Epigenetic Modifications of the Liver Tumor Cell Line HepG2 Increase Their Drug Metabolic Capacity

 ,
,  ,
, 

 and
and
Abstract
:1. Introduction
2. Results
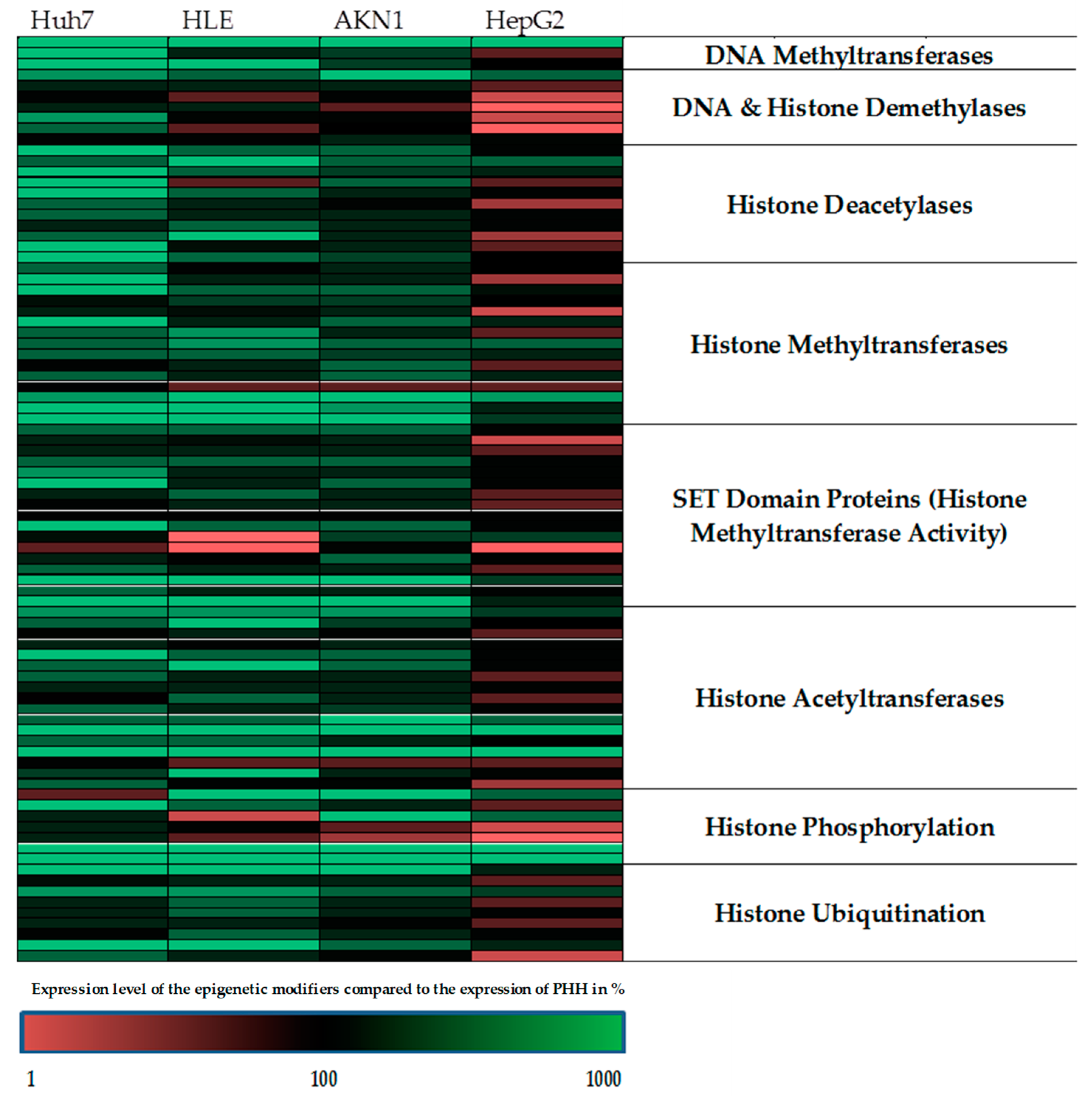
2.1. The Regulation of the Epigenetic Enzymes in HepG2 is Most Closely Comparable to the Expression of Primary Human Hepatocytes
2.2. Treatment of HepG2 with Epigenetic Modifying Compounds Revealed a Positive Impact on the Expression of Genes From Xenobiotic Metabolism
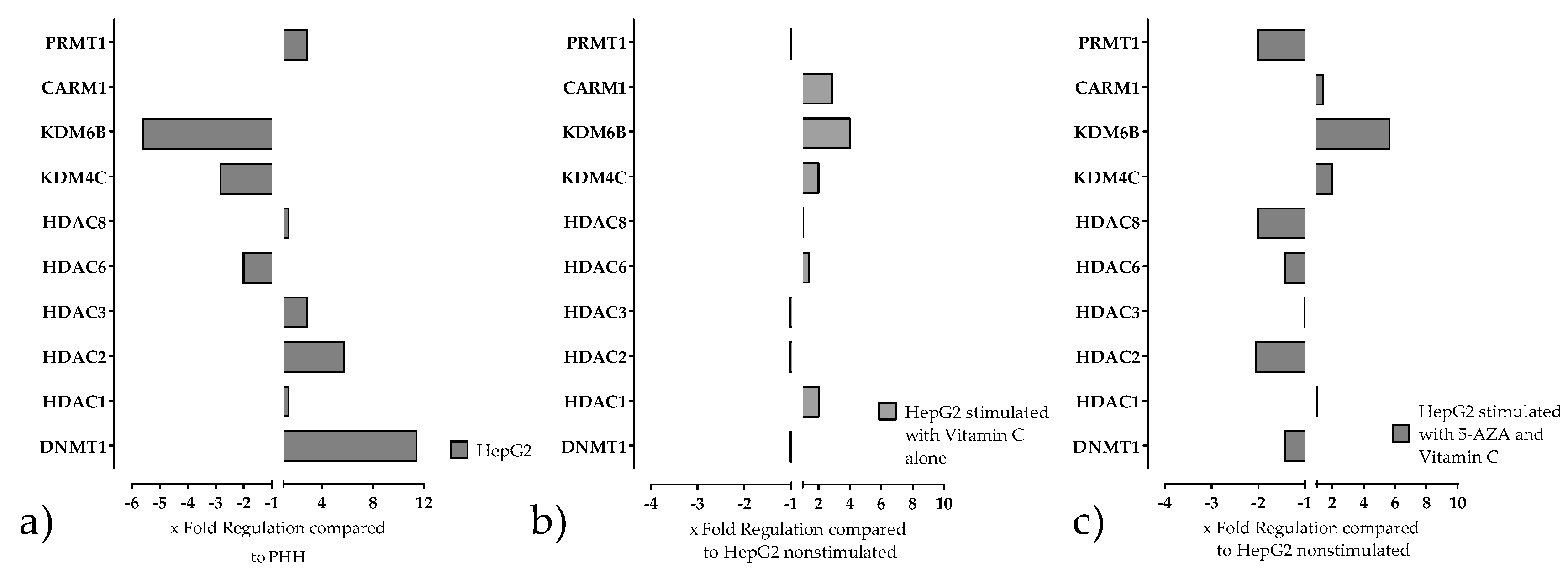
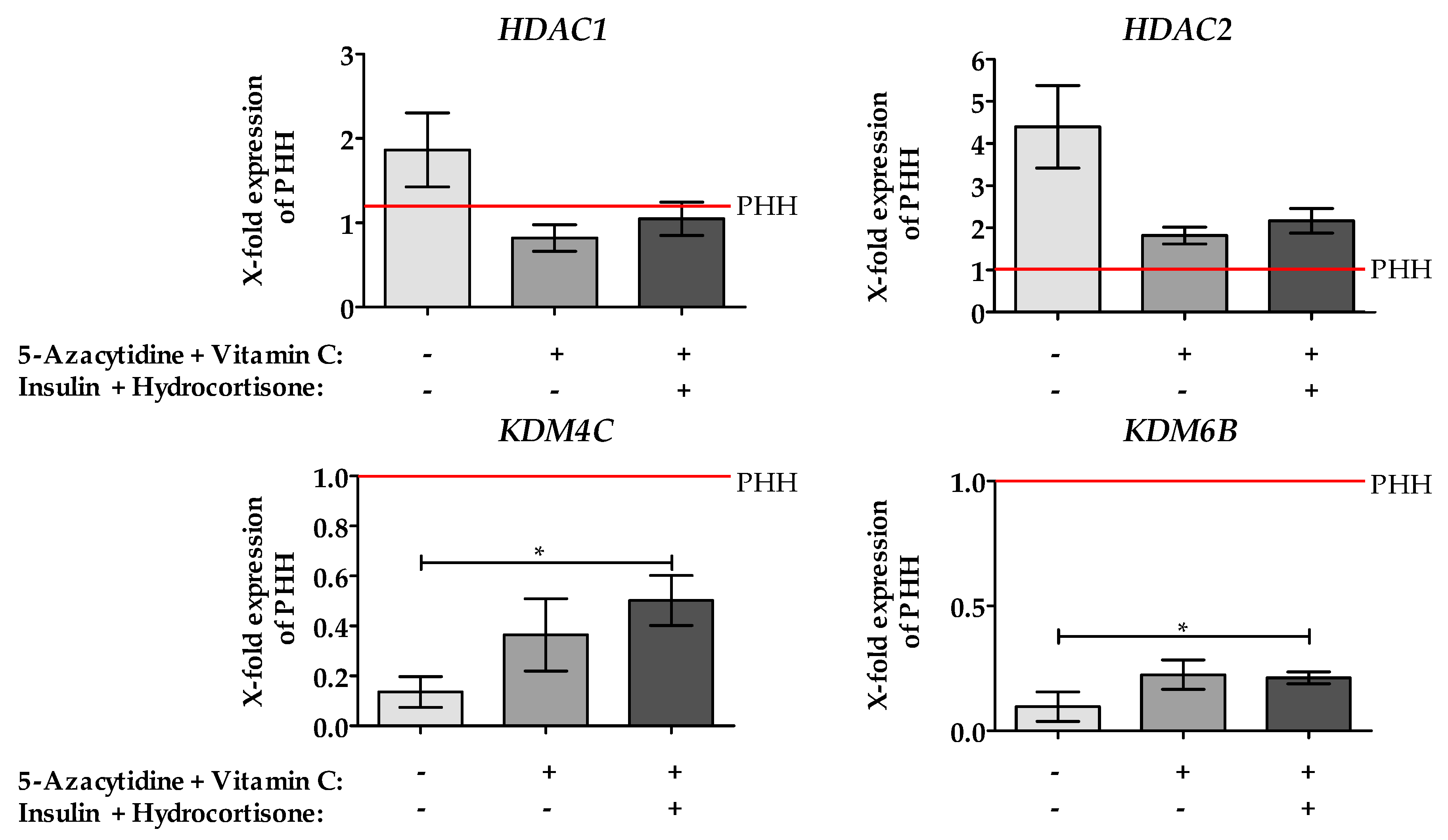
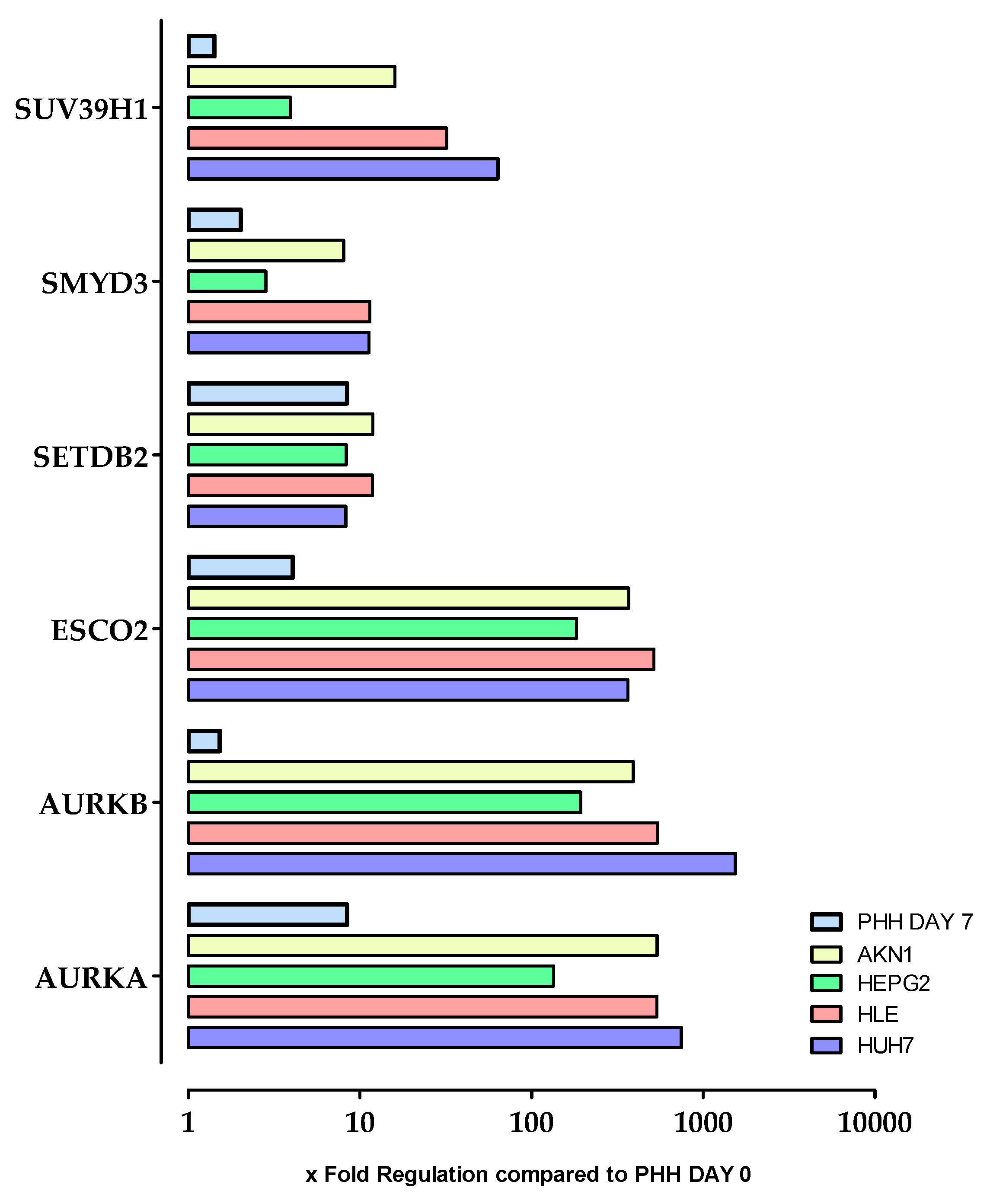
2.3. Stimulation of HepG2 Cells with Epigenetic Modifying Compounds Result in Changes in Gene Expression of Epigenetic Modifying Enzymes
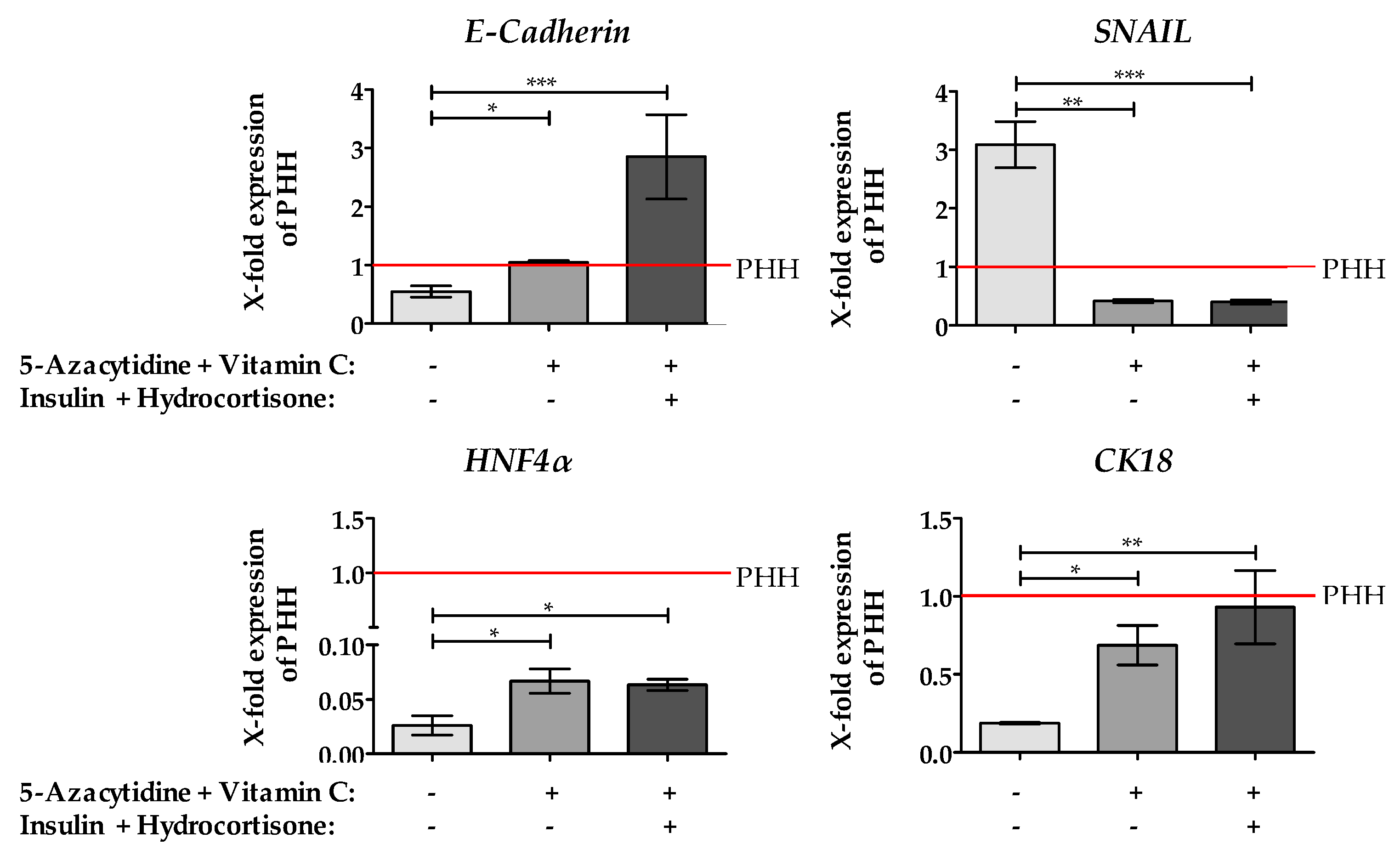
2.4. Stimulation of HepG2 Cells with 5-AZA Plus Vitamin C Led to the Downregulation of the EMT Marker Gene SNAIL, an Increase of Epithelial Marker Genes and the Hepatic Key Regulator HNF4α
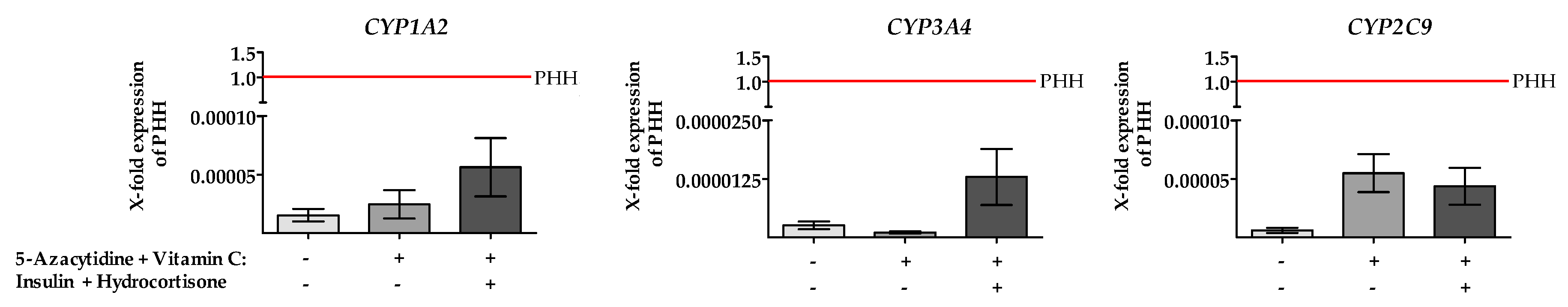
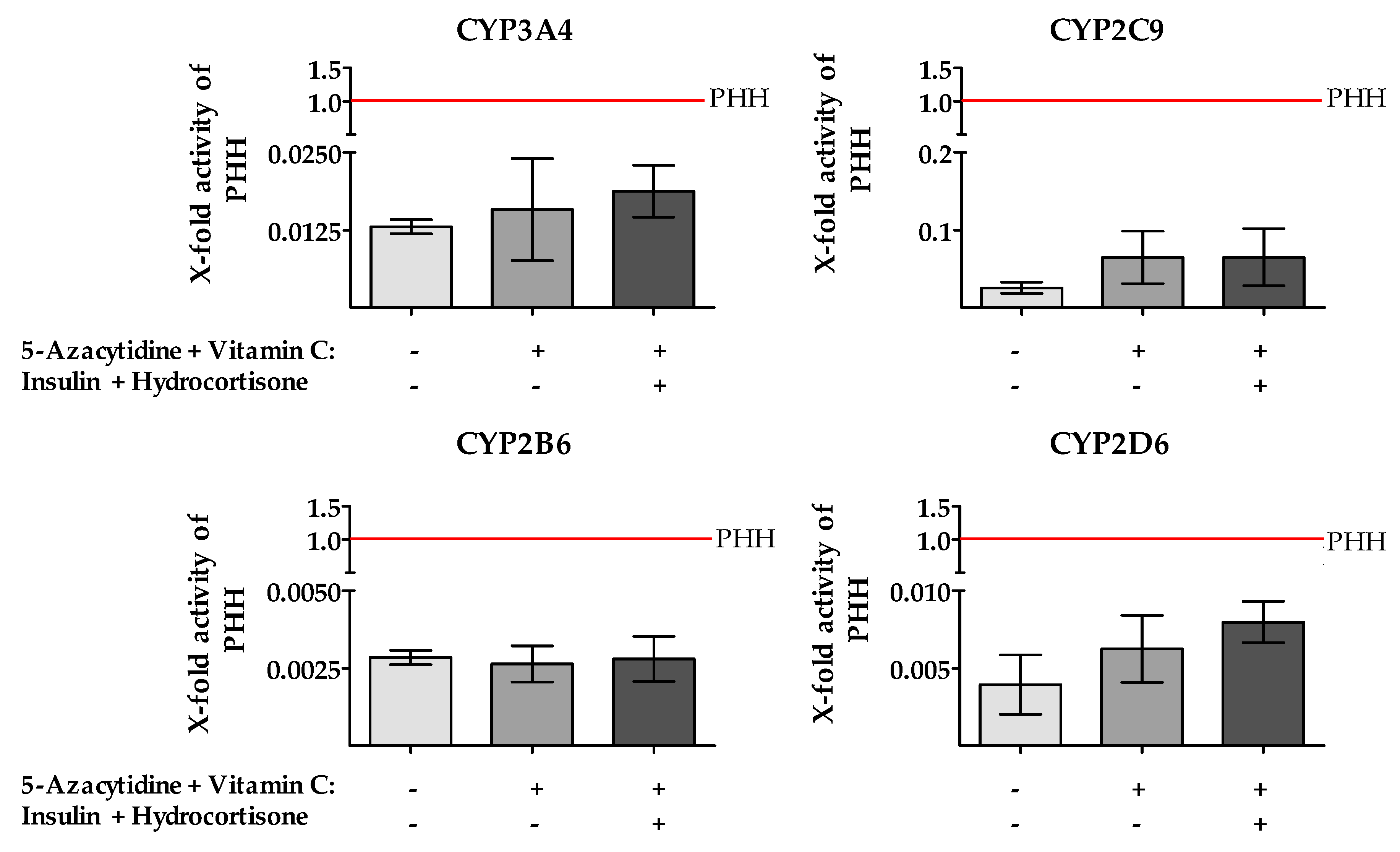
2.5. Treatment of HepG2 Cells with 5-AZA Plus Vitamin C in Combination with Insulin and Hydrocortisone Resulted in an Increased CYP450 Gene Expression and Enzyme Activity
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Isolation of Primary Human Hepatocytes
4.3. Culture of Primary Cells and Cell Lines
4.3.1. Primary Cells
4.3.2. Cell Lines
4.4. Epigenetic Modification Array
4.5. cDNA Synthesis and RT-PCR
4.6. CYP Activity Measurement
4.7. SRB Staining for Normalisation of the Results
4.8. Statistic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CYP | Cytochrome P450 |
| cDNA | Complementary DNA |
| EMT | Epithelial–Mesenchymal Transition |
| PHH | primary human hepatocytes |
| SRB | Sulforhodamine B |
| 5-AZA | 5-aza-2′-deoxycytidine |
| CK18 | Cytokeratin 18 |
| HNF4α | Hepatocyte nuclear factor 4α |
| SNAIL | Snail1 |
| KDM6B | Lysine Demethylase 6B |
| KDM4C | Lysine Demethylase 4C |
| JCRB | Japanese Collection of Research Bioresources Cell Bank |
