Identification of Cerebrospinal Fluid Metabolites as Biomarkers for Enterovirus Meningitis

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Demographic and Standard Blood and CSF Diagnostic Parameters
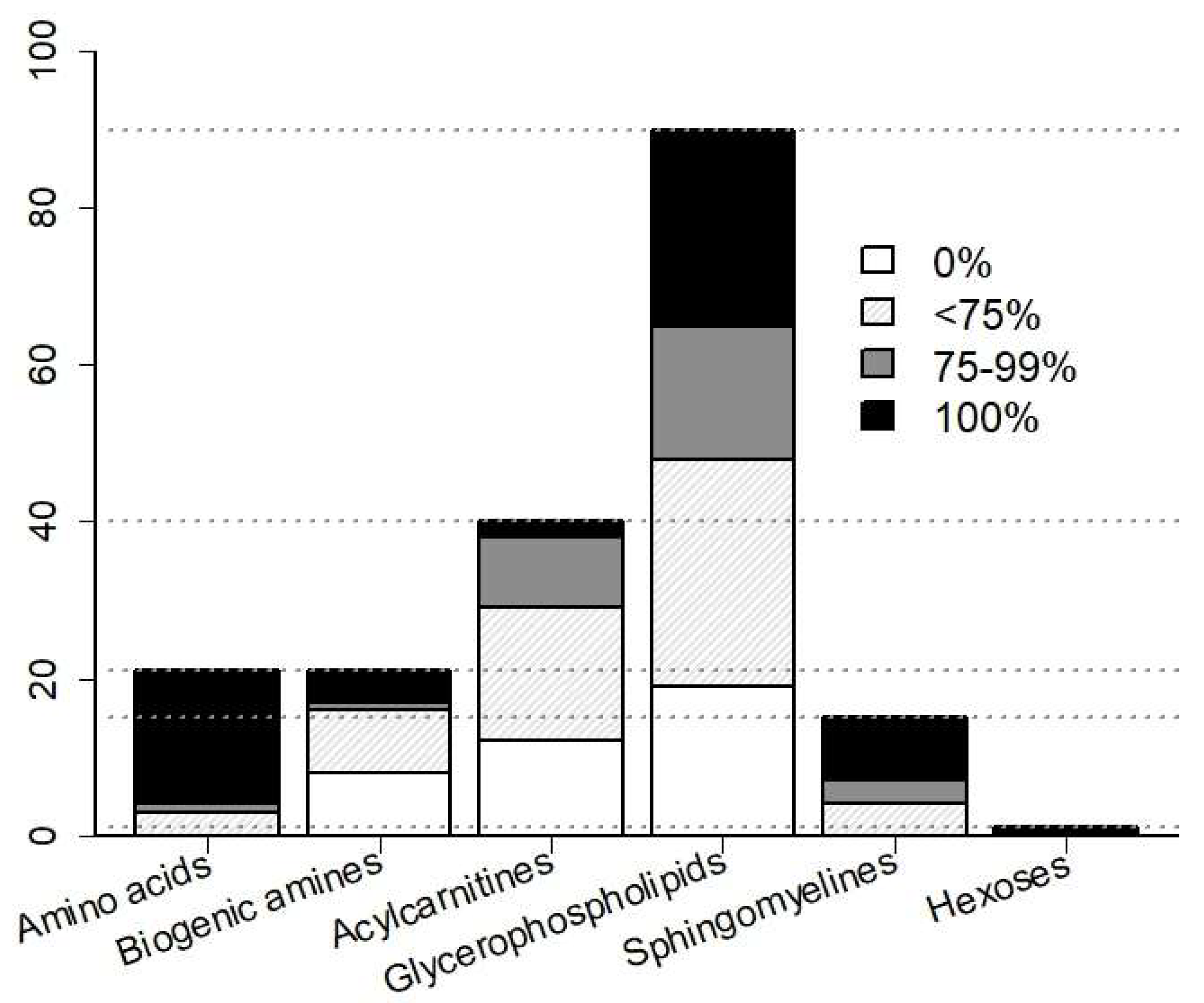
2.2. Efficiency of CSF Metabolite Detection by Mass Spectrometry
2.3. Metabolite Reprogramming in CSF in Enteroviral Meningitis
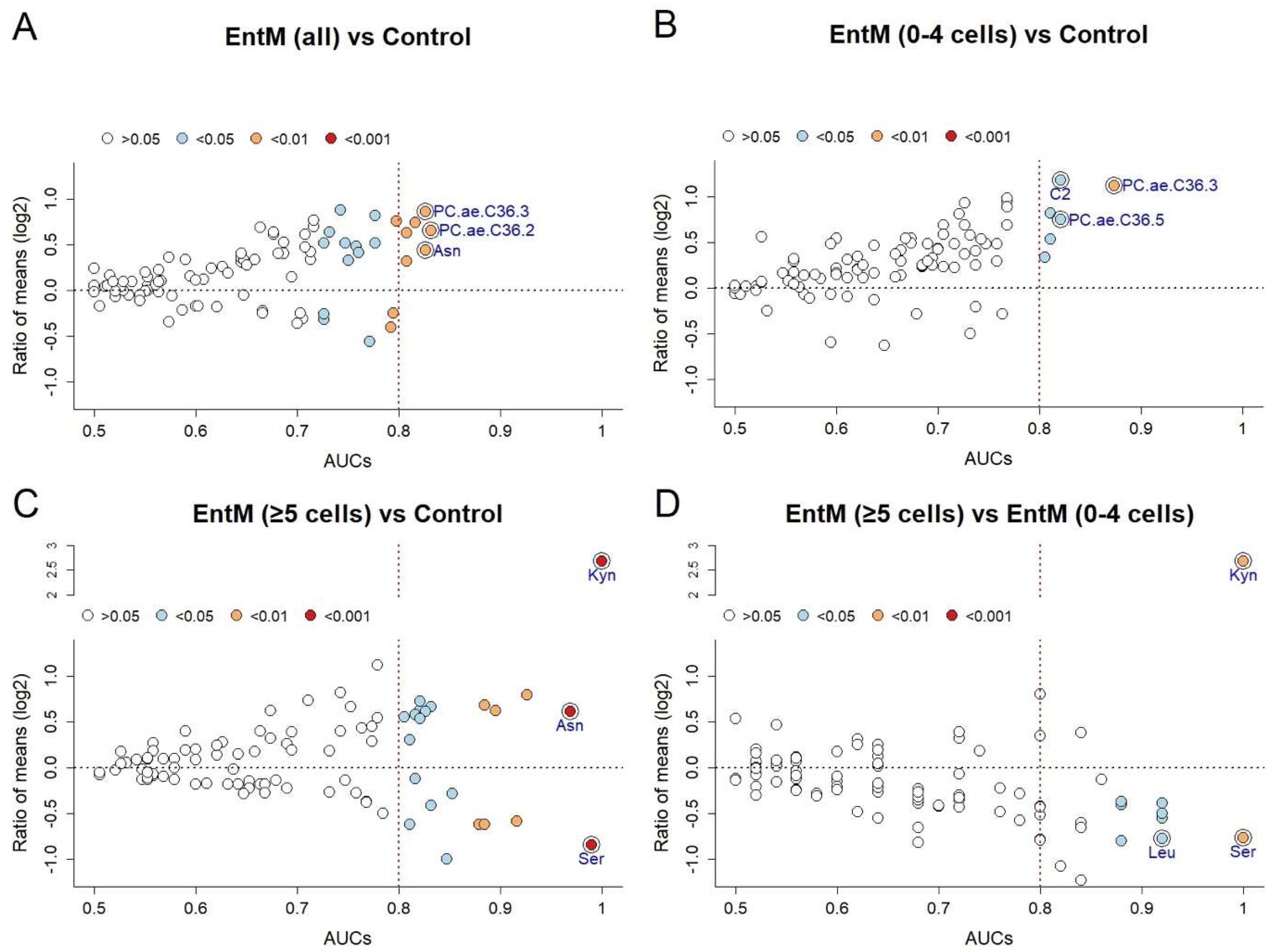
2.4. Selection of Specific Biomarkers by Internal Cross-Validation
2.5. Identification of Optimal Metabolite Classifiers by Random Forest Construction
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Metabolite Profiling
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AUC | Area under the curve |
| BCB | Blood-CSF-Barrier |
| CI | Confidence interval |
| CRP | C-reactive protein |
| CSF | Cerebrospinal fluid |
| CNS | central nervous system |
| Cho2p | Choline requiring protein 2 |
| ER | Emergency room |
| EntM | Enterovirus meningitis |
| HSV | Herpes simplex virus |
| IgG | Immunoglobulin G |
| LOD | Limit of detection |
| mTOR | Mechanistic target of rapamycin |
| NPH | Normal pressure hydrocephalus |
| PC | Phosphatidylcholine |
| PCR | Polymerase chain reaction |
| RNA | Ribonucleic acid |
| ROC | Receiver operating characteristic |
| VZV | Varicella zoster virus |
References
- Hoffman, O.; Weber, R.J. Pathophysiology and treatment of bacterial meningitis. Ther. Adv. Neurol. Disord. 2009, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bartt, R. Acute bacterial and viral meningitis. Continuum 2012, 18, 1255–1270. [Google Scholar] [CrossRef] [PubMed]
- De Ory, F.; Avellon, A.; Echevarria, J.E.; Sanchez-Seco, M.P.; Trallero, G.; Cabrerizo, M.; Casas, I.; Pozo, F.; Fedele, G.; Vicente, D.; et al. Viral infections of the central nervous system in Spain: A prospective study. J. Med. Virol. 2013, 85, 554–562. [Google Scholar] [CrossRef]
