Structure and Mechanism of the Divalent Anion/Na+ Symporter

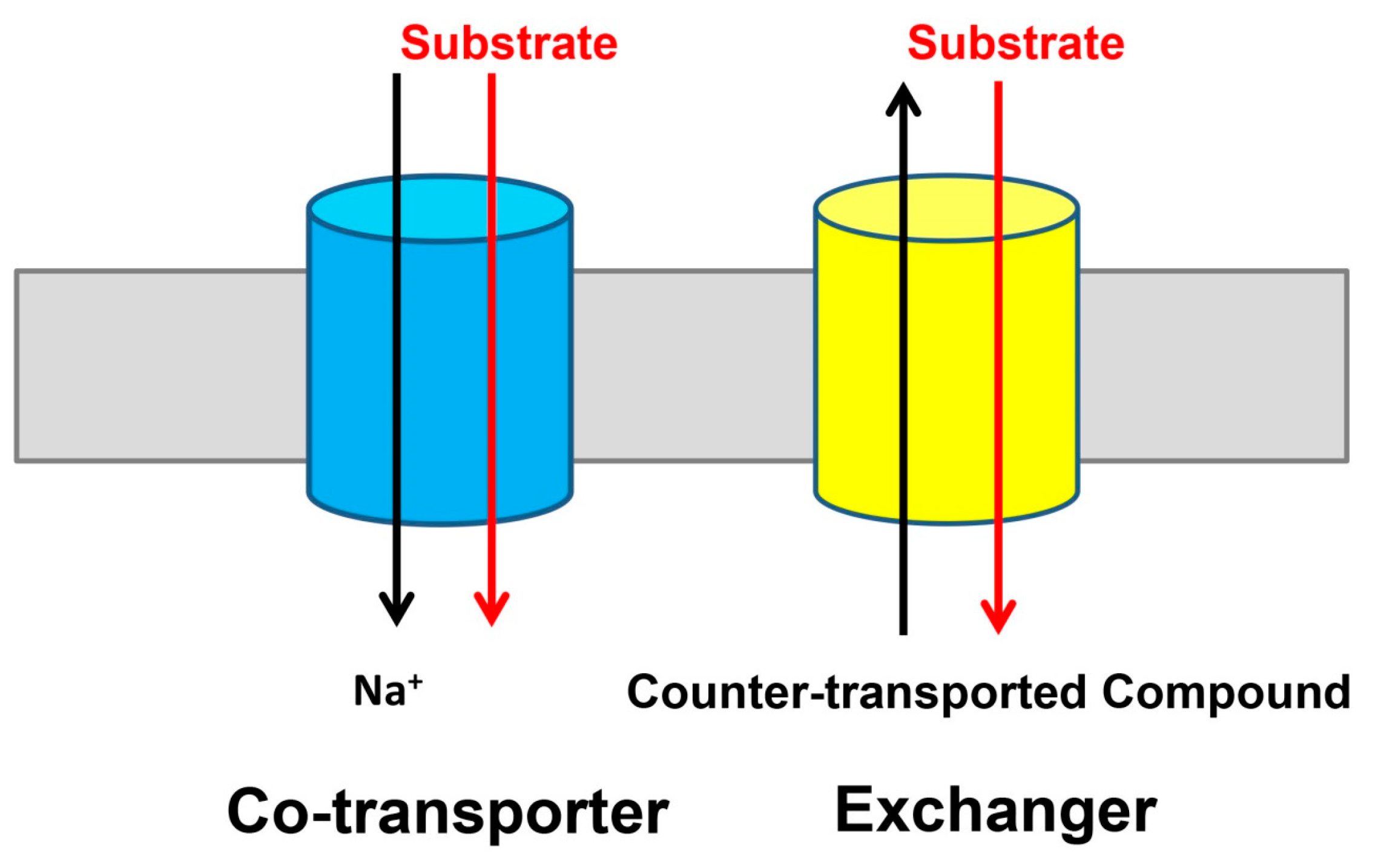{kind=link}
{kind=link}
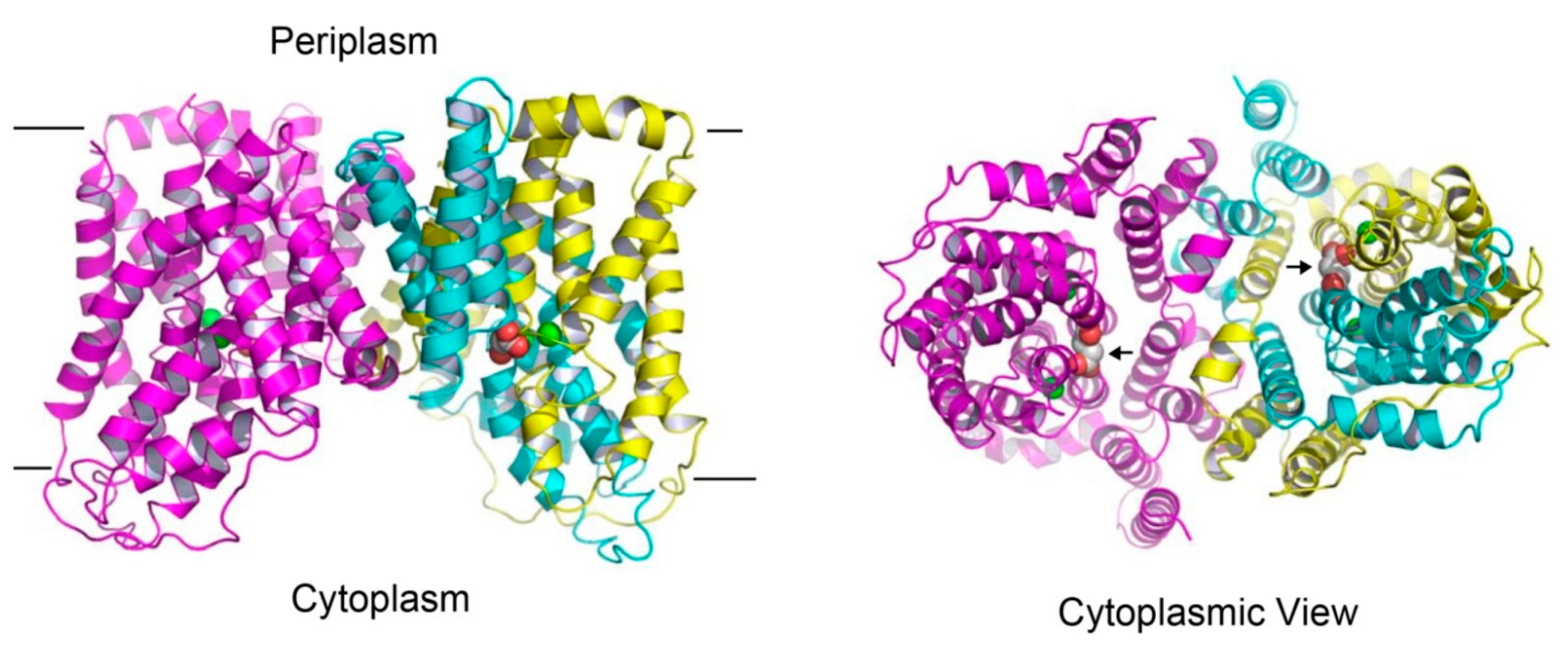{kind=link}
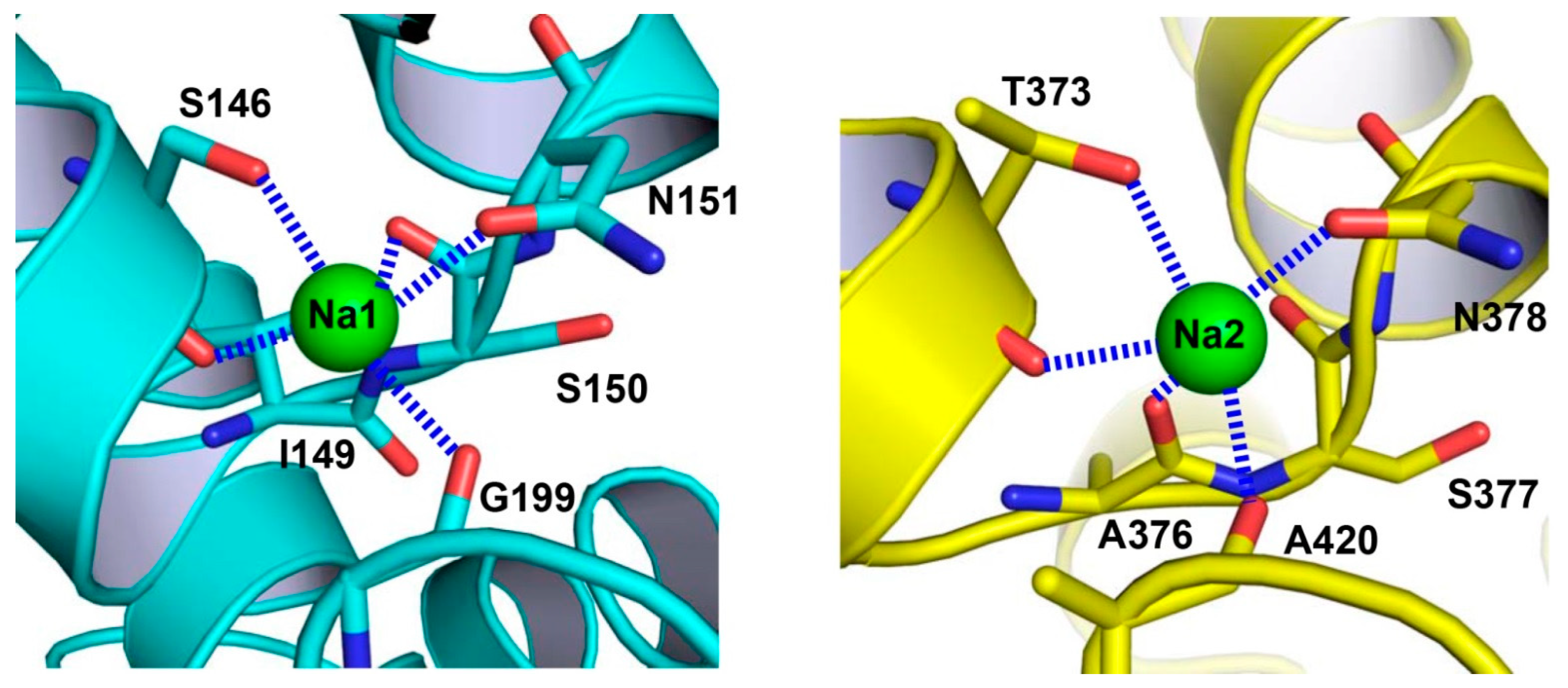{kind=link}
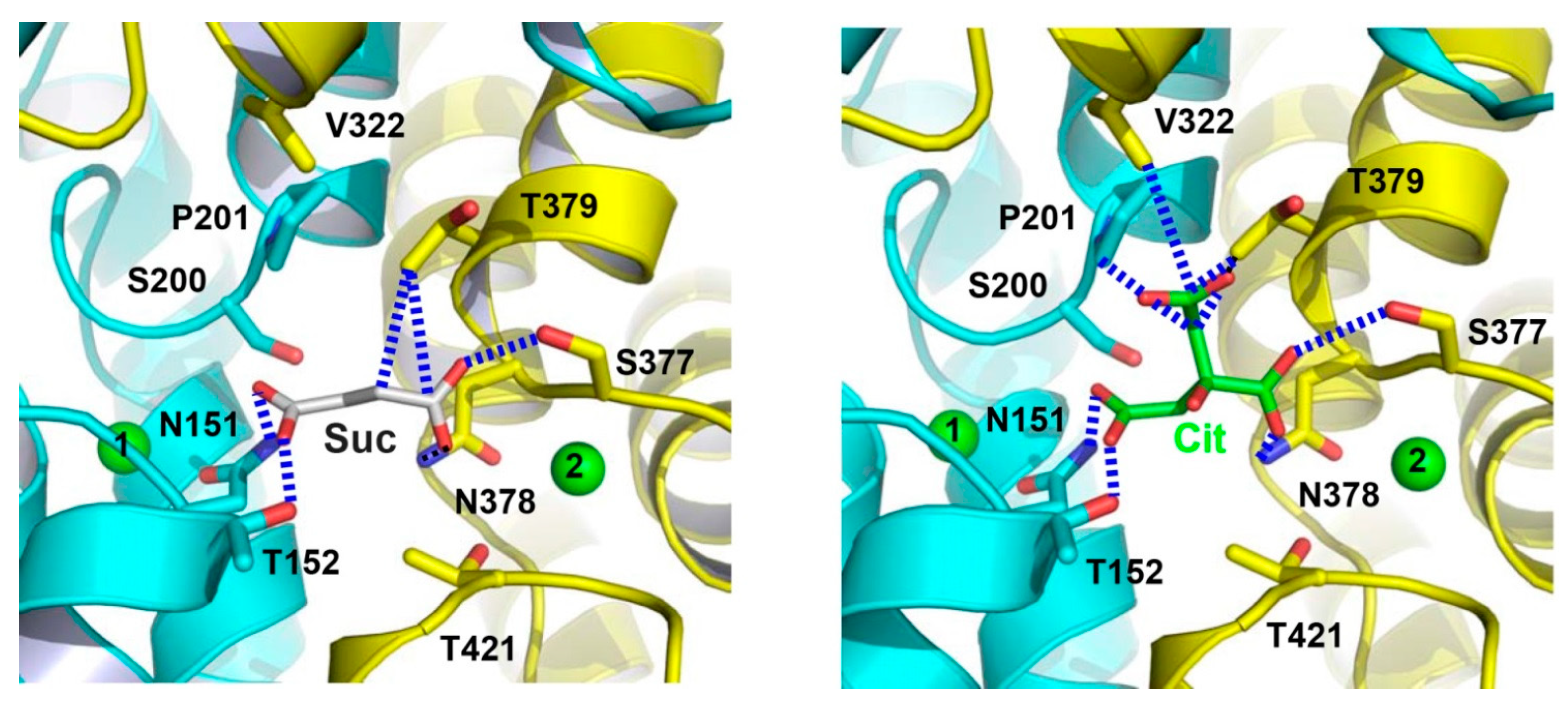{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Structure Determination of DASS
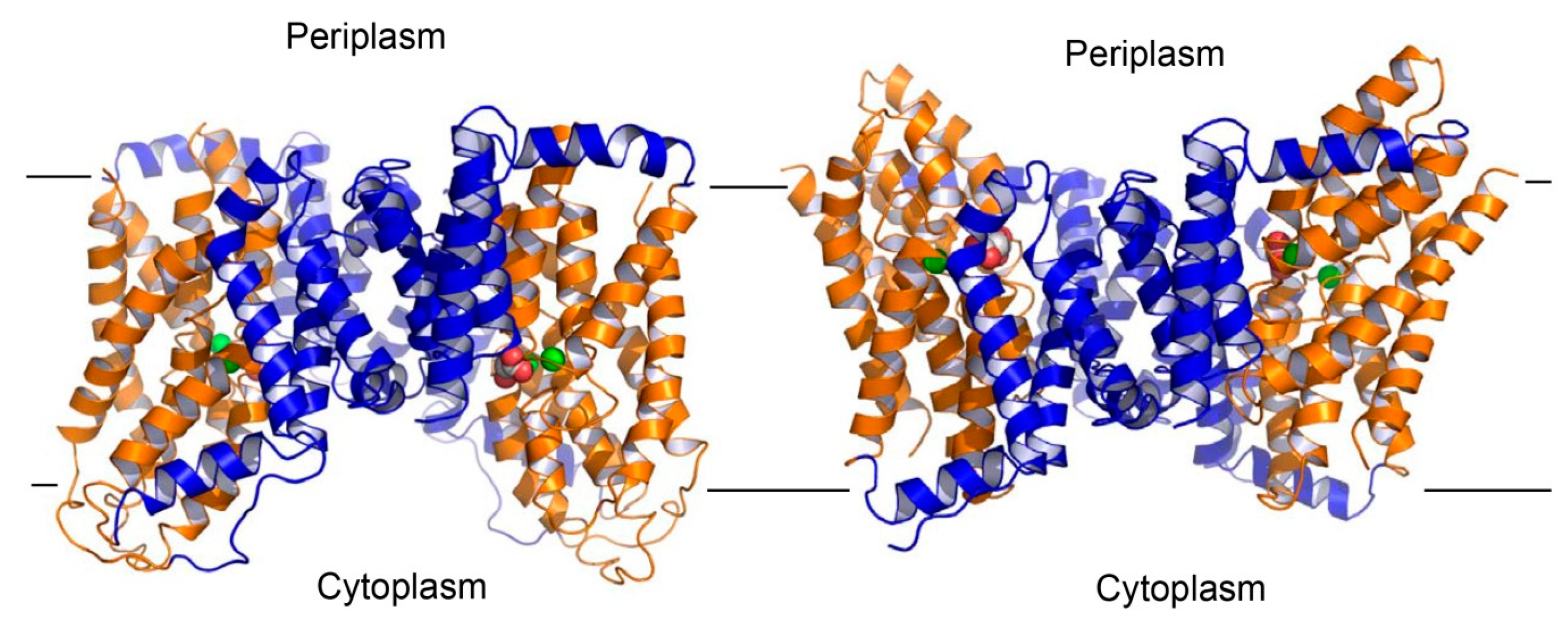
3. Overall Structure of VcINDY
4. Na+-binding Sites in VcINDY
5. Di- and Tri-carboxylate Binding Sites in VcINDY
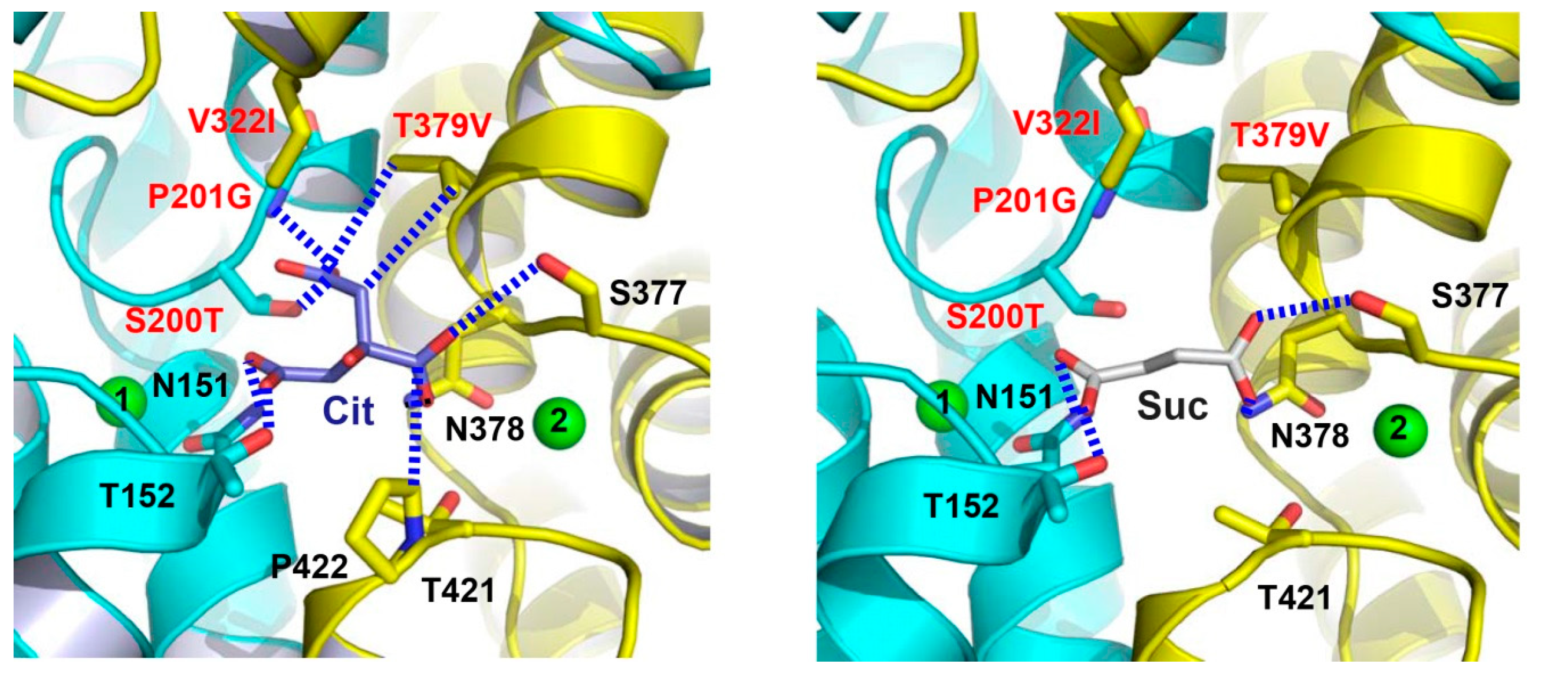
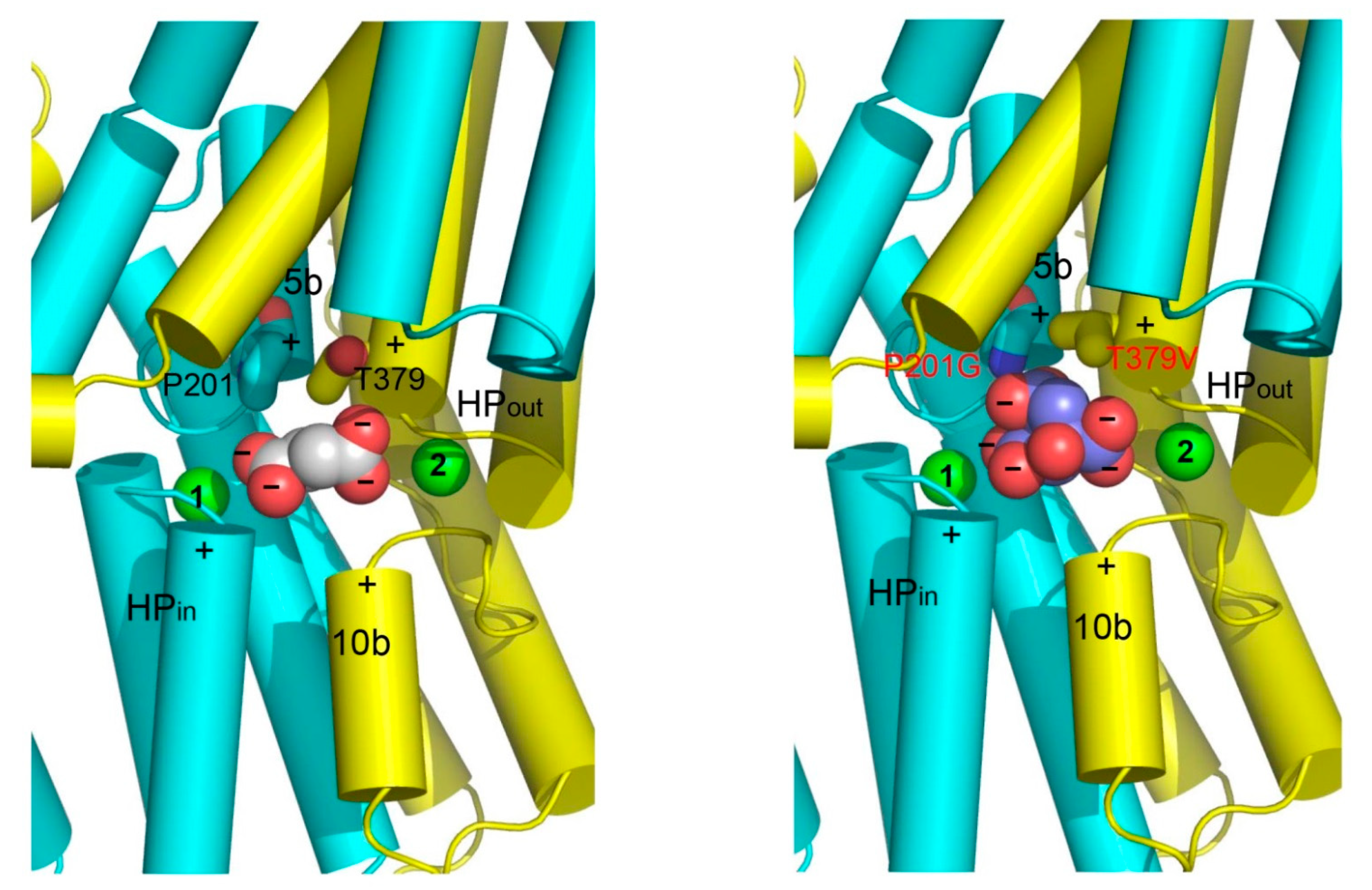
6. Structures of A Humanized Variant of VcINDY
7. Substrate Recognition By DASS and Other anion Transporters
8. Elevator-like Mechanism and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DASS | Divalent Anion/Sodium Symporter |
| NaDC1 | Na+-dependent DiCarboxylate symporter1 |
| NaDC3 | Na+-dependent DiCarboxylate symporter3 |
| NaCT | Na+-dependent Citrate Transporter |
| INDY | I’m Not Dead Yet |
| NaS1 | Na+-dependent Sulphate symporter1 |
| NaS2 | Na+-dependent Sulphate symporter2 |
| VcINDY | Vibrio Cholerae I’m Not Dead Yet (a divalent anion/sodium symporter from Vibrio cholerae) |
References
- Pajor, A.M. Sodium-coupled dicarboxylate and citrate transporters from the SLC13 family. Pflugers Arch. 2014, 466, 119–130. [Google Scholar] [CrossRef]
- Markovich, D. Na+-sulfate cotransporter SLC13A1. Pflugers Arch. 2014, 466, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Copper, G.; Singhi, S.; Saier, M.H. The ion transporter superfamily. Biochim. Biophys. Acta 2003, 1618, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S.H.; Kippen, I.; Klinenberg, J.R.; Wright, E.M. Specificity of the transport system for tricarboxylic acid cycle intermediates in renal brush borders. J. Membrane Biol. 1980, 57, 73–82. [Google Scholar] [CrossRef]