| 5-AZA-dC | 5-Aza-2′-deoxycytidine |
| HCC | hepatocellular carcinoma |
| HDAC | Histone deacetylase |
| HDACi | Histone deacetylase inhibitor |
| DNMT | DNA methyltransferases |
| DNMTi | DNA methyltransferase inhibitors |
References
- Peng, L.; Zhong, X. Epigenetic regulation of drug metabolism and transport. Acta Pharm. Sin. B 2015, 5, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Ingelman-Sundberg, M.; Zhong, X.B.; Hankinson, O.; Beedanagari, S.; Yu, A.M.; Peng, L.; Osawa, Y. Potential role of epigenetic mechanisms in the regulation of drug metabolism and transport. Drug. Metab. Dispos. 2013, 41, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Choi, Y.J.; Kim, J.W.; Chun, H.S.; Im, I.; Yoon, S.; Han, Y.M.; Song, C.W.; Kim, H. Differences in the epigenetic regulation of cytochrome p450 genes between human embryonic stem cell-derived hepatocytes and primary hepatocytes. PLoS ONE 2015, 10, e0132992. [Google Scholar] [CrossRef]
- Stadler, S.C.; Allis, C.D. Linking epithelial-to-mesenchymal-transition and epigenetic modifications. Semin. Cancer Biol. 2012, 22, 404–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Sciacovelli, M.; Frezza, C. Metabolic reprogramming and epithelial-to-mesenchymal transition in cancer. FEBS J. 2017, 284, 3132–3144. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Gomez, S.J.; Maziveyi, M.; Alahari, S.K. Regulation of epithelial-mesenchymal transition through epigenetic and post-translational modifications. Mol. Cancer 2016, 15, 18. [Google Scholar] [CrossRef]
- Cicchini, C.; Filippini, D.; Coen, S.; Marchetti, A.; Cavallari, C.; Laudadio, I.; Spagnoli, F.M.; Alonzi, T.; Tripodi, M. Snail controls differentiation of hepatocytes by repressing hnf4alpha expression. J. Cell Physiol. 2006, 209, 230–238. [Google Scholar] [CrossRef]