- Dumaidi, K.; Frantzidou, F.; Papa, A.; Diza, E.; Antoniadis, A. Enterovirus meningitis in Greece from 2003–2005: Diagnosis, CSF laboratory findings, and clinical manifestations. J. Clin. Lab. Anal. 2006, 20, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Majed, B.; Zephir, H.; Pichonnier-Cassagne, V.; Yazdanpanah, Y.; Lestavel, P.; Valette, P.; Vermersch, P. Lumbar punctures: Use and diagnostic efficiency in emergency medical departments. Int. J. Emerg. Med. 2009, 2, 227–235. [Google Scholar] [CrossRef]
- Glimaker, M.; Johansson, B.; Grindborg, O.; Bottai, M.; Lindquist, L.; Sjolin, J. Adult bacterial meningitis: Earlier treatment and improved outcome following guideline revision promoting prompt lumbar puncture. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 60, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Ostergaard, A.A.; Sydenham, T.V.; Nybo, M.; Andersen, A.B. Cerebrospinal fluid pleocytosis level as a diagnostic predictor? A cross-sectional study. BMC Clin. Pathol. 2017, 17, 15. [Google Scholar] [CrossRef] [Green Version]
- Ahlbrecht, J.; Hillebrand, L.K.; Schwenkenbecher, P.; Ganzenmueller, T.; Heim, A.; Wurster, U.; Stangel, M.; Suhs, K.W.; Skripuletz, T. Cerebrospinal fluid features in adults with enteroviral nervous system infection. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2018, 68, 94–101. [Google Scholar] [CrossRef]
- Debiasi, R.L.; Tyler, K.L. Molecular methods for diagnosis of viral encephalitis. Clin. Microbiol. Rev. 2004, 17, 903–925, table of contents. [Google Scholar] [CrossRef]
- Ihekwaba, U.K.; Kudesia, G.; McKendrick, M.W. Clinical features of viral meningitis in adults: Significant differences in cerebrospinal fluid findings among herpes simplex virus, varicella zoster virus, and enterovirus infections. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2008, 47, 783–789. [Google Scholar] [CrossRef]
- Patriquin, G.; Hatchette, J.; Forward, K. Clinical presentation of patients with aseptic meningitis, factors influencing treatment and hospitalization, and consequences of enterovirus cerebrospinal fluid polymerase chain reaction testing. Can. J. Infect. Dis. Med Microbiol. J. Can. Des Mal. Infect. Et De La Microbiol. Med. 2012, 23, e1–e5. [Google Scholar] [CrossRef]
