The Rice Alpha-Amylase, Conserved Regulator of Seed Maturation and Germination



Abstract
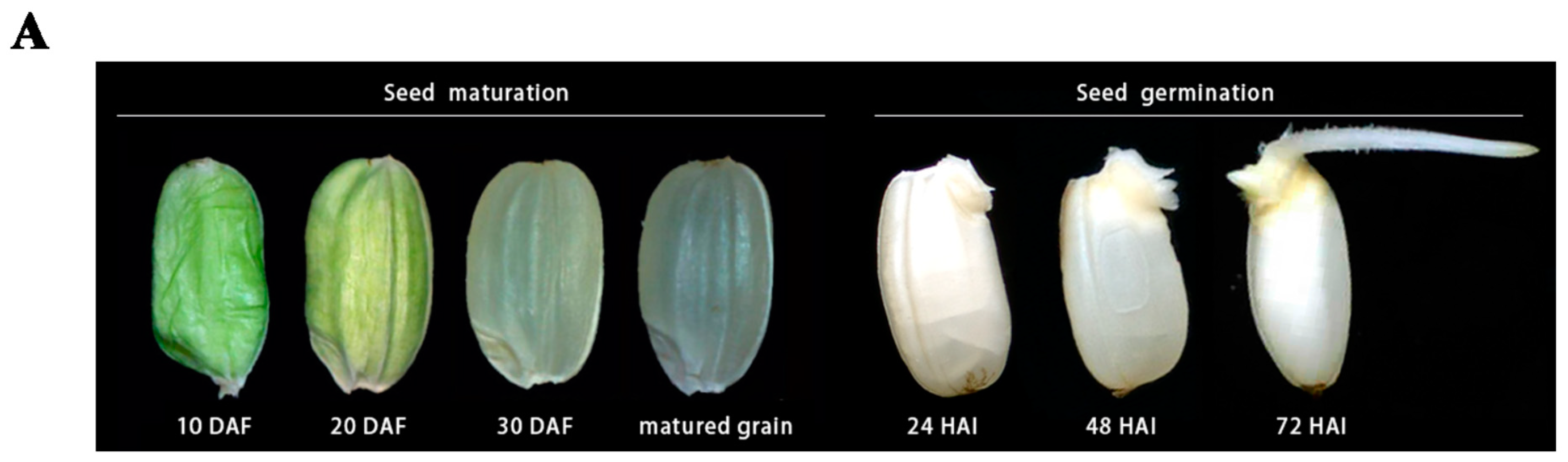
:1. Rice Seed Maturation and Germination
1.1. Rice Seed Maturation and Starch Storage
1.2. Seed Germination
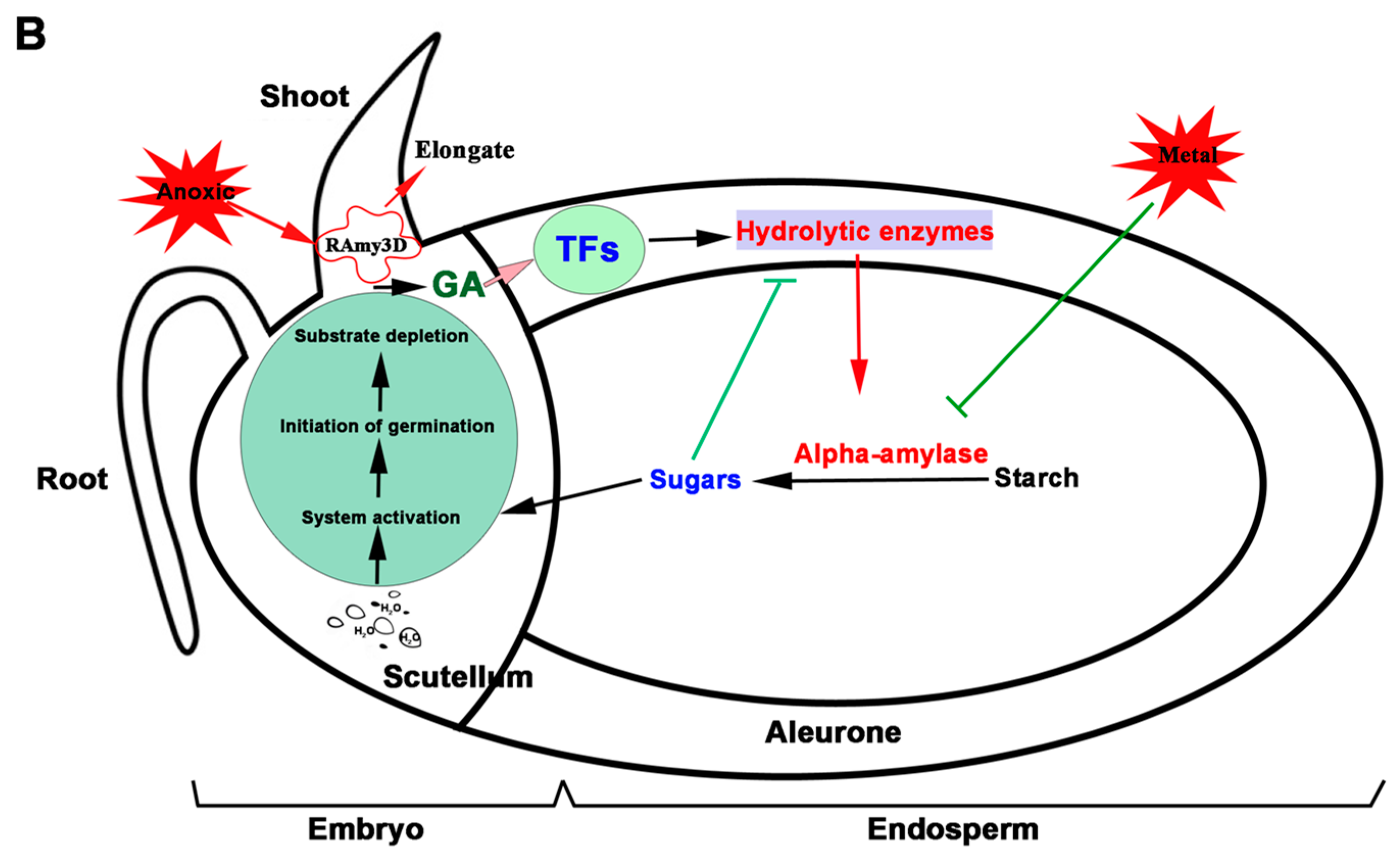
2. Functions of Different Rice Seed Organs in Germination
2.1. The Embryo
2.2. The Endosperm
2.3. The Scutellum and Aleurone Layer
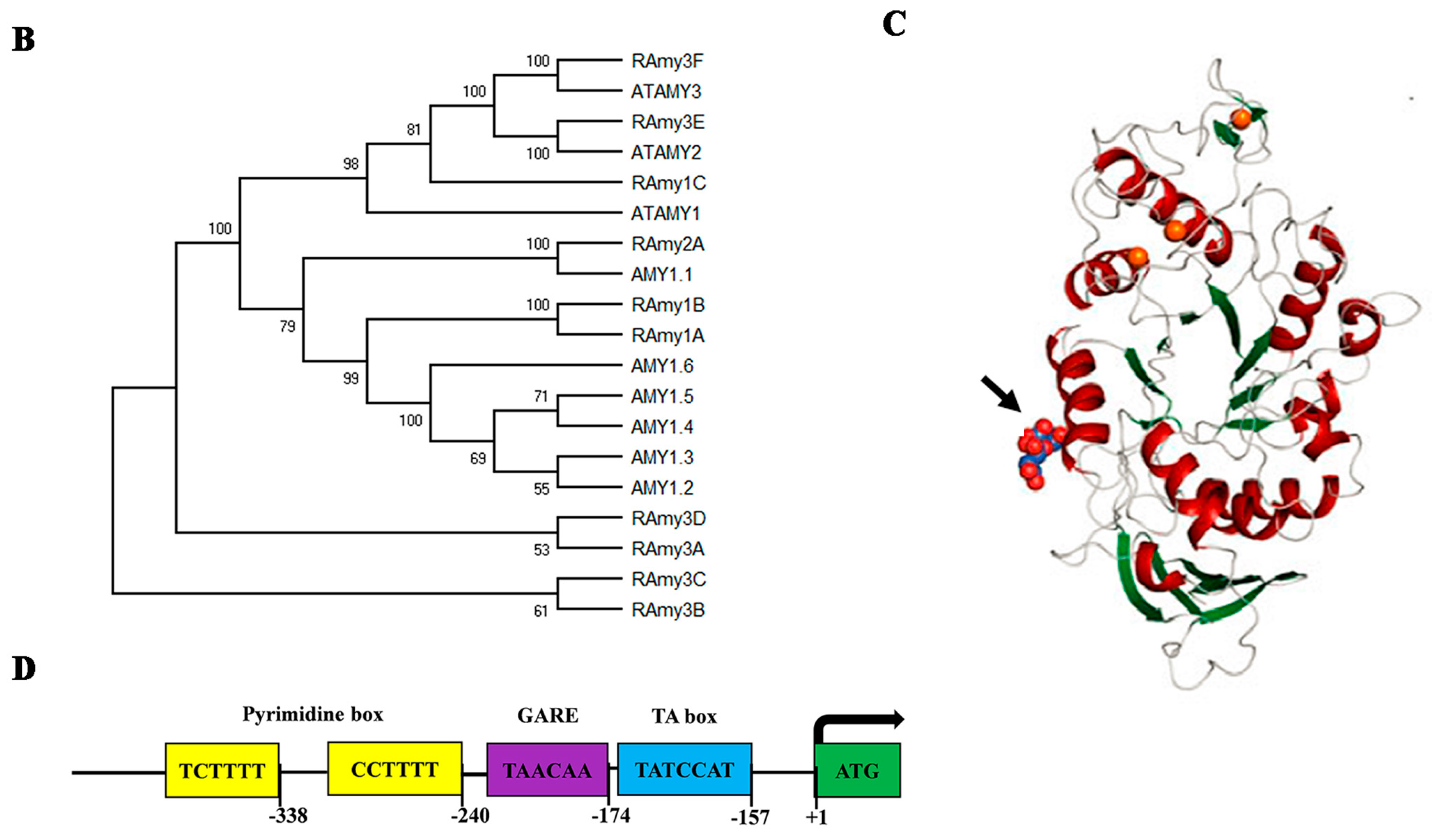
3. Classification of Alpha Amylase
4. Structure of the Rice Alpha-Amylase Gene
5. Expression Traits of Alpha-Amylase
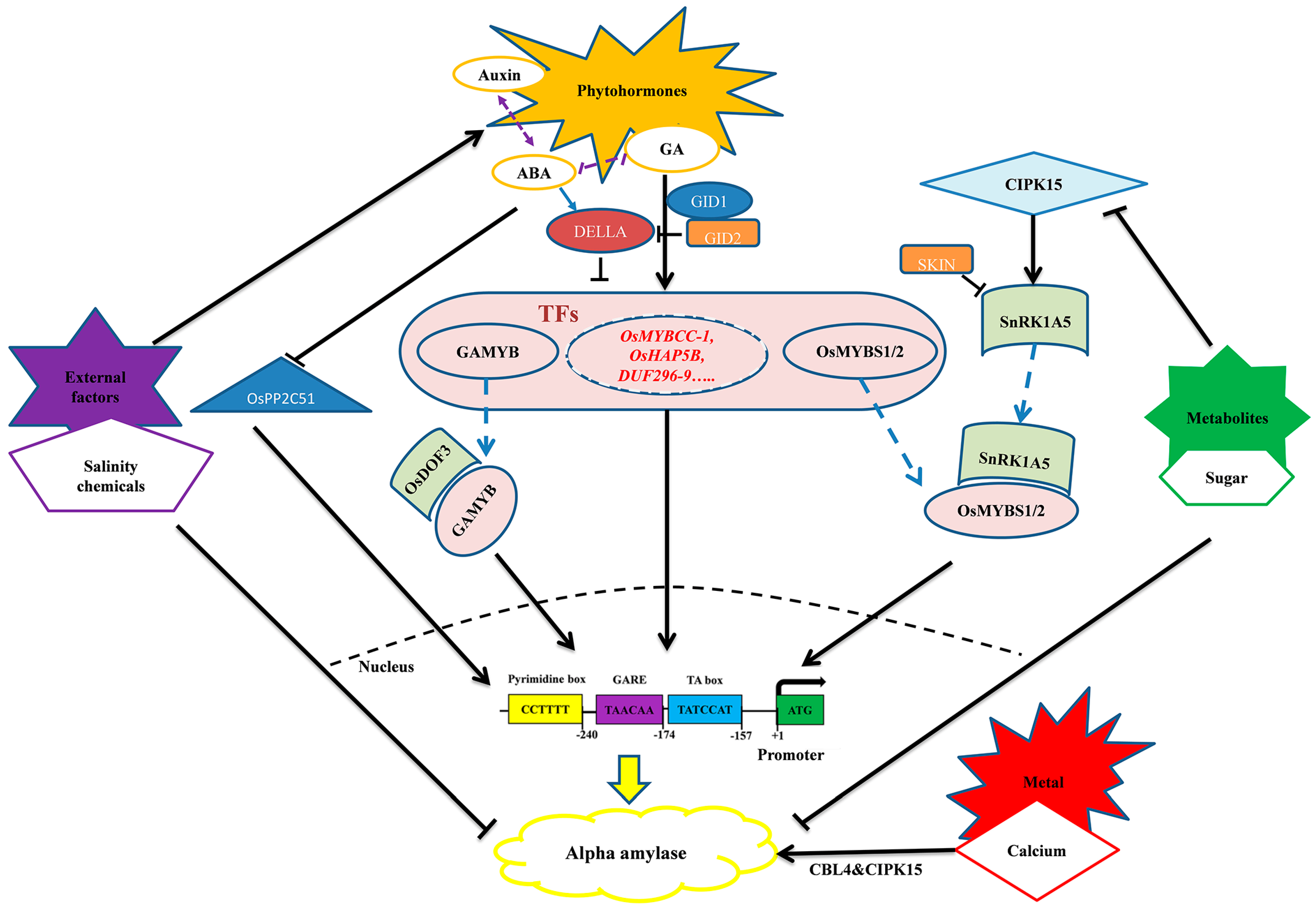
6. Regulation of Alpha-Amylase Expression
6.1. Hormonal Regulation of Alpha-Amylase
6.2. Metabolite Regulation of Alpha-Amylase
6.3. Other Factors Influencing the Expression of Alpha-Amylase
7. Challenges and Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Xue, L.-J.; Zhang, J.-J.; Xue, H.-W. Genome-Wide Analysis of the Complex Transcriptional Networks of Rice Developing Seeds. PLoS ONE 2012, 7, e31081. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, Y.; Michiyama, H.; Hirano, T. Involvement of α-Amylase Genes in Starch Degradation in Rice Leaf Sheaths at the Post-Heading Stage. Plant Prod. Sci. 2015, 18, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Cock, J.H.; Yoshida, S. Accumulation of 14C-labelled cabohydrate before flowering and its subsequent redistribution and respiration in the rice plant. Nippon Sakumotsu Gakkai Kiji 1972, 41, 226–234. [Google Scholar]
- Hakata, M.; Kuroda, M.; Miyashita, T.; Yamaguchi, T.; Kojima, M.; Sakakibara, H.; Mitsui, T.; Yamakawa, H. Suppression of alpha-amylase genes improves quality of rice grain ripened under high temperature. Plant Biotechnol. J. 2012, 10, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- James, M.G.; Denyer, K.; Myers, A.M. Starch synthesis in the cereal endosperm. Curr. Opin. Plant Biol. 2003, 6, 215–222. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry & Molecular Biology of Plants; American Society of Plant Physiologists: Rockville, MD, USA, 2015. [Google Scholar]
- Scofield, G.N.; Ruuska, S.A.; Aoki, N.; Lewis, D.C.; Tabe, L.M.; Jenkins, C.L.D. Starch storage in the stems of wheat plants: Localization and temporal changes. Ann. Bot. 2009, 103, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Van den Ende, W.; Schroeven, L.; Clerens, S.; Geuten, K.; Cheng, S.; Bennett, J. The rice genome encodes two vacuolar invertases with fructan exohydrolase activity but lacks the related fructan biosynthesis genes of the Pooideae. New Phytol. 2007, 173, 50–62. [Google Scholar] [CrossRef]
- Matsushima, R. Morphological Variations of Starch Grains. In Starch: Metabolism and Structure; Nakamura, Y., Ed.; Springer: Tokyo, Japan, 2015; pp. 425–441. [Google Scholar] [CrossRef]
- Matsushima, R.; Maekawa, M.; Sakamoto, W. Geometrical Formation of Compound Starch Grains in Rice Implements Voronoi Diagram. Plant Cell Physiol. 2015, 56, 2150–2157. [Google Scholar] [CrossRef]
- Matsushima, R.; Maekawa, M.; Fujita, N.; Sakamoto, W. A Rapid, Direct Observation Method to Isolate Mutants with Defects in Starch Grain Morphology in Rice. Plant Cell Physiol. 2010, 51, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef]