- Snykers, S.; Henkens, T.; De Rop, E.; Vinken, M.; Fraczek, J.; De Kock, J.; De Prins, E.; Geerts, A.; Rogiers, V.; Vanhaecke, T. Role of epigenetics in liver-specific gene transcription, hepatocyte differentiation and stem cell reprogrammation. J. Hepatol. 2009, 51, 187–211. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, Y.; Shimada, M.; Harimoto, N.; Rikimaru, T.; Shirabe, K.; Tanaka, S.; Sugimachi, K. Histone deacetylase inhibitor trichostatin a induces cell-cycle arrest/apoptosis and hepatocyte differentiation in human hepatoma cells. Int. J. Cancer 2003, 103, 572–576. [Google Scholar] [CrossRef]
- Sajadian, S.O.; Tripura, C.; Samani, F.S.; Ruoss, M.; Dooley, S.; Baharvand, H.; Nussler, A.K. Vitamin c enhances epigenetic modifications induced by 5-azacytidine and cell cycle arrest in the hepatocellular carcinoma cell lines hle and huh7. Clin. Epigenetics. 2016, 8, 46. [Google Scholar] [CrossRef]
- Michalopoulos, G.; Pitot, H.C. Primary culture of parenchymal liver cells on collagen membranes. Morphological and biochemical observations. Exp. Cell Res. 1975, 94, 70–78. [Google Scholar] [CrossRef]
- Kinoshita, T.; Miyajima, A. Cytokine regulation of liver development. Biochim. Biophys. Acta 2002, 1592, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Schyschka, L.; Muhl-Benninghaus, R.; Neumann, J.; Hao, L.; Nussler, N.; Dooley, S.; Liu, L.; Stockle, U.; Nussler, A.K.; et al. Comparative analysis of phase i and ii enzyme activities in 5 hepatic cell lines identifies huh-7 and hcc-t cells with the highest potential to study drug metabolism. Arch. Toxicol. 2012, 86, 87–95. [Google Scholar] [CrossRef]
- Nussler, A.K.; Vergani, G.; Gollin, S.M.; Dorko, K.; Morris, S.M., Jr.; Demetris, A.J.; Nomoto, M.; Beger, H.G.; Strom, S.C. Isolation and characterization of a human hepatic epithelial-like cell line (akn-1) from a normal liver. In. Vitro Cell Dev. Biol. Anim. 1999, 35, 190–197. [Google Scholar] [CrossRef]
- Ruoß, M.; Häussling, V.; Schügner, F.; Olde Damink, L.; Lee, S.; Ge, L.; Ehnert, S.; Nussler, A. A standardized collagen-based scaffold improves human hepatocyte shipment and allows metabolic studies over 10 days. Bioengineering 2018, 5, 86. [Google Scholar] [CrossRef]
