Expression Analysis of mRNA Decay of Maternal Genes during Bombyx mori Maternal-to-Zygotic Transition

Abstract
:1. Introduction
2. Results
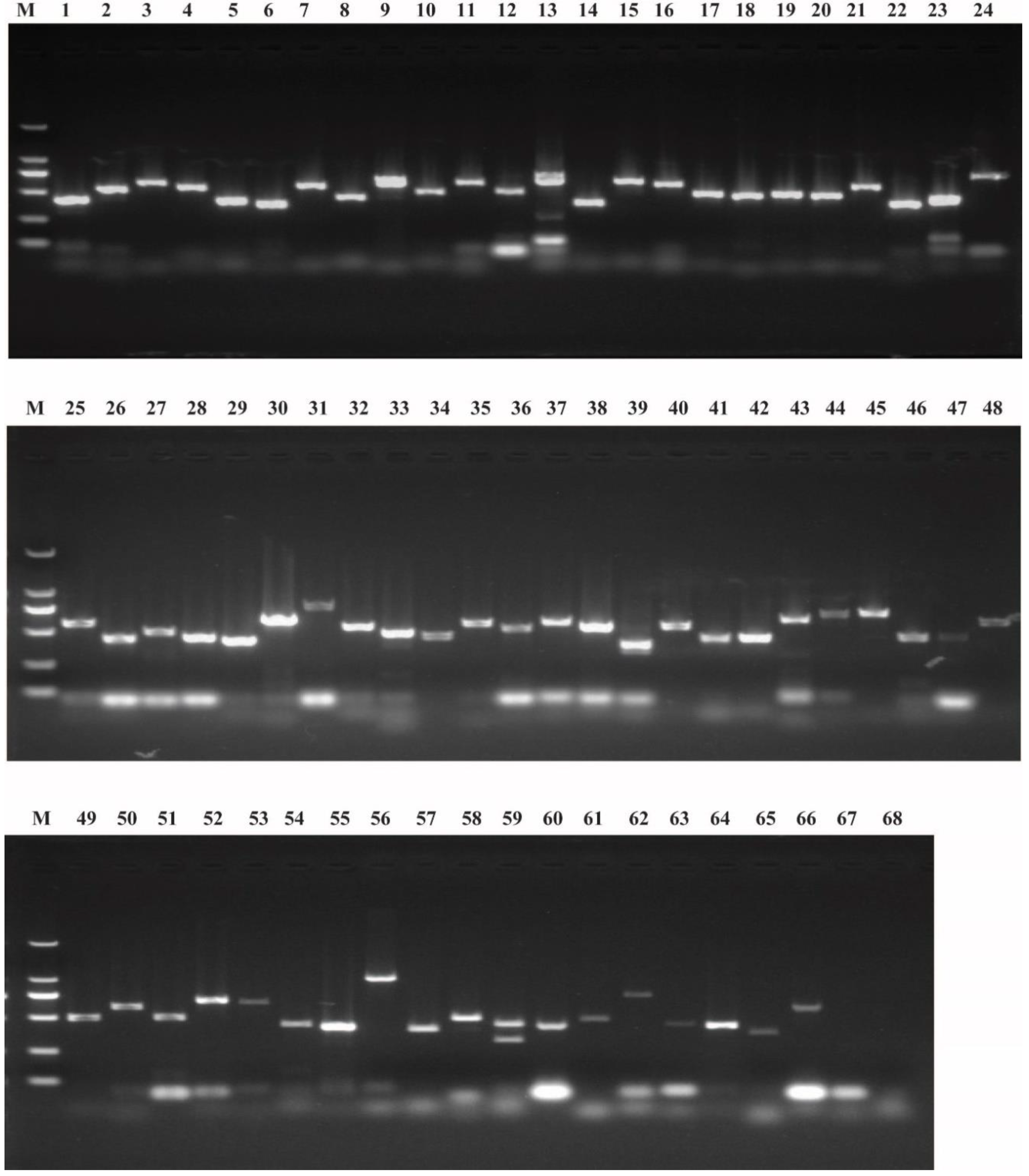
2.1. Identification of Potential Maternal Genes
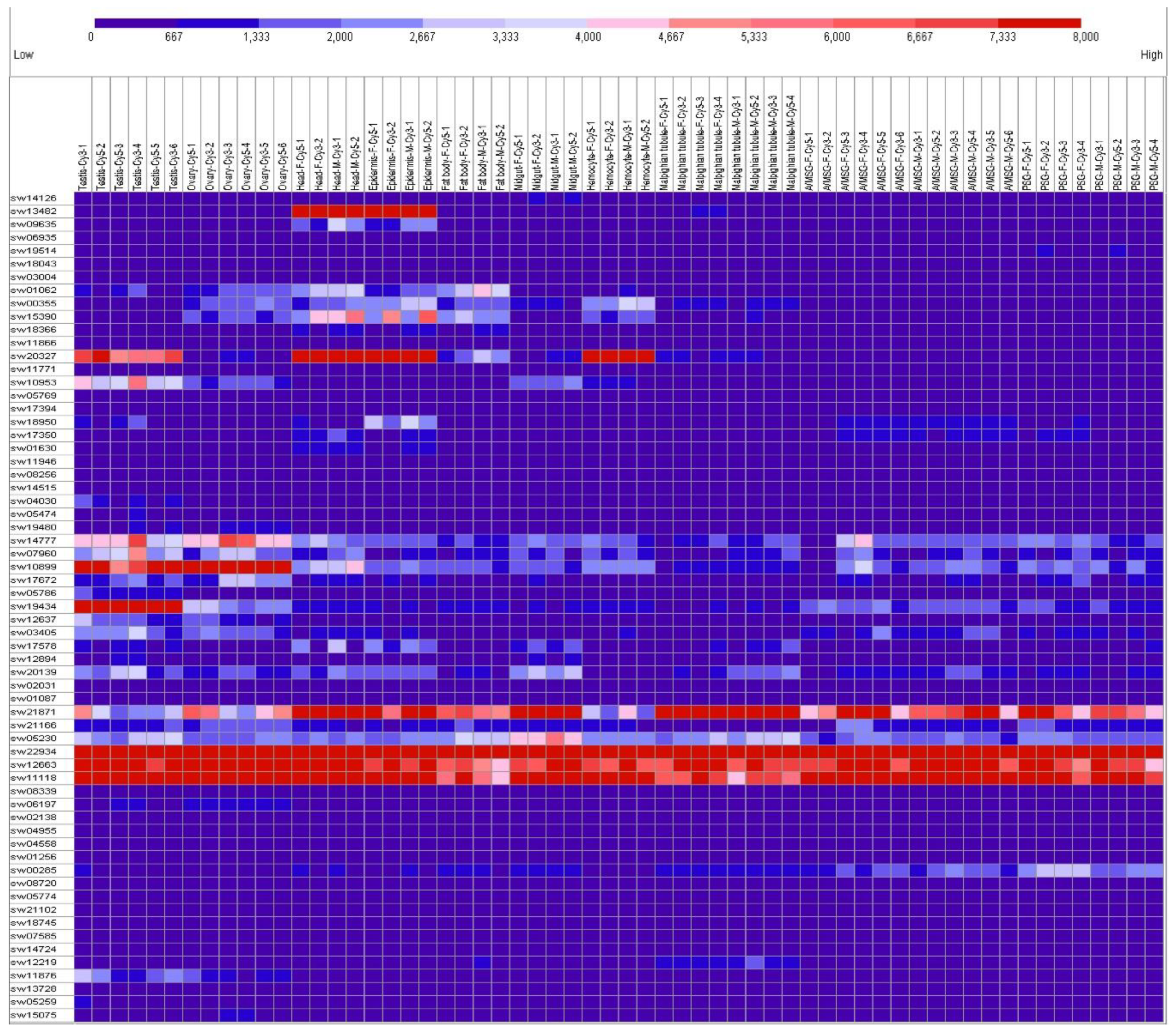
2.2. Tissue Expression Patterns on Day 3 of the Fifth Instar
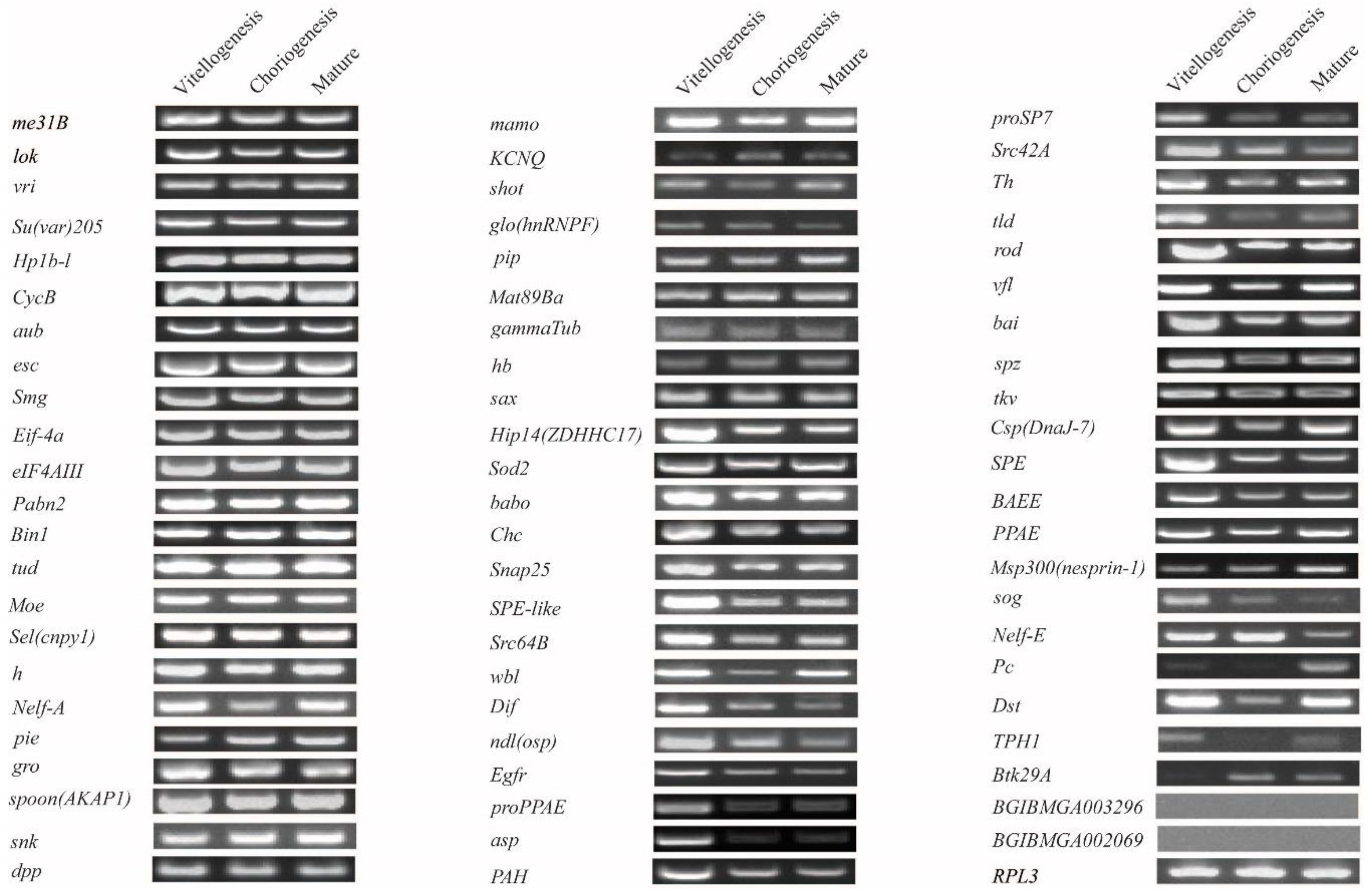
2.3. Expression Analysis in Developing Oocytes in Day 8 Pupae
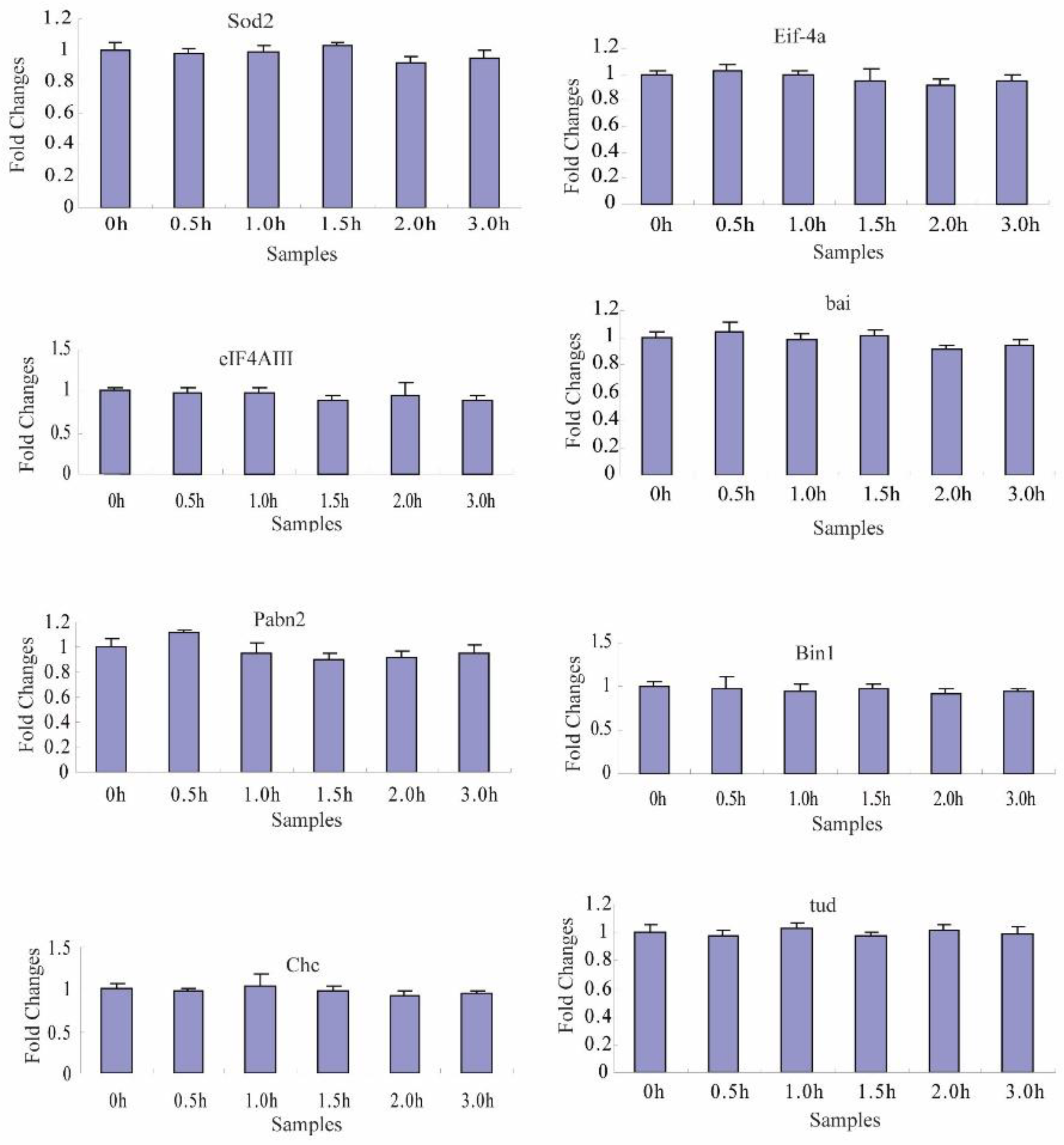