- Burckhardt, B.C.; Drinkuth, B.; Menzel, C.; Konig, A.; Steffgen, J.; Wright, S.H.; Burckhardt, G. The renal Na+-dependent dicarboxylate transporter, NADC-3, translocates dimethyl- and disulfhydryl-compounds and contributes to renal heavy metal detoxification. J. Am. Soc. Nephrol. 2002, 13, 2628–2638. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Fei, Y.J.; Zhuang, L.; Gopal, E.; Miyauchi, S.; Ganapathy, V. Functional features and genomic organization of mouse NaCT, a sodium-coupled transporter for tricarboxylic acid cycle intermediates. Biochem. J. 2004, 378, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.E.; Waldegger, S.; Herzer, T.; Biber, J.; Markovich, D.; Murer, H.; Lang, F. Electrogenic cotransport of Na+ and sulfate in Xenopus oocytes expressing the cloned Na+SO4(2−) transporter protein NaSi-1. J. Biol. Chem. 1994, 269, 12407–12409. [Google Scholar]
- Markovich, D.; Regeer, R.R.; Kunzelmann, K.; Dawson, P.A. Functional characterization and genomic organization of the human Na+-sulfate cotransporter hNaS2 gene (SLC13A4). Biochem. Biophys. Res. Commun. 2005, 326, 729–734. [Google Scholar] [CrossRef]
- Bergeron, M.J.; Clemencon, B.; Hediger, M.A.; Markovich, D. SLC family of Na+-coupled di- an dtri-carboxylate/sulfate transporter. Mol. Asp. Med. 2013, 34, 299–312. [Google Scholar] [CrossRef]
- Hall, J.A.; Pajor, A.M. Functional characterization of Na+-coupled dicarboxylate carrier protein from Staphylococcus aureus. J. Bacteriol. 2005, 187, 5189–5194. [Google Scholar] [CrossRef]