- Rodriguez-Antona, C.; Donato, M.T.; Boobis, A.; Edwards, R.J.; Watts, P.S.; Castell, J.V.; Gomez-Lechon, M.J. Cytochrome p450 expression in human hepatocytes and hepatoma cell lines: Molecular mechanisms that determine lower expression in cultured cells. Xenobiotica 2002, 32, 505–520. [Google Scholar] [CrossRef]
- Seeliger, C.; Culmes, M.; Schyschka, L.; Yan, X.; Damm, G.; Wang, Z.; Kleeff, J.; Thasler, W.E.; Hengstler, J.; Stockle, U.; et al. Decrease of global methylation improves significantly hepatic differentiation of ad-mscs: Possible future application for urea detoxification. Cell Transplant. 2013, 22, 119–131. [Google Scholar] [CrossRef]
- Quint, K.; Agaimy, A.; Di Fazio, P.; Montalbano, R.; Steindorf, C.; Jung, R.; Hellerbrand, C.; Hartmann, A.; Sitter, H.; Neureiter, D.; et al. Clinical significance of histone deacetylases 1, 2, 3, and 7: Hdac2 is an independent predictor of survival in hcc. Virchows Arch. 2011, 459, 129–139. [Google Scholar] [CrossRef]
- Gou, Q.; He, S.; Zhou, Z. Protein arginine n-methyltransferase 1 promotes the proliferation and metastasis of hepatocellular carcinoma cells. Tumour Biol. 2017, 39, 1010428317691419. [Google Scholar] [CrossRef]
- Filipp, F.V. Crosstalk between epigenetics and metabolism-yin and yang of histone demethylases and methyltransferases in cancer. Brief Funct. Genomics 2017, 16, 320–325. [Google Scholar] [CrossRef]
- Dong, C.; Wu, Y.; Yao, J.; Wang, Y.; Yu, Y.; Rychahou, P.G.; Evers, B.M.; Zhou, B.P. G9a interacts with snail and is critical for snail-mediated e-cadherin repression in human breast cancer. J. Clin. Invest. 2012, 122, 1469–1486. [Google Scholar] [CrossRef]
- Agger, K.; Cloos, P.A.; Rudkjaer, L.; Williams, K.; Andersen, G.; Christensen, J.; Helin, K. The h3k27me3 demethylase jmjd3 contributes to the activation of the ink4a-arf locus in response to oncogene- and stress-induced senescence. Genes Dev. 2009, 23, 1171–1176. [Google Scholar] [CrossRef]