- Foolad, M.R.; Subbiah, P.; Zhang, L. Common QTL Affect the Rate of Tomato Seed Germination under Different Stress and Nonstress Conditions. Int. J. Plant Genom. 2007, 2007, 97386. [Google Scholar] [CrossRef] [PubMed]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Macovei, A.; Balestrazzi, A.; Confalonieri, M.; Fae, M.; Carbonera, D. New insights on the barrel medic MtOGG1 and MtFPG functions in relation to oxidative stress response in planta and during seed imbibition. Plant Physiol. Biochem. PPB 2011, 49, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Dandoy, E.; Schnys, R.; Deltour, R.; Verly, W.G. Appearance and repair of apurinic/apyrimidinic sites in DNA during early germination of Zea mays. Mutat. Res./Fundam. Mol. Mech. Mutagen. 1987, 181, 57–60. [Google Scholar] [CrossRef]
- Howell, K.A.; Millar, A.H.; Whelan, J. Ordered assembly of mitochondria during rice germination begins with pro-mitochondrial structures rich in components of the protein import apparatus. Plant Mol. Biol. 2006, 60, 201–223. [Google Scholar] [CrossRef] [PubMed]
- Dinkova, T.D.; Márquez-Velázquez, N.A.; Aguilar, R.; Lázaro-Mixteco, P.E.; Sánchez de Jiménez, E. Tight translational control by the initiation factors eIF4E and eIF(iso)4E is required for maize seed germination. Seed Sci. Res. 2011, 21, 85–93. [Google Scholar] [CrossRef]
- Yang, P.; Li, X.; Wang, X.; Chen, H.; Chen, F.; Shen, S. Proteomic analysis of rice (Oryza sativa) seeds during germination. Proteomics 2007, 7, 3358–3368. [Google Scholar] [CrossRef]
- Lee, J.; Koh, H.-J. A label-free quantitative shotgun proteomics analysis of rice grain development. Proteome Sci. 2011, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.F.; Ho, T.H.; Wu, C.F.; Ho, S.L.; Yeh, R.H.; Lu, C.A.; Chen, P.W.; Yu, L.C.; Chao, A.; Yu, S.M. Convergent starvation signals and hormone crosstalk in regulating nutrient mobilization upon germination in cereals. Plant Cell 2012, 24, 2857–2873. [Google Scholar] [CrossRef]
- Matsukura, C.-A.; Saitoh, T.; Hirose, T.; Ohsugi, R.; Perata, P.; Yamaguchi, J. Sugar Uptake and Transport in Rice Embryo. Expression of Companion Cell-Specific Sucrose Transporter (OsSUT1) Induced by Sugar and Light. Plant Physiol. 2000, 124, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Duermeyer, L.; Leoveanu, C.; Nambara, E. The functions of the endosperm during seed germination. Plant Cell Physiol. 2014, 55, 1521–1533. [Google Scholar] [CrossRef] [PubMed]
- West, C.E.; Waterworth, W.M.; Stephens, S.M.; Smith, C.P.; Bray, C.M. Cloning and functional characterisation of a peptide transporter expressed in the scutellum of barley grain during the early stages of germination. Plant J. Cell Mol. Biol. 1998, 15, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Pujadas, G.; Palau, J. Evolution of alpha-amylases: Architectural features and key residues in the stabilization of the (β/α)(8) scaffold. Mol. Biol. Evol. 2001, 18, 38–54. [Google Scholar] [CrossRef]
- Yamaguchi, J.; Itoh, S.; Saitoh, T.; Ikeda, A.; Tashiro, T.; Nagato, Y. Characterization of β-amylase and its deficiency in various rice cultivars. Theor. Appl. Genet. 1999, 98, 32–38. [Google Scholar] [CrossRef]
- Sugimoto, N.; Takeda, G.; Nagato, Y.; Yamaguchi, J. Temporal and Spatial Expression of the α-Amylase Gene during Seed Germination in Rice and Barley. Plant Cell Physiol. 1998, 39, 323–333. [Google Scholar] [CrossRef]