- Hall, J.A.; Pajor, A.M. Functional reconstitution of SdcS, a Na+-coupled dicarboxylate carrier protein from Staphylococcus aureus. J. Bacteriol. 2007, 189, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.W.; Jolkver, E.; Kramer, R.; Marin, K.; Wendisch, V.F. Identification and characterization of the dicarboxylate uptake system DccT in Corynebacterium glutamicum. J. Bacteriol. 2008, 190, 6458–6466. [Google Scholar] [CrossRef] [PubMed]
- Strickler, M.A.; Hall, J.A.; Gaiko, O.; Pajor, A.M. Functional characterization of a Na+-coupled dicarboxylate transporter from Bacillus licheniformis. Biochim. Biophys. Acta 2009, 1788, 2489–2496. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.; Fitzgerald, G.A.; Wang, D.N.; Mindell, J.A. Functional characterization of a Na+-dependent dicarboxylate transporter from Vibrio cholera. J. Gen. Physiol. 2014, 143, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Knauf, F.; Rogina, B.; Jiang, Z.; Aronson, P.S.; Helfand, S.L. Functional characterization and immunolocalization of the transporter encoded by the life-extending gene Indy. Proc. Natl. Acad. Sci. USA 2002, 99, 14315–14319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Fei, Y.J.; Huang, W.; Zhuang, L.; Chen, Z.; Ganapathy, V. Functional identity of Drosophila melanogaster Indy as a cation-independent, electroneutral transporter for tricarboxylic acid-cycle intermediates. Biochem. J. 2002, 367, 313–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogina, B.; Reenan, R.A.; Nilsen, S.P.; Helfand, S.L. Extended life-span conferred by cotransporter gene mutations in Drosophila. Science 2000, 290, 2137–2140. [Google Scholar] [CrossRef] [PubMed]