- Zhang, P.P.; Wang, X.L.; Zhao, W.; Qi, B.; Yang, Q.; Wan, H.Y.; Shuang, Z.Y.; Liu, M.; Li, X.; Li, S.; et al. DNA methylation-mediated repression of mir-941 enhances lysine (k)-specific demethylase 6b expression in hepatoma cells. J. Biol. Chem. 2014, 289, 24724–24735. [Google Scholar] [CrossRef]
- Kochat, V.; Equbal, Z.; Baligar, P.; Kumar, V.; Srivastava, M.; Mukhopadhyay, A. Jmjd3 aids in reprogramming of bone marrow progenitor cells to hepatic phenotype through epigenetic activation of hepatic transcription factors. PLoS ONE 2017, 12, e0173977. [Google Scholar] [CrossRef]
- Wu, J.; Du, C.; Lv, Z.; Ding, C.; Cheng, J.; Xie, H.; Zhou, L.; Zheng, S. The up-regulation of histone deacetylase 8 promotes proliferation and inhibits apoptosis in hepatocellular carcinoma. Dig. Dis. Sci. 2013, 58, 3545–3553. [Google Scholar] [CrossRef]
- Zhong, X.Y.; Yuan, X.M.; Xu, Y.Y.; Yin, M.; Yan, W.W.; Zou, S.W.; Wei, L.M.; Lu, H.J.; Wang, Y.P.; Lei, Q.Y. Carm1 methylates gapdh to regulate glucose metabolism and is suppressed in liver cancer. Cell Rep. 2018, 24, 3207–3223. [Google Scholar] [CrossRef]
- Sugimachi, K.; Tanaka, S.; Kameyama, T.; Taguchi, K.; Aishima, S.; Shimada, M.; Sugimachi, K.; Tsuneyoshi, M. Transcriptional repressor snail and progression of human hepatocellular carcinoma. Clin. Cancer Res. 2003, 9, 2657–2664. [Google Scholar]
- Zhaeentan, S.; Amjadi, F.S.; Zandie, Z.; Joghataei, M.T.; Bakhtiyari, M.; Aflatoonian, R. The effects of hydrocortisone on tight junction genes in an in vitro model of the human fallopian epithelial cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 229, 127–131. [Google Scholar] [CrossRef]
- Zhang, Q.; Lei, X.; Lu, H. Alterations of epigenetic signatures in hepatocyte nuclear factor 4alpha deficient mouse liver determined by improved chip-qpcr and (h)medip-qpcr assays. PLoS ONE 2014, 9, e84925. [Google Scholar]
- Weng, Y.R.; Cui, Y.; Fang, J.Y. Biological functions of cytokeratin 18 in cancer. Mol. Cancer Res. 2012, 10, 485–493. [Google Scholar] [CrossRef]
- Dannenberg, L.O.; Edenberg, H.J. Epigenetics of gene expression in human hepatoma cells: Expression profiling the response to inhibition of DNA methylation and histone deacetylation. BMC Genomics 2006, 7, 181. [Google Scholar] [CrossRef]