- Kuhn, M.; Suhs, K.W.; Akmatov, M.K.; Klawonn, F.; Wang, J.; Skripuletz, T.; Kaever, V.; Stangel, M.; Pessler, F. Mass-spectrometric profiling of cerebrospinal fluid reveals metabolite biomarkers for CNS involvement in varicella zoster virus reactivation. J. Neuroinflamm. 2018, 15, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000. [Google Scholar]
- Demler, O.V.; Pencina, M.J.; D’Agostino, R.B., Sr. Impact of correlation on predictive ability of biomarkers. Stat. Med. 2013, 32, 4196–4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, Z.; Chukkapalli, V.; Nchoutmboube, J.A.; Li, J.; Randall, G.; Belov, G.A.; Wang, X. Positive-strand RNA viruses stimulate host phosphatidylcholine synthesis at viral replication sites. Proc. Natl. Acad. Sci. USA 2016, 113, E1064–E1073. [Google Scholar] [CrossRef] [PubMed]
- Boumann, H.A.; Chin, P.T.; Heck, A.J.; De Kruijff, B.; De Kroon, A.I. The yeast phospholipid N-methyltransferases catalyzing the synthesis of phosphatidylcholine preferentially convert di-C16:1 substrates both in vivo and in vitro. J. Biol. Chem. 2004, 279, 40314–40319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Randell, E.W.; Sun, G.; Likhodii, S.; Liu, M.; Furey, A.; Zhai, G. Hyperglycemia-related advanced glycation end-products is associated with the altered phosphatidylcholine metabolism in osteoarthritis patients with diabetes. PLoS ONE 2017, 12, e0184105. [Google Scholar] [CrossRef] [PubMed]
- Cambiaghi, A.; Pinto, B.B.; Brunelli, L.; Falcetta, F.; Aletti, F.; Bendjelid, K.; Pastorelli, R.; Ferrario, M. Characterization of a metabolomic profile associated with responsiveness to therapy in the acute phase of septic shock. Sci. Rep. 2017, 7, 9748. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhou, B.; Su, M.; Baxter, S.; Zheng, X.; Zhao, X.; Yen, Y.; Jia, W. Mass spectrometry-based quantitative metabolomics revealed a distinct lipid profile in breast cancer patients. Int. J. Mol. Sci. 2013, 14, 8047–8061. [Google Scholar] [CrossRef]