- Neretti, N.; Wang, P.Y.; Brodsky, A.S.; Nyguyen, H.H.; White, K.P.; Rogina, B.; Helfand, S.L. Long-lived Indy induces reduced mitochondrial reactive oxygen species production and oxidative damage. Proc. Natl. Acad. Sci. USA 2009, 106, 2277–2282. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.J.; Inoue, K.; Ganapathy, V. Structural and functional characteristics of two-sodium-coupled dicarboxylate transporters (ceNaDC1 and ceNaDC2) from Caenorhabditis elegans and their relevance to life span. J. Biol. Chem. 2003, 278, 6136–6144. [Google Scholar] [CrossRef]
- Fei, Y.J.; Liu, J.C.; Inoue, K.; Zhaung, L.; Miyake, K.; Miyauchi, S.; Ganapathy, V. Relevance of NAC-2, an Na+-coupled citrate transporter, to life span, body size and fat content in Caenorhabditis elegans. Biochem. J. 2004, 379, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Birkenfeld, A.L.; Lee, H.-Y.; Guebre-Egziabher, F.; Alves, T.C.; Jurczak, M.J.; Jornayvaz, F.R.; Zhang, D.; Hsiao, J.J.; Martin-Montalvo, A.; Fischer-Rosinsky, A.; et al. Deletion of the mammalian INDY homolog mimics aspects of dietary restriction and protects against adiposity and insulin resistance in mice. Cell Metab. 2011, 14, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Bhutia, Y.D.; Kopel, J.J.; Lawrence, J.J.; Neugebauer, V.; Ganapathy, V. Plasma Membrane Na⁺-Coupled Citrate Transporter (SLC13A5) and Neonatal Epileptic Encephalopathy. Molecules 2017, 22, 378. [Google Scholar] [CrossRef]
- Li, Z.; Li, D.; Choi, E.Y.; Lapidus, R.; Zhang, L.; Huang, S.M.; Shapiro, P.; Wang, H. Silencing of solute carrier family 13 member 5 disrupts energy homeostasis and inhibits proliferation of human hepatocarcinoma cells. J. Biol. Chem. 2017, 292, 13890–13901. [Google Scholar] [CrossRef] [Green Version]
- Mancusso, R.; Gregorio, G.G.; Liu, Q.; Wang, D.N. Structure and mechanism of a bacterial sodium-dependent dicarboxylate transporter. Nature 2012, 491, 622–626. [Google Scholar] [CrossRef] [Green Version]