- Weng, M.K.; Natarajan, K.; Scholz, D.; Ivanova, V.N.; Sachinidis, A.; Hengstler, J.G.; Waldmann, T.; Leist, M. Lineage-specific regulation of epigenetic modifier genes in human liver and brain. PLoS ONE 2014, 9, e102035. [Google Scholar] [CrossRef]
- Oka, M.; Meacham, A.M.; Hamazaki, T.; Rodic, N.; Chang, L.J.; Terada, N. De novo DNA methyltransferases dnmt3a and dnmt3b primarily mediate the cytotoxic effect of 5-aza-2′-deoxycytidine. Oncogene 2005, 24, 3091–3099. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Kegel, V.; Zeilinger, K.; Hengstler, J.G.; Nussler, A.K.; Seehofer, D.; Damm, G. Featured article: Isolation, characterization, and cultivation of human hepatocytes and non-parenchymal liver cells. Exp. Biol. Med. 2015, 240, 645–656. [Google Scholar] [CrossRef]
- Knobeloch, D.; Ehnert, S.; Schyschka, L.; Buchler, P.; Schoenberg, M.; Kleeff, J.; Thasler, W.E.; Nussler, N.C.; Godoy, P.; Hengstler, J.; et al. Human hepatocytes: Isolation, culture, and quality procedures. Methods Mol. Biol. 2012, 806, 99–120. [Google Scholar]
- Sajadian, S.O.; Ehnert, S.; Vakilian, H.; Koutsouraki, E.; Damm, G.; Seehofer, D.; Thasler, W.; Dooley, S.; Baharvand, H.; Sipos, B.; et al. Induction of active demethylation and 5hmc formation by 5-azacytidine is tet2 dependent and suggests new treatment strategies against hepatocellular carcinoma. Clin. Epigenetics. 2015, 7, 98. [Google Scholar] [CrossRef]
- Lopez-Terrada, D.; Cheung, S.W.; Finegold, M.J.; Knowles, B.B. Hep g2 is a hepatoblastoma-derived cell line. Hum. Pathol. 2009, 40, 1512–1515. [Google Scholar] [CrossRef]
- Nakabayashi, H.; Taketa, K.; Miyano, K.; Yamane, T.; Sato, J. Growth of human hepatoma cells lines with differentiated functions in chemically defined medium. Cancer Res. 1982, 42, 3858–3863. [Google Scholar]
- Doi, I.; Namba, M.; Sato, J. Establishment and some biological characteristics of human hepatoma cell lines. Gann 1975, 66, 385–392. [Google Scholar]