- Mastrokolias, A.; Pool, R.; Mina, E.; Hettne, K.M.; van Duijn, E.; van der Mast, R.C.; van Ommen, G.; t Hoen, P.A.; Prehn, C.; Adamski, J.; et al. Integration of targeted metabolomics and transcriptomics identifies deregulation of phosphatidylcholine metabolism in Huntington’s disease peripheral blood samples. Metab. Off. J. Metab. Soc. 2016, 12, 137. [Google Scholar] [CrossRef]
- Yetisen, A.K.; Akram, M.S.; Lowe, C.R. Paper-based microfluidic point-of-care diagnostic devices. Lab Chip 2013, 13, 2210–2251. [Google Scholar] [CrossRef]
- Boulos, A.; Fairley, D.; McKenna, J.; Coyle, P. Evaluation of a rapid antigen test for detection of Streptococcus pneumoniae in cerebrospinal fluid. J. Clin. Pathol. 2017, 70, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Krall, A.S.; Xu, S.; Graeber, T.G.; Braas, D.; Christofk, H.R. Asparagine promotes cancer cell proliferation through use as an amino acid exchange factor. Nat. Commun. 2016, 7, 11457. [Google Scholar] [CrossRef] [PubMed]
- Perez-Torres, I.; Zuniga-Munoz, A.M.; Guarner-Lans, V. Beneficial Effects of the Amino Acid Glycine. Mini Rev. Med. Chem. 2017, 17, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Corcuera, B.; Geerlings, A.; Aragon, C. Glycine neurotransmitter transporters: An update. Mol. Membr. Biol. 2001, 18, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef]
- Adams, R.D.; Fisher, C.M.; Hakim, S.; Ojemann, R.G.; Sweet, W.H. Symptomatic Occult Hydrocephalus with “Normal” Cerebrospinal-Fluid Pressure. A Treatable Syndrome. N. Engl. J. Med. 1965, 273, 117–126. [Google Scholar] [CrossRef]
- Novoselova, N.; Wang, J.; Pessler, F.; Klawonn, F. Biocomb: Feature Selection and Classification with the Embedded Validation Procedures for Biomedical Data Analysis. R Package Version 0.4. Available online: https://CRAN.R-project.org/package=Biocomb (accessed on 1 October 2018).