- Wang, S.-M.; Lue, W.-L.; Eimert, K.; Chen, J. Phytohormone-regulated β-amylase gene expression in rice. Plant Mol. Biol. 1996, 31, 975–982. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Han, C.; Yao, J.; Shen, S.; Yang, P. Constructing the metabolic and regulatory pathways in germinating rice seeds through proteomic approach. Proteomics 2011, 11, 2693–2713. [Google Scholar] [CrossRef]
- Galland, M.; He, D.; Lounifi, I.; Arc, E.; Clément, G.; Balzergue, S.; Huguet, S.; Cueff, G.; Godin, B.; Collet, B.; et al. An Integrated “Multi-Omics” Comparison of Embryo and Endosperm Tissue-Specific Features and Their Impact on Rice Seed Quality. Front. Plant Sci. 2017, 8, 1984. [Google Scholar] [CrossRef]
- Howell, K.A.; Narsai, R.; Carroll, A.; Ivanova, A.; Lohse, M.; Usadel, B.; Millar, A.H.; Whelan, J. Mapping Metabolic and Transcript Temporal Switches during Germination in Rice Highlights Specific Transcription Factors and the Role of RNA Instability in the Germination Process. Plant Physiol. 2009, 149, 961–980. [Google Scholar] [CrossRef]
- Rajjou, L.; Gallardo, K.; Debeaujon, I.; Vandekerckhove, J.; Job, C.; Job, D. The effect of alpha-amanitin on the Arabidopsis seed proteome highlights the distinct roles of stored and neosynthesized mRNAs during germination. Plant Physiol. 2004, 134, 1598–1613. [Google Scholar] [CrossRef]
- Shingaki-Wells, R.N.; Huang, S.; Taylor, N.L.; Carroll, A.J.; Zhou, W.; Millar, A.H. Differential molecular responses of rice and wheat coleoptiles to anoxia reveal novel metabolic adaptations in amino acid metabolism for tissue tolerance. Plant Physiol. 2011, 156, 1706–1724. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed Germination and Vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Tuncel, A.; Cakir, B.; Hwang, S.K.; Okita, T.W. The role of the large subunit in redox regulation of the rice endosperm ADP-glucose pyrophosphorylase. FEBS J. 2014, 281, 4951–4963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saulnier, L.; Guillon, F.; Sado, P.E.; Rouau, X. 2.20—Plant Cell Wall Polysaccharides in Storage Organs: Xylans (Food Applications). In Comprehensive Glycoscience; Kamerling, H., Ed.; Elsevier: Oxford, UK, 2007; pp. 653–689. [Google Scholar] [CrossRef]
- Henrissat, B. A Classification of glycosyl hydrolases based on amino-acid-sequence similarities. Biochem. J. 1991, 280, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Henrissat, B.; Bairoch, A. Updating the sequence-based classification of glycosyl hydrolases. Biochem. J. 1996, 316, 695–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majzlova, K.; Pukajova, Z.; Janecek, S. Tracing the evolution of the alpha-amylase subfamily GH13_36 covering the amylolytic enzymes intermediate between oligo-1,6-glucosidases and neopullulanases. Carbohydr. Res. 2013, 367, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Stebbins, G.L.; Rodriguez, R.L. Classification and evolution of α-amylase genes in plants. Proc. Natl. Acad. Sci. USA 1992, 89, 7526–7530. [Google Scholar] [CrossRef]
- Thomas, B.R.; Terashima, M.; Katoh, S.; Stoltz, T.; Rodriguez, R.L. Metabolic Regulation of Source-Sink Relations in Cereal Seedlings; Food and Agriculture Organization: Rome, Italy, 1995; Volume 13, pp. 78–90. [Google Scholar]
- Miyata, S.; Akazawa, T. Enzymic Mechanism of Starch Breakdown in Germinating Rice Seeds. 12. Biosynthesis of Alpha-Amylase in Relation to Protein Glycosylation. Plant Physiol. 1982, 70, 147–153. [Google Scholar] [CrossRef]