- Saha, S.; Bardelli, A.; Buckhaults, P.; Velculescu, V.E.; Rago, C.; St Croix, B.; Romans, K.E.; Choti, M.A.; Lengauer, C.; Kinzler, K.W.; et al. A phosphatase associated with metastasis of colorectal cancer. Science 2001, 294, 1343–1346. [Google Scholar] [CrossRef]
- Hoffmann, S.A.; Muller-Vieira, U.; Biemel, K.; Knobeloch, D.; Heydel, S.; Lubberstedt, M.; Nussler, A.K.; Andersson, T.B.; Gerlach, J.C.; Zeilinger, K. Analysis of drug metabolism activities in a miniaturized liver cell bioreactor for use in pharmacological studies. Biotechnol. Bioeng. 2012, 109, 3172–3181. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Origin/Disease | Donor | Reference |
|---|---|---|---|
| HepG2 | hepatoblastoma | 15 year old Caucasian male | [39] |
| Huh7 | HCC | 57 year old Japanese male | [40] |
| HLE | HCC | 68-year-old patient | [41] |
| AKN1 | Healthy | 10 year old male | [16] |
| Supplement | Concentration | Day | Unstimulated | Condition 1 | Condition 2 |
|---|---|---|---|---|---|
| FCS | 10% | 0–3 | + | + | + |
| P/S | 1% | 0–3 | + | + | + |
| 5-Azacytidin | 10 µM | 1–3 | - | + | + |
| Vitamin C | 0.5 mM | 1–3 | - | + | + |
| Human Insulin | 1 mM | 2–3 | - | - | + |
| Hydrocortisone | 0.8 µg/mL | 2–3 | - | - | + |
| Gen | Forward/Reverse Sequences | Annealing Tm | Product Length (bp) | GenBank Accession |
|---|---|---|---|---|
| hHNF4A | CAGGCTCAAGAAATGCTTCC GGCTGCTGTCCTCATAGCTT | 59 | 101 | NM_001287184.1 |
| hCK18 | GAGTATGAGGCCCTGCTGAACAT GCGGGTGGTGGTCTTTTGGAT | 65 | 150 | NM_199187.1 |
| hHDAC1 | AACTGCTAAAGTATCACCAGAGGGT CCGGTCCGTGGTGTAGAAGG | 62 | 92 | NM_004964.2 |
| hHDAC2 | TGAAGGAGAAGGAGGTCGAA GGATTTATCTTCTTCCTTAACGTCTG | 59 | 124 | NM_001527.3 |
| hCYP1A2 | GCTTCGGACAGCACTTCCCT AGAAGTCCAGGGGGTTCCCG | 63 | 105 | NM_000761.4 |
| hCYP3A4 | AGCCCAGCAAAGAGCAACAC TCCATATAGATAGAGGAGCACCAGG | 60 | 147 | NM_017460.5 |
| hCYP2C9 | GACATGAACAACCCTCAGGACTTT TGCTTGTCGTCTCTGTCCCA | 62 | 145 | NM_000771.3 |
| hKDM4C | TGGATCCCAGATAGCAATGA TGTCTTCAAATCGCATGTCA | 59 | 110 | NM_001304340.1 |