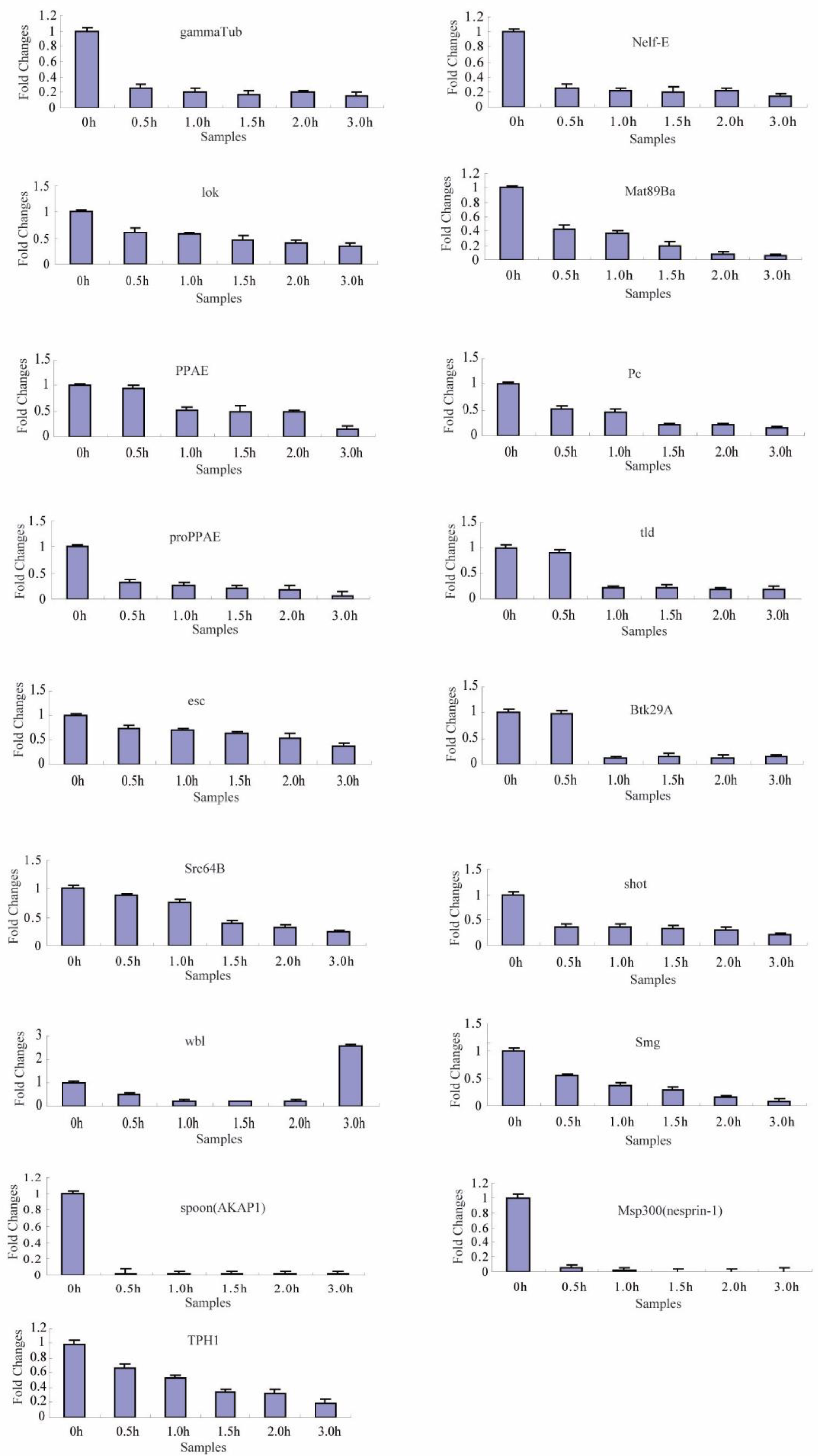
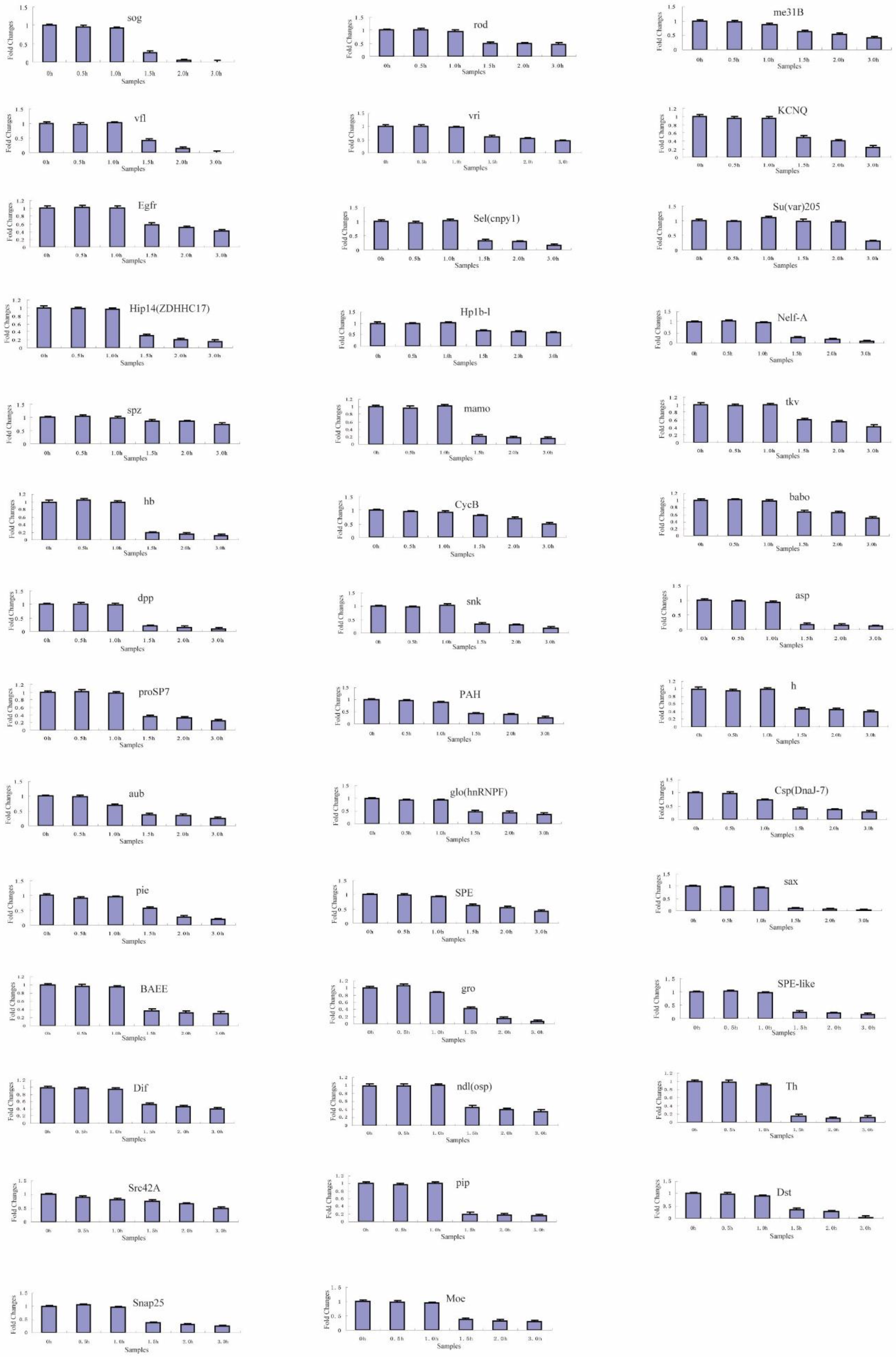
2.4. Transcriptional Degradation during the Maternal-to-Zygotic Transition
3. Discussion
4. Materials and Methods
4.1. Insects and Collection of Samples
4.2. Identification of B. mori Maternal Genes
4.3. Transcript Detection Reverse Transcription-PCR
4.4. Tissue Expression Patterns Based on Microarray Database
4.5. Transcript DecayDetection by Reverse Transcription-Quantitative PCR