- Nie, R.; Stark, S.; Symersky, J.; Kaplan, R.S.; Lu, M. Structure and function of the divalent anion/Na+ symporter from Vibrio cholerae and a humanized variant. Nat. Commun. 2017, 8, 15009. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Bolla, J.R.; Kumar, N.; Radhakrishnan, A.; Long, F.; Delmar, J.A.; Chou, T.-H.; Rajashankar, K.R.; Shafer, W.M.; Yu, E.W. Structure and function of Neisseria gonorrhoeae MtrF illuminates a class of antimetabolite efflux pumps. Cell Rep. 2015, 11, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Delmar, J.A.; Radhakrishnan, A.; Chou, T.-H.; Rajashankar, K.R.; Yu, E.W.; Bolla, J.R.; Kumar, N.; Long, F. Crystal structure of the Alcanivorax borkumensis YdaH transporter reveals an unusual topology. Nat. Commun. 2015, 6, 6874. [Google Scholar]
- Johnson, Z.L.; Cheong, C.G.; Lee, S.Y. Crystal structure of a concentrative nucleotide transporter from Vibrio cholerae at 2.4 Å. Nature 2012, 483, 489–493. [Google Scholar] [CrossRef]
- Boudker, O.; Ryan, R.M.; Yernool, D.; Shimamoto, K.; Gouaux, E. Coupling substrate and ion binding to extracellular gate of a sodium-dependent aspartate transporter. Nature 2007, 445, 387–393. [Google Scholar] [CrossRef]
- Yamashita, A.; Singh, S.K.; Kawate, T.; Jin, Y.; Gouaux, E. Crystal structure of a bacterial homologue of Na+/Cl−-dependent neurotransmitter transporters. Nature 2005, 437, 215–223. [Google Scholar] [CrossRef]
- Gouaux, J.E.; Lipscomb, W.N. Three-dimensional structure of carbamoyl phosphate and succinate bound to aspartate carbamoyltransferase. Proc. Natl. Acad. Sci. USA 1988, 85, 4205–4208. [Google Scholar] [CrossRef] [PubMed]