| hKDM6B | GGAGGCCACACGCTGCTAC GCCAGTATGAAAGTTCCAGAGCTG | 63 | 112 | NM_001348716.1 |
| hSNAIL | ACCACTATGCCGCGCTCTT GGTCGTAGGGCTGCTGGAA | 60 | 115 | NM_005985.3 |
| hGAPDH | TGCACCACCAACTGCTTAGC GGCATGGACTGTGGTCATGAG | 59 | 87 | NM_002046.3 |
| Substrate | Isoenzyme | Incubation Time in Hours | Concentration | Reaction |
|---|---|---|---|---|
| Bupropion | CYP2B6 | 1 | 100 μM | Bupropion-hydroxylation |
| Diclofenac | CYP2C9 | 1 | 9 μM | Diclofenac-4’-hydroxylation |
| Testosterone | CYP3A4 | 1 | 50 μM | Testosterone-6β-hydroxylation |
| Bufuralol | CYP2D6 | 2 | 9 μM | Bufuralol-1-hydroxylation |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruoß, M.; Damm, G.; Vosough, M.; Ehret, L.; Grom-Baumgarten, C.; Petkov, M.; Naddalin, S.; Ladurner, R.; Seehofer, D.; Nussler, A.; et al. Epigenetic Modifications of the Liver Tumor Cell Line HepG2 Increase Their Drug Metabolic Capacity. Int. J. Mol. Sci. 2019, 20, 347. https://doi.org/10.3390/ijms20020347
Ruoß M, Damm G, Vosough M, Ehret L, Grom-Baumgarten C, Petkov M, Naddalin S, Ladurner R, Seehofer D, Nussler A, et al. Epigenetic Modifications of the Liver Tumor Cell Line HepG2 Increase Their Drug Metabolic Capacity. International Journal of Molecular Sciences. 2019; 20(2):347. https://doi.org/10.3390/ijms20020347
Chicago/Turabian StyleRuoß, Marc, Georg Damm, Massoud Vosough, Lisa Ehret, Carl Grom-Baumgarten, Martin Petkov, Silvio Naddalin, Ruth Ladurner, Daniel Seehofer, Andreas Nussler, and et al. 2019. "Epigenetic Modifications of the Liver Tumor Cell Line HepG2 Increase Their Drug Metabolic Capacity" International Journal of Molecular Sciences 20, no. 2: 347. https://doi.org/10.3390/ijms20020347
APA StyleRuoß, M., Damm, G., Vosough, M., Ehret, L., Grom-Baumgarten, C., Petkov, M., Naddalin, S., Ladurner, R., Seehofer, D., Nussler, A., & Sajadian, S. (2019). Epigenetic Modifications of the Liver Tumor Cell Line HepG2 Increase Their Drug Metabolic Capacity. International Journal of Molecular Sciences, 20(2), 347. https://doi.org/10.3390/ijms20020347