- Daussant, J.; Miyata, S.; Mitsui, T.; Akazawa, T. Enzymic Mechanism of Starch Breakdown in Germinating Rice Seeds.15. Immunochemical Study on Multiple Forms of Amylase. Plant Physiol. 1983, 71, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, T.; Yamaguchi, J.; Akazawa, T. Physicochemical and serological characterization of rice α-amylase isoforms and identification of their corresponding genes. Plant Physiol. 1996, 110, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Nanjo, Y.; Asatsuma, S.; Itoh, K.; Hori, H.; Mitsui, T. Proteomic identification of alpha-amylase isoforms encoded by RAmy3B/3C from germinating rice seeds. Biosci. Biotechnol. Biochem. 2004, 68, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.V.; Higgins, T.J.V. Characterization of the α-amylases synthesized by aleurone layers of himalaya barley in response to gibberellic-acid. Plant Physiol. 1982, 70, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Huang, X.; Shen, Y.; Huang, Z. A novel wheat -amylase inhibitor gene, TaHPS, significantly improves the salt and drought tolerance of transgenic Arabidopsis. Physiol. Plant. 2013, 148, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Gale, M.D.; Law, C.N.; Chojecki, A.J.; Kempton, R.A. Genetic-control of α-amylase production in wheat. Theor. Appl. Genet. 1983, 64, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Karrer, E.E.; Litts, J.C.; Rodriguez, R.L. Differential expression of alpha-amylase genes in germinating rice and barley-seeds. Plant Mol. Biol. 1991, 16, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Fu, Y.; Guan, Y.; Lin, C.; Cao, D.; Hu, W.; Sheteiwy, M.; Hu, J. Inhibitory effect of chemical combinations on seed germination and pre-harvest sprouting in hybrid rice. Plant Growth Regul. 2016, 80, 281–289. [Google Scholar] [CrossRef]
- Nakata, M.; Fukamatsu, Y.; Miyashita, T.; Hakata, M.; Kimura, R.; Nakata, Y.; Kuroda, M.; Yamaguchi, T.; Yamakawa, H. High Temperature-Induced Expression of Rice alpha-Amylases in Developing Endosperm Produces Chalky Grains. Front. Plant Sci. 2017, 8, 2089. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.W.; Lu, C.A.; Yu, T.S.; Tseng, T.H.; Wang, C.S.; Yu, S.M. Rice α-amylase transcriptional enhancers direct multiple mode regulation of promoters in transgenic rice. J. Biol. Chem. 2002, 277, 13641–13649. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.-S.; Karrer, E.E.; Thomas, B.R.; Chen, L.; Rodriguez, R.L. Three cis-elements required for rice α-amylase Amy3D expression during sugar starvation. Plant Mol. Biol. 1998, 36, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.T.; Tran, A.N.; Cardarelli, F.; Perata, P.; Pucciariello, C. A calcineurin B-like protein participates in low oxygen signalling in rice. Funct. Plant Biol. 2017, 44, 917–928. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, A.; Sugai, H.; Harada, K.; Tanaka, S.; Ishiyama, Y.; Ito, K.; Tanaka, T.; Uchiumi, T.; Taniguchi, M.; Mitsui, T. Crystal structure of α-amylase from Oryza sativa: Molecular insights into enzyme activity and thermostability. Biosci. Biotechnol. Biochem. 2014, 78, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.M.; Bozonnet, S.; Seo, E.S.; Motyan, J.A.; Andersen, J.M.; Dilokpimol, A.; Abou Hachem, M.; Gyemant, G.; Naested, H.; Kandra, L.; et al. Two secondary carbohydrate binding sites on the surface of barley alpha-amylase 1 have distinct functions and display synergy in hydrolysis of starch granules. Biochemistry 2009, 48, 7686–7697. [Google Scholar] [CrossRef] [PubMed]