{kind=link}
{kind=link}
{kind=link}
| Control | EntM | ||||||
|---|---|---|---|---|---|---|---|
| Subgroup | p Value §, Control vs. | ||||||
| (n = 19) | All (n = 10) | 0–4 Cells (n = 5) | ≥5 Cells (n = 5) | EntM All | EntM 0–4 Cells | EntM ≥5 Cells | |
| Age (Years) | |||||||
| Median | 48 (22–77) | 32.5 (22–76) | 47 (33–76) | 30 (22–32) | 0.22 | 0.52 | 0.01 |
| Mean (SD) | 48 (16.1) | 41 (19.4) | 55 (19) | 27.8 (4.5) | |||
| Sex | |||||||
| Female | 58% (11) | 40% (4) | 40% (2) | 40% (2) | 0.37 | 0.48 | 0.48 |
| Male | 42% (8) | 60% (6) | 60% (3) | 60% (3) | |||
| Blood Parameter | |||||||
| Leukocytes (1000/µL) (normal range 3.6–10.5) | |||||||
| Median | 7.4 (4.6–11.9) | 7 (4–14) | 6.6 (5.1–9.8) | 7.3 (4–14) | 0.55 | 0.5 | 0.80 |
| Mean (SD) | 7.8 (2.3) | 7.4 (2.9) | 7.1 (1.83) | 7.7 (3.8) | |||
| CRP (mg/L) (normal range 0–5) | |||||||
| Median | 3 (1–31) | 4 (1–39) | 4 (1–17) | 4 (1–39) | 0.09 | 0.18 | 0.20 |
| Mean (SD) | 5.4 (8.3) | 9.5 (11.6) | 7.4 (6.5) | 11.6 (15.8) | |||
| CSF Parameter | |||||||
| Cell count (1/µL) (normal range 0–4) | |||||||
| Median | 1.3 (0.3–4) | 9.2 (0.7–619) | 1.7 (0.7–4) | 97.3 (14.3–619) | 0.008 | n/a | n/a |
| Mean (SD) | 1.6 (1.1) | 129.1 (235.3) | 2 (1.35) | 256.2 (290.3) | |||
| Protein (g/L) (0.17–0.52) | |||||||
| Median | 0.39 (0.26–0.83) | 0.51 (0.24–0.98) | 0.52 (0.24–0.98) | 0.54 (0.46–0.75) | 0.14 | 0.41 | 0.15 |
| Mean (SD) | 0.45 (0.15) | 0.57 (0.19) | 0.58 (0.51) | 0.56 (0.16) | |||
| Lactate (mmol/L) (normal range 1.1–1.9) | |||||||
| Median | 1.57 (1.2–2.1) | 1.88 (1.55–3.55) | 1.7 (1.55–2.18) | 2.15 (1.84–3.55) | 0.007 | 0.24 | 0.003 |
| Mean (SD) | 1.6 (0.25) | 2.1 (0.62) | 1.76 (0.5) | 2.43 (0.71) | |||
| IgG Index | |||||||
| Median | 0.49 (0.43–0.6) | 0.54 (0.47–0.63) | 0.55 (0.49–0.63) | 0.52 (0.47–0.54) | 0.041 | 0.035 | 0.28 |
| Mean (SD) | 0.5 (0.05) | 0.53 (0.05) | 0.56 (0.05) | 0.51 (0.03) | |||
| Blood-CSF-Barrier Dysfunction | |||||||
| No | 58% (11) | 20% (2) | 40% (2) | 0 | 0.037 | 0.31 | 0.023 |
| Light | 42% (8) | 70% (7) | 40% (2) | 100% (5) | |||
| Moderate | 0 | 10% (1) | 20% (1) | ||||