- Leys, D.; Tsapin, A.S.; Nealson, K.H.; Meyer, T.E.; Cusanovich, M.A.; Van Beeumen, J.J. Structure and mechanism of the flavocytochrome c fumarate reductase of Shewanella putrefaciens MR-1. Nat. Struct. Biol. 1999, 6, 1113–1117. [Google Scholar] [PubMed]
- Muller, I.; Stuckl, C.; Wakeley, J.; Kertesz, M.; Uson, I. Succinate complex crystal structures of the α-ketoglutarate-dependent dioxygenase AtsK. J. Biol. Chem. 2005, 280, 5716–5723. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-F.; Nan, B.; Nan, J.; Ma, Q.; Panjikar, S.; Liang, Y.-H.; Wang, Y.; Su, X.-D. C4-dicarboxylates sensing mechanism revealed by the crystal structures of DctB sensor domain. J. Mol. Biol. 2008, 383, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.; Hendrickson, W.A. Crystal structures of C4-dicarboxylate ligand complexes with sensor domains of histidine kinases DcuS and DctB. J. Biol. Chem. 2008, 283, 30256–30265. [Google Scholar] [CrossRef] [PubMed]
- Dutzler, R.; Campbell, E.B.; Cadene, M.; Chait, B.T.; MacKinnon, R. X-ray structure of a ClC chloride channel at 3.0 Å reveals the molecular basis of anion selectivity. Nature 2002, 415, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Hibbs, R.E.; Gouaux, E. Principles of activation and permeation in an anion-selective Cys-loop receptor. Nature 2011, 474, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Dickson, V.K.; Pedi, L.; Long, S.B. Structure and insights into the function of a Ca2+-activated Cl− channel. Nature 2014, 516, 213–218. [Google Scholar] [CrossRef]
- Geertsma, E.R.; Chang, Y.N.; Shaik, F.; Neldner, Y.; Pardon, E.; Steyaert, J.; Dutzler, R. Structure of a prokaryotic fumarate transporter reveals the architecture of the SLC26 family. Nat. Struct. Mol. Biol. 2015, 22, 803–808. [Google Scholar] [CrossRef]