- Gubler, F.; Jacobsen, J.V. Gibberellin-responsive elements in the promoter of a barley high-PL α-amylase gene. Plant Cell 1992, 4, 1435–1441. [Google Scholar] [PubMed]
- Washio, K. Functional dissections between GAMYB and Dof transcription factors suggest a role for protein-protein associations in the gibberellin-mediated expression of the RAmy1A gene in the rice aleurone. Plant Physiol. 2003, 133, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Sutliff, T.D.; Litts, J.C.; Rodriguez, R.L. Classification and characterization of the rice α-amylase multigene family. Plant Mol. Biol. 1990, 14, 655–668. [Google Scholar] [CrossRef]
- Gubler, F.; Raventos, D.; Keys, M.; Watts, R.; Mundy, J.; Jacobsen, J.V. Target genes and regulatory domains of the GAMYB transcriptional activator in cereal aleurone. Plant J. 1999, 17, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.A.; Lim, E.K.; Yu, S.M. Sugar response sequence in the promoter of a rice alpha-amylase gene serves as a transcriptional enhancer. J. Biol. Chem. 1998, 273, 10120–10131. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, C. Comparisons of Copy Number, Genomic Structure, and Conserved Motifs for α-Amylase Genes from Barley, Rice, and Wheat. Front. Plant Sci. 2017, 8, 1727. [Google Scholar] [CrossRef]
- Sato, Y.; Antonio, B.A.; Namiki, N.; Takehisa, H.; Minami, H.; Kamatsuki, K.; Sugimoto, K.; Shimizu, Y.; Hirochika, H.; Nagamura, Y. RiceXPro: A platform for monitoring gene expression in japonica rice grown under natural field conditions. Nucleic Acids Res. 2011, 39, D1141–D1148. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-J.; Wang, S.-J. Molecular regulation of sink-source transition in rice leaf sheaths during the heading period. Acta Physiol. Plant. 2008, 30, 639–649. [Google Scholar] [CrossRef]
- Barrero, J.M.; Mrva, K.; Talbot, M.J.; White, R.G.; Taylor, J.; Gubler, F.; Mares, D.J. Genetic, Hormonal, and Physiological Analysis of Late Maturity alpha-Amylase in Wheat. Plant Physiol. 2013, 161, 1265–1277. [Google Scholar] [CrossRef]
- Huang, N.; Chandler, J.; Thomas, B.R.; Koizumi, N.; Rodriguez, R.L. Metabolic-regulation of alpha-amylase gene-expression in transgenic cell-cultures of rice (Oryza-sativa L). Plant Mol. Biol. 1993, 23, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Karrer, E.E.; Rodriguez, R.L. Metabolic-regulation of rice α-amylase and sucrose synthase genes inplanta. Plant J. 1992, 2, 517–523. [Google Scholar] [PubMed]
- Subbarao, K.V.; Datta, R.; Sharma, R. Amylases synthesis in scutellum and aleurone layer of maize seeds. Phytochemistry 1998, 49, 657–666. [Google Scholar] [CrossRef]
- Gubler, F.; Kalla, R.; Roberts, J.K.; Jacobsen, J.V. Gibberellin-regulated expression of a myb gene in barley aleurone cells: Evidence for Myb transactivation of a high-pI α-amylase gene promoter. Plant Cell 1995, 7, 1879–1891. [Google Scholar] [CrossRef] [PubMed]
