressiveness of the barley powdery mildew fungus. Plant Pathol. 2016, 65, 953–958. [Google Scholar] [CrossRef]
- Qi, T.; Zhu, X.; Tan, C.; Liu, P.; Guo, J.; Kang, Z. Host-induced gene silencing of an important pathogenicity factor PsCPK1 in Puccinia striiformis f. sp. tritici enhances resistance of wheat to stripe rust. Plant Biotechnol. J. 2018, 16, 797–807. [Google Scholar] [PubMed]
- Cheng, W.; Song, X.S.; Li, H.P.; Cao, L.H.; Sun, K.; Qiu, X.L.; Xu, Y.B.; Yang, P.; Huang, T.; Zhang, J.B. Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to Fusarium head blight and seedling blight in wheat. Plant Biotechnol. J. 2015, 13, 1335–1345. [Google Scholar] [CrossRef]
- Debroy, S.; Thilmony, R.; Kwack, Y.B.; Nomura, K.; He, S.Y. A family of conserved bacterial effectors inhibits salicylic acid-mediated basal immunity and promotes disease necrosis in plants. Proc. Natl. Acad. Sci. USA 2004, 101, 9927–9932. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Lin, N.C.; Martin, G.B. Two distinct Pseudomonas effector proteins interact with the pto kinase and activate plant immunity. Cell 2002, 109, 589–598. [Google Scholar] [CrossRef]
- Ahmed, A.A.; Pedersen, C.; Thordal-Christensen, H. The barley powdery mildew effector candidates CSEP0081 and CSEP0254 promote fungal infection success. PLoS ONE 2016, 11, e0157586. [Google Scholar] [CrossRef] [PubMed]
- Bourras, S.; McNally, K.E.; Ben-David, R.; Parlange, F.; Roffler, S.; Praz, C.R.; Oberhaensli, S.; Menardo, F.; Stirnweis, D.; Frenkel, Z. Multiple avirulence loci and allele-specific effector recognition control the Pm3 race-specific resistance of wheat to powdery mildew. Plant Cell 2015, 27, 2991–3012. [Google Scholar] [PubMed]
- Praz, C.R.; Bourras, S.; Zeng, F.; Sanchez-Martin, J.; Menardo, F.; Xue, M.; Yang, L.; Roffler, S.; Boni, R.; Herren, G. AvrPm2 encodes an RNase-like avirulence effector which is conserved in the two different specialized forms of wheat and rye powdery mildew fungus. New Phytol. 2017, 213, 1301–1314. [Google Scholar] [CrossRef] [PubMed]
- Pennington, H.G.; Gheorghe, D.M.; Damerum, A.; Pliego, C.; Spanu, P.D.; Cramer, R.; Bindschedler, L.V. Interactions between the powdery mildew effector BEC1054 and barley proteins identify candidate host targets. J. Proteome Res. 2016, 15, 826–839. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yang, L.; Zhou, X.; Zhou, M.; Lu, Y.; Ma, L.; Ma, H.; Zhang, Z. Transgenic wheat expressing Thinopyrum intermedium MYB transcription factor TiMYB2R-1 shows enhanced resistance to the take-all disease. J. Exp. Bot. 2013, 64, 2243–2253. [Google Scholar] [CrossRef] [PubMed]
- Rong, W.; Qi, L.; Wang, J.; Du, L.; Xu, H.; Wang, A.; Zhang, Z. Expression of a potato antimicrobial peptide SN1 increases resistance to take-all pathogen Gaeumannomyces graminis var. tritici in transgenic wheat. Funct. Integr. Genom. 2013, 13, 403–409. [Google Scholar] [CrossRef]
- Wang, A.; Wei, X.; Rong, W.; Dang, L.; Du, L.P.; Qi, L.; Xu, H.J.; Shao, Y.; Zhang, Z. GmPGIP3 enhanced resistance to both take-all and common root rot diseases in transgenic wheat. Funct. Integr. Genom. 2015, 15, 375–381. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, X.; Liu, D.; Xu, W.; Wise, R.; Shen, Q.H. The miR9863 family regulates distinct Mla alleles in barley to attenuate NLR receptor-triggered disease resistance and cell-death signaling. PLoS Genet. 2014, 10, e1004755. [Google Scholar] [CrossRef]