- Wöhlert, D.; Grötzinger, M.J.; Kühlbrandt, W.; Yildiz, Ö. Mechanism of Na(+)-dependent citrate transport from the structure of an asymmetrical CitS dimer. eLife 2015, 4, e09375. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, S.; Kim, S.; Lee, H.; Lee, J.O.; Jin, M.S. Structural insights into the elevator-like mechanism of the sodium/citrate symporter CitS. Sci. Rep. 2017, 7, 2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guskov, A.; Jensen, S.; Faustino, I.; Marrink, S.J.; Slotboom, D.J. Coupled binding mechanism of three sodium ions and aspartate in the glutamate transporter homologue GltTk. Nat. Commun. 2016, 7, 13420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Wisedchaisri, G.; Gonen, T. Crystal structure of a nitrate/nitrite exchanger. Nature 2013, 497, 647–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Huang, W.; Yan, C.; Gong, X.; Jiang, S.; Zhao, Y.; Wang, J.; Shi, Y. Structure and mechanism of a nitrate transporter. Cell Rep. 2013, 3, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Takeda, H.; Kato, H.; Doki, S.; Ito, K.; Maturna, A.D.; Ishitani, R.; Nureki, O. Structural basis for dynamic mechanism of nitrate/nitrite antiport by NarK. Nat. Commun. 2015, 6, 7097. [Google Scholar] [CrossRef] [Green Version]
- Scopelliti, A.J.; Font, J.; Vandenberg, R.J.; Boudker, O.; Ryan, R.M. Structural characterisation reveals insights into substrate recognition by the glutamine transporter ASCT2/SLC1A5. Nat. Commun. 2018, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P. A general theory of membrane transport from studies of bacteria. Nature 1957, 180, 134–136. [Google Scholar] [CrossRef]
- Jardetzky, O. Simple allosteric model for membrane pumps. Nature 1966, 211, 969–970. [Google Scholar] [CrossRef]