- Scofield, G.N.; Aoki, N.; Hirose, T.; Takano, M.; Jenkins, C.L.; Furbank, R.T. The role of the sucrose transporter, OsSUT1, in germination and early seedling growth and development of rice plants. J. Exp. Bot. 2007, 58, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Aoki, N.; Scofield, G.N.; Wang, X.-D.; Offler, C.E.; Patrick, J.W.; Furbank, R.T. Pathway of Sugar Transport in Germinating Wheat Seeds. Plant Physiol. 2006, 141, 1255–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Nambara, E.; Okamoto, M.; Tatematsu, K.; Yano, R.; Seo, M.; Kamiya, Y. Abscisic acid and the control of seed dormancy and germination. Seed Sci. Res. 2010, 20, 55–67. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Min, M.K.; Choi, E.H.; Kim, N.; Moon, S.J.; Yoon, I.; Kwon, T.; Jung, K.H.; Kim, B.G. The protein phosphatase 2C clade A protein OsPP2C51 positively regulates seed germination by directly inactivating OsbZIP10. Plant Mol. Biol. 2017, 93, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.W.; Chiang, C.M.; Tseng, T.H.; Yu, S.M. Interaction between rice MYBGA and the gibberellin response element controls tissue-specific sugar sensitivity of alpha-amylase genes. Plant Cell 2006, 18, 2326–2340. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Itoh, H.; Ueguchi-Tanaka, M.; Ashikari, M.; Matsuoka, M. The alpha-amylase induction in endosperm during rice seed germination is caused by gibberellin synthesized in epithelium. Plant Physiol. 2002, 128, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Jelsema, C.L.; Ruddat, M.; Morre, D.J.; Williamson, F.A. Specific binding of gibberellin a-1 to aleurone grain fractions from wheat endosperm. Plant Cell Physiol. 1977, 18, 1009–1019. [Google Scholar]
- Lovegrove, A.; Barratt, D.H.P.; Beale, M.H.; Hooley, R. Gibberellin-photoaffinity labelling of two polypeptides in plant plasma membranes. Plant J. 1998, 15, 311–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooley, R.; Beale, M.H.; Smith, S.J. Gibberellin perception at the plasma-membrane of avena-fatua aleurone protoplasts. Planta 1991, 183, 274–280. [Google Scholar] [CrossRef]
- Gilroy, S.; Jones, R.L. Perception of gibberellin and abscisic-acid at the external face of the plasma-membrane of barley (hordeum-vulgare l) aleurone protoplasts. Plant Physiol. 1994, 104, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Aya, K.; Hirano, K.; Ordonio, R.L.; Ueguchi-Tanaka, M.; Matsuoka, M. Comprehensive Gene Expression Analysis of Rice Aleurone Cells: Probing the Existence of an Alternative Gibberellin Receptor. Plant Physiol. 2015, 167, 531–544. [Google Scholar] [CrossRef]
- Sun, T.P.; Gubler, F. Molecular mechanism of gibberellin signaling in plants. Annu. Rev. Plant Biol. 2004, 55, 197–223. [Google Scholar] [CrossRef]
- Ueguchi-Tanaka, M.; Nakajima, M.; Katoh, E.; Ohmiya, H.; Asano, K.; Saji, S.; Hongyu, X.; Ashikari, M.; Kitano, H.; Yamaguchi, I.; et al. Molecular Interactions of a Soluble Gibberellin Receptor, GID1, with a Rice DELLA Protein, SLR1, and Gibberellin. Plant Cell 2007, 19, 2140–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakura, H.; Kanei-Ishii, C.; Nagase, T.; Nakagoshi, H.; Gonda, T.J.; Ishii, S. Delineation of three functional domains of the transcriptional activator encoded by the c-myb protooncogene. Proc. Natl. Acad. Sci. USA 1989, 86, 5758–5762. [Google Scholar] [CrossRef]
- Kaneko, M.; Inukai, Y.; Ueguchi-Tanaka, M.; Itoh, H.; Izawa, T.; Kobayashi, Y.; Hattori, T.; Miyao, A.; Hirochika, H.; Ashikari, M.; et al. Loss-of-function mutations of the rice GAMYB gene impair alpha-amylase expression in aleurone and flower development. Plant Cell 2004, 16, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Yamaguchi, J.; Huang, N.; Rodriguez, R.L.; Akazawa, T.; Shimamoto, K. Developmental and Hormonal Regulation of Rice [alpha]-Amylase(RAmy1A)-gusA Fusion Genes in Transgenic Rice Seeds. Plant Physiol. 1995, 107, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.A.; Ho, T.H.D.; Ho, S.L.; Yu, S.M. Three novel MYB proteins with one DNA binding repeat mediate sugar and hormone regulation of alpha-amylase gene expression. Plant Cell 2002, 14, 1963–1980. [Google Scholar] [CrossRef]