| Severe | 0 | 0 | 0 | 0 | |||
| EntM (all) vs. Control | AUC | p-Value | Lower CI | Ratio of Means | Selection Frequency |
|---|---|---|---|---|---|
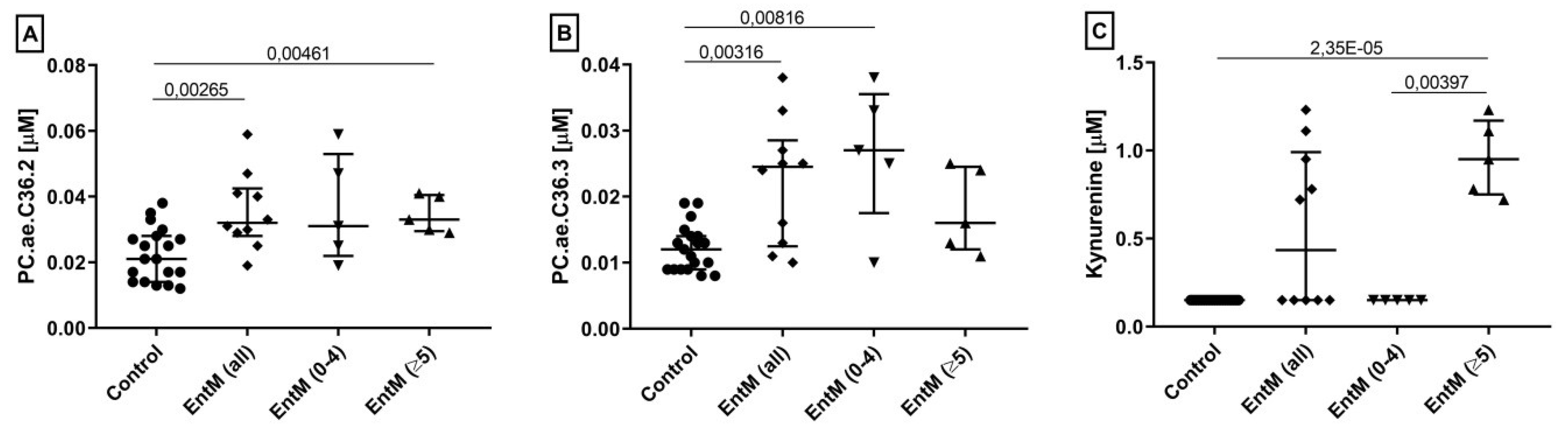
| PC.ae.C36.2* | 0.832 | 0.00265 | 0.653 | 1.58 | 1.000 |
| Asn | 0.826 | 0.00316 | 0.621 | 1.35 | 1.000 |
| PC.ae.C36.3 | 0.826 | 0.00316 | 0.634 | 1.82 | 1,000 |
| Gly | 0.816 | 0.00445 | 0.601 | 1.67 | 1.000 |
| C5.1 | 0.808 | 0.00568 | 0.656 | 1.25 | 0.966 |
| PC.aa.C34.2 | 0.808 | 0.00568 | 0.592 | 1.55 | 1.000 |
| PC.ae.C38.5 | 0.797 | 0.00779 | 0.647 | 1.69 | 0.931 |
| H1 | 0.795 | 0.00843 | 0.625 | 0.84 | 0.776 |
| Ser | 0.792 | 0.00907 | 0.574 | 0.75 | 0.707 |
| PC.aa.C34.3 | 0.776 | 0.01406 | 0.587 | 1.43 | 0.397 |
| Ent M (0-4 cells) vs. Control | |||||
| PC.ae.C36.3* | 0.874 | 0.00816 | 0.590 | 0.46 | 1.000 |
| C2 | 0.821 | 0.02675 | 0.548 | 0.44 | 1.000 |
| PC.ae.C36.5 | 0.821 | 0.02675 | 0.556 | 0.60 | 0.958 |
| C0 | 0.811 | 0.03287 | 0.530 | 0.69 | 1.000 |
| PC.aa.C36.5 | 0.811 | 0.03287 | 0.593 | 0.57 | 0.958 |
| C5.1 | 0.805 | 0.03618 | 0.565 | 0.79 | 0.917 |
| Ent M (≥5 cells) vs. Control | |||||
| Kyn* | 1.000 | 2.35 × 10−5 | 1.000 | 6.39 | 1.000 |
| Ser | 0.989 | 7.06 × 10−5 | 0.937 | 0.56 | 1.000 |
| Asn | 0.968 | 0.00026 | 0.872 | 1.53 | 1.000 |
| Gly | 0.926 | 0.00167 | 0.748 | 1.73 | 1.000 |
| Lys | 0.916 | 0.00240 | 0.792 | 0.66 | 0.958 |
| PC.ae.C36.2 | 0.895 | 0.00461 | 0.738 | 1.54 | 0.958 |
| PC.aa.C34.2 | 0.884 | 0.00619 | 0.710 | 1.61 | 1.000 |
| Tyr | 0.884 | 0.00619 | 0.731 | 0.65 | 0.925 |
| Leu | 0.879 | 0.00706 | 0.654 | 0.65 | 0.883 |
| H1 | 0.853 | 0.01360 | 0.669 | 0.82 | 0.467 |
| EntM (≥5 cells) vs. EntM (0–4 cells) | |||||
| Kyn* | 1.000 | 0.00397 | 1.000 | 6.39 | 1.000 |
| Ser | 1.000 | 0.00397 | 1.000 | 0.59 | 1.000 |
| Leu | 0.920 | 0.02381 | 0.700 | 0.58 | 0.940 |
| Tyr | 0.920 | 0.02381 | 0.667 | 0.68 | 0.900 |
| PC.aa.C32.0 | 0.920 | 0.02381 | 0.714 | 0.76 | 0.940 |
| PC.aa.C34.1 | 0.920 | 0.02381 | 0.672 | 0.71 | 0.940 |
| Phe | 0.880 | 0.04365 | 0.577 | 0.77 | 0.333 |
| Val | 0.880 | 0.04365 | 0.583 | 0.76 | 0.407 |
| Creatinine | 0.880 | 0.04365 | 0.577 | 0.77 | 0.367 |