- Mulligan, C.; Fenollar-Ferrer, C.; Fitzgerald, G.A.; Vergara-Jaque, A.; Kaufmann, D.; Li, Y.; Forrest, L.R.; Mindell, J.A. The bacterial dicarboxylate transporter VcINDY uses a two-domain elevator-type mechanism. Nat. Struct. Mol. Biol. 2016, 23, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Drew, D.; Boudker, O. Shared molecular mechanisms of membrane transporters. Annu. Rev. Biochem. 2016, 85, 543–572. [Google Scholar] [CrossRef]
- Colas, C.; Pajor, A.M.; Schlessinger, A. Structure-based identification of inhibitors for the SLC13 family pf Na+/dicarboxylate cotransporters. Biochemistry 2015, 54, 4900–4908. [Google Scholar] [CrossRef] [PubMed]
- Huard, K.; Gosset, J.R.; Montgomery, J.I.; Gilbert, A.; Hayward, M.M.; Magee, T.V.; Cabral, S.; Uccello, D.P.; Bahnck, K.; Brown, J.; et al. Optimization of a dicarboxylate series for in vivo inhibition of citrate transport by the solute carrier 13 (SLC13) family. J. Med. Chem. 2016, 59, 1165–1175. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, M. Structure and Mechanism of the Divalent Anion/Na+ Symporter. Int. J. Mol. Sci. 2019, 20, 440. https://doi.org/10.3390/ijms20020440
Lu M. Structure and Mechanism of the Divalent Anion/Na+ Symporter. International Journal of Molecular Sciences. 2019; 20(2):440. https://doi.org/10.3390/ijms20020440
Chicago/Turabian StyleLu, Min. 2019. "Structure and Mechanism of the Divalent Anion/Na+ Symporter" International Journal of Molecular Sciences 20, no. 2: 440. https://doi.org/10.3390/ijms20020440
APA StyleLu, M. (2019). Structure and Mechanism of the Divalent Anion/Na+ Symporter. International Journal of Molecular Sciences, 20(2), 440. https://doi.org/10.3390/ijms20020440