- Yu, S.-M.; Lee, Y.-C.; Fang, S.-C.; Chan, M.-T.; Hwa, S.-F.; Liu, L.-F. Sugars Act as Signal Molecules and Osmotica to Regulate the Expression of Amylase Genes and Metabolic Activities in Germinating Cereal Grains. Plant Mol. Biol. 1996, 30, 1277–1289. [Google Scholar] [CrossRef]
- Perata, P.; Matsukura, C.; Vernieri, P.; Yamaguchi, J. Sugar Repression of a Gibberellin-Dependent Signaling Pathway in Barley Embryos. Plant Cell 1997, 9, 2197–2208. [Google Scholar] [CrossRef]
- Yu, S.; Tzou, W.-S.; Lo, W.; Kuo, Y.; Lee, H.; Wu, R. Regulation of α-Amylase-Encoding Gene Expression in Germinating Seeds and Cultured Cells of Rice. Gene 1993, 122, 247–253. [Google Scholar]
- Yu, S.M.; Kuo, Y.H.; Sheu, G.; Sheu, Y.J.; Liu, L.F. Metabolic derepression of alpha-amylase gene expression in suspension-cultured cells of rice. J. Biol. Chem. 1991, 266, 21131–21137. [Google Scholar]
- Chan, M.T.; Chao, Y.C.; Yu, S.M. Novel gene expression system for plant cells based on induction of alpha-amylase promoter by carbohydrate starvation. J. Biol. Chem. 1994, 269, 17635–17641. [Google Scholar]
- Sheu, J.J.; Yu, T.S.; Tong, W.F.; Yu, S.M. Carbohydrate starvation stimulates differential expression of rice alpha-amylase genes that is modulated through complicated transcriptional and posttranscriptional processes. J. Biol. Chem. 1996, 271, 26998–27004. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.-J.; Jan, S.-P.; Lee, H.-T.; Yu, S.-M. Control of transcription and mRNA turnover as mechanisms of metabolic repression of α-amylase gene expression. Plant J. 1994, 5, 655–664. [Google Scholar] [CrossRef]
- Chan, M.-T.; Yu, S.-M. The 3′ untranslated region of a rice α-amylase gene mediates sugar-dependent abundance of mRNA. Plant J. 1998, 15, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.; Chen, P.W.; Lu, C.A.; Chen, S.; Ho, T.H.; Yu, S.M. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Chao, Y.C.; Tseng, T.W.; Huang, C.K.; Lo, P.C.; Lu, C.A. Two MYB-related transcription factors play opposite roles in sugar signaling in Arabidopsis. Plant Mol. Biol. 2017, 93, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.R.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Xia, W.; Li, H.; Zeng, H.; Wei, B.; Han, S.; Yin, C. Salinity Inhibits Rice Seed Germination by Reducing Alpha-Amylase Activity via Decreased Bioactive Gibberellin Content. Front. Plant Sci. 2018, 9, 275. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, D.-G.; Lee, S.-H.; Kang, K.Y.; Lee, J.J.; Kim, P.J.; Yoon, H.-S.; Kim, J.-S.; Lee, B.-H. Excess copper induced physiological and proteomic changes in germinating rice seeds. Chemosphere 2007, 67, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Zhang, S.Z.; Shan, X.Q.; Zhu, Y.G. Toxicity of arsenate and arsenite on germination, seedling growth and amylolytic activity of wheat. Chemosphere 2005, 61, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perata, P.; Pozuetaromero, J.; Akazawa, T.; Yamaguchi, J. Effect of anoxia on starch breakdown in rice and wheat seeds. Planta 1992, 188, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, Y.; Kato-Noguchi, H. Effects of benzoxazinoids in wheat residues may inhibit the germination, growth and gibberellin-induced α-amylase activity in rice. Acta Physiol. Plant. 2015, 38, 24. [Google Scholar] [CrossRef]