| PC.ae.C36.5 | 0.880 | 0.04365 | 0.577 | 0.57 | 0.433 |
| Controls vs. | |||
|---|---|---|---|
| EntM (All) | EntM (0–4 Cells) | EntM (≥5 Cells) | |
| Standard CSF Parameter | |||
| Leukocyte count | 0.80 ** | n/a | n/a |
| Protein concentration | 0.67 | 0.62 | 0.72 |
| IgG-index | 0.73 * | 0.81* | 0.66 |
| Lactate | 0.81 ** | 0.67 | 0.94 ** |
| BCB dysfunction | 0.71 | 0.63 | 0.79 * |
| Best Single Metabolites 1 | |||
| Best internally validated marker | PC.ae.C36.2 | PC.ae.C36.3 | Kyn |
| AUC | 0.83 ** | 0.87 ** | 1.0 *** |
| Best Metabolite Classifier 2 | |||
| No. of markers | 3 | 3 | 1 |
| Markers (frequency) | Asn (1.0) Gly (1.0) PC.ae.C36.3 (1.0) | PC.ae.C36.3 (1.0) PC.ae.C36.5 (0.96) PC.ae.C38.5 (0.6) | Kyn (1.0) |
| AUC (95% CI) | 0.92 *** (0.61–1.0) | 0.99 *** (0.53–1.0) | 1.0 *** (1.0–1.0) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ratuszny, D.; Sühs, K.-W.; Novoselova, N.; Kuhn, M.; Kaever, V.; Skripuletz, T.; Pessler, F.; Stangel, M. Identification of Cerebrospinal Fluid Metabolites as Biomarkers for Enterovirus Meningitis. Int. J. Mol. Sci. 2019, 20, 337. https://doi.org/10.3390/ijms20020337
Ratuszny D, Sühs K-W, Novoselova N, Kuhn M, Kaever V, Skripuletz T, Pessler F, Stangel M. Identification of Cerebrospinal Fluid Metabolites as Biomarkers for Enterovirus Meningitis. International Journal of Molecular Sciences. 2019; 20(2):337. https://doi.org/10.3390/ijms20020337
Chicago/Turabian StyleRatuszny, Dominica, Kurt-Wolfram Sühs, Natalia Novoselova, Maike Kuhn, Volkhard Kaever, Thomas Skripuletz, Frank Pessler, and Martin Stangel. 2019. "Identification of Cerebrospinal Fluid Metabolites as Biomarkers for Enterovirus Meningitis" International Journal of Molecular Sciences 20, no. 2: 337. https://doi.org/10.3390/ijms20020337
APA StyleRatuszny, D., Sühs, K. -W., Novoselova, N., Kuhn, M., Kaever, V., Skripuletz, T., Pessler, F., & Stangel, M. (2019). Identification of Cerebrospinal Fluid Metabolites as Biomarkers for Enterovirus Meningitis. International Journal of Molecular Sciences, 20(2), 337. https://doi.org/10.3390/ijms20020337