- Walker, A.S.; Bouguennec, A.; Confais, J.; Morgant, G.; Leroux, P. Evidence of host-range expansion from new powdery mildew (Blumeria graminis) infections of triticale (×Triticosecale) in France. Plant Pathol. 2011, 60, 207–220. [Google Scholar] [CrossRef]
- Daval, S.; Lebreton, L.; Gazengel, K.; Boutin, M.; Guillerm-Erckelboudt, A.Y.; Sarniguet, A. The biocontrol bacterium Pseudomonas fluorescens Pf29Arp strain affects the pathogenesis-related gene expression of the take-all fungus Gaeumannomyces graminis var. tritici on wheat roots. Mol. Plant Pathol. 2011, 12, 839–854. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 2010, 11, 1187–1194. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Primers Purpose |
|---|---|---|
| BEC1019-F | ATGCAGTCTGTATTGCTTTT | Homolog gene cloning of BEC1019 |
| BEC1019-R | CTAGACACAATGAACCTCGC | |
| BEC1019-F1 | GGGGACAGTTTGTACAAAAAAGCAGGCTTCATGCAGTCTGTATTGCTTTT | Construction of CTAPi-GW-3HA-BEC1019 vector |
| BEC1019-R1 | GGGGACCACTTTGTACAAGAAAGCTGGGTGACACAATGAACCTCGCCAT | |
| BEC1019-F2 | GGGGACAGTTTGTACAAAAAAGCAGGCTTCGTGATGACCCGGACAAAA | Construction of pCAMBIA2301-BEC1019-RNAi vector |
| BEC1019-R2 | GGGGACCACTTTGTACAAGAAAGCTGGGTAGGGCATCTTGGTAACCA | |
| BEC1019-F3 | GGGGACAGTTTGTACAAAAAAGCAGGCTTCGTGATGACCCGGACAAAA | Construction of CTAPi-GW-3HA-BEC1019-RNAi vector |
| BEC1019-R3 | GGGGACCACTTTGTACAAGAAAGCTGGGTAGGGCATCTTGGTAACCA | |
| BEC1019-F4 | TCATGTGGACATCGTCGGTC | q RT-PCR analysis of BEC1019 over-expression wheat lines |
| BEC1019-R4 | CACGCTGATGTCAAACGCAT | |
| BEC1019-F5 | AATGTGCAACCGAGAACCGA | q RT-PCR analysis of BEC1019 silencing wheat lines |
| BEC1019-R5 | TCCTCCAAAGGAAGCCGTTC | |
| TaPR2-F | CCGGCCATACTACCCGGC | q RT-PCR analysis for TaPR2 |
| TaPR2-R | ACACCTTGATGGCGCTGAGA | |
| TaPR10-F | ACGGAGCGGATGTGGAAG | q RT-PCR analysis for TaPR10 |
| TaPR10-R | GCCACCTGCGACTTGAGC | |
| TaActin-F | CCAGGTATCGCTGACCGTAT | Reference gene of wheat |
| TaActin-R | GCTGAGTGAGGCTAGGATGG | |
| Bgt-EF1a-F | GTCGGATTTAACCCCAAGGT | Reference gene of Bgt |
| Bgt-EF1a-R | TTTATCGGTAGGGCGACTTG | |
| Ggt-18S rRNA-F | CGAACTCGGTCGTTTAGAGG | Reference gene of Ggt |
| Ggt-18S rRNA-R | GGTATGTTCACAGGGGTTGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Xu, K.; Yu, D.; Liu, Z.; Peng, C.; Li, X.; Zhang, J.; Dong, Y.; Zhang, Y.; Tian, P.; et al. The Highly Conserved Barley Powdery Mildew Effector BEC1019 Confers Susceptibility to Biotrophic and Necrotrophic Pathogens in Wheat. Int. J. Mol. Sci. 2019, 20, 4376. https://doi.org/10.3390/ijms20184376
Zhang Y, Xu K, Yu D, Liu Z, Peng C, Li X, Zhang J, Dong Y, Zhang Y, Tian P, et al. The Highly Conserved Barley Powdery Mildew Effector BEC1019 Confers Susceptibility to Biotrophic and Necrotrophic Pathogens in Wheat. International Journal of Molecular Sciences. 2019; 20(18):4376. https://doi.org/10.3390/ijms20184376
Chicago/Turabian StyleZhang, Yi, Kedong Xu, Deshui Yu, Zhihui Liu, Chunfeng Peng, Xiaoli Li, Ju Zhang, Yinghui Dong, Yazhen Zhang, Pan Tian, and et al. 2019. "The Highly Conserved Barley Powdery Mildew Effector BEC1019 Confers Susceptibility to Biotrophic and Necrotrophic Pathogens in Wheat" International Journal of Molecular Sciences 20, no. 18: 4376. https://doi.org/10.3390/ijms20184376
APA StyleZhang, Y., Xu, K., Yu, D., Liu, Z., Peng, C., Li, X., Zhang, J., Dong, Y., Zhang, Y., Tian, P., Guo, T., & Li, C. (2019). The Highly Conserved Barley Powdery Mildew Effector BEC1019 Confers Susceptibility to Biotrophic and Necrotrophic Pathogens in Wheat. International Journal of Molecular Sciences, 20(18), 4376. https://doi.org/10.3390/ijms20184376