- Muralikrishna, G.; Nirmala, M. Cereal α-amylases—An overview. Carbohydr. Polym. 2005, 60, 163–173. [Google Scholar] [CrossRef]
- Asatsuma, S.; Sawada, C.; Kitajima, A.; Asakura, T.; Mitsui, T. ALPHA.-Amylase Affects Starch Accumulation in Rice Grains. J. Appl. Glycosci. 2006, 53, 187–192. [Google Scholar] [CrossRef]
- Gubler, F.; Millar, A.A.; Jacobsen, J.V. Dormancy release, ABA and pre-harvest sprouting. Curr. Opin. Plant Biol. 2005, 8, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Ral, J.P.; Whan, A.; Larroque, O.; Leyne, E.; Pritchard, J.; Dielen, A.S.; Howitt, C.A.; Morell, M.K.; Newberry, M. Engineering high α-amylase levels in wheat grain lowers Falling Number but improves baking properties. Plant Biotechnol. J. 2016, 14, 364–376. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Subfamily | Group | Regulation | Expression Stage | Expression Site |
|---|---|---|---|---|---|
| RAmy1A RAmy1B | RAmy1 | 1 | Phytohormones GA, ABA, sugar repression Chemical repression [50] High temperature during rice seed maturation [4] | Seed development [51] Seed germination [45] | Aleurone layer [52] Endosperm [51] |
| RAMY1C | RAmy1 | 1 | High temperature during rice seed maturation [4] | Entire growth period * | Aleurone layer [52] All tissues * |
| RAmy2A | RAmy2 | 4 | NF | Entire growth period * | Leaf sheath [2] All tissues * |
| RAmy3A | - | 3 | High temperature during rice seed maturation [4] | Seed development * | Seed maturation (endosperm) |
| RAmy3B | - | 3 | Chemicals repression [50] | Seed germination [45] | Aleurone layer [52] |
| RAmy3C | - | 3 | NF | Seed germination [45] Seed development [51] | Endosperm [51] Leaf sheath [2] |
| RAmy3D | RAmy3 | 2 | Sugar repression [53] Calcium [54] High temperature during rice seed maturation [4] | Seed development [51] Seed germination [45] | Embryo [49] Aleurone layer [51] |
| RAmy3E | RAmy3 | 5 | Chemical repression [50] High temperature during rice seed maturation [4] | Seed development [51] Seed germination [45] | Aleurone layer [52] |
| RAmy3F | RAmy3 | 6 | NF | Entire growth period * | All tissues * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damaris, R.N.; Lin, Z.; Yang, P.; He, D. The Rice Alpha-Amylase, Conserved Regulator of Seed Maturation and Germination. Int. J. Mol. Sci. 2019, 20, 450. https://doi.org/10.3390/ijms20020450
Damaris RN, Lin Z, Yang P, He D. The Rice Alpha-Amylase, Conserved Regulator of Seed Maturation and Germination. International Journal of Molecular Sciences. 2019; 20(2):450. https://doi.org/10.3390/ijms20020450
Chicago/Turabian StyleDamaris, Rebecca Njeri, Zhongyuan Lin, Pingfang Yang, and Dongli He. 2019. "The Rice Alpha-Amylase, Conserved Regulator of Seed Maturation and Germination" International Journal of Molecular Sciences 20, no. 2: 450. https://doi.org/10.3390/ijms20020450
APA StyleDamaris, R. N., Lin, Z., Yang, P., & He, D. (2019). The Rice Alpha-Amylase, Conserved Regulator of Seed Maturation and Germination. International Journal of Molecular Sciences, 20(2), 450. https://doi.org/10.3390/ijms20020450