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Horner, V.L.; Wolfner, M.F. Mechanical stimulation by osmotic and hydrostatic pressure activates Drosophila oocytes in vitro in a calcium-dependent manner. Dev. Biol. 2008, 316, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Tadros, W.; Lipshitz, H.D. The maternal-to-zygotic transition: A play in two acts. Development 2009, 136, 3033–3042. [Google Scholar] [CrossRef] [PubMed]
- Marlow, F.L. Maternal Control of Development in Vertebrates: My Mother Made Me Do It; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010. [Google Scholar]
- Yartseva, V.; Giraldez, A.J. The Maternal-to-Zygotic Transition During Vertebrate Development: A Model for Reprogramming. Curr. Top. Dev. Biol. 2015, 113, 191–232. [Google Scholar] [PubMed]
- Giraldez, A.J.; Mishima, Y.; Rihel, J.; Grocock, R.J.; Van Dongen, S.; Inoue, K.; Enright, A.J.; Schier, A.F. Zebrafish MiR-430 promotes deadenylation and clearance of maternal mRNAs. Science 2006, 312, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Langley, A.R.; Smith, J.C.; Stemple, D.L.; Harvey, S.A. New insights into the maternal to zygotic transition. Development 2014, 141, 3834–3841. [Google Scholar] [CrossRef] [PubMed]
- Walser, C.B.; Lipshitz, H.D. Transcript clearance during the maternal-to-zygotic transition. Curr. Opin. Genet. Dev. 2011, 21, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Bashirullah, A.; Halsell, S.R.; Cooperstock, R.L.; Kloc, M.; Karaiskakis, A.; Fisher, W.W.; Fu, W.; Hamilton, J.K.; Etkin, L.D.; Lipshitz, H.D. Joint action of two RNA degradation pathways controls the timing of maternal transcript elimination at the midblastula transition in Drosophila melanogaster. EMBO J. 1999, 18, 2610–2620. [Google Scholar] [CrossRef] [PubMed]
- Smibert, C.A.; Wilson, J.E.; Kerr, K.; Macdonald, P.M. smaug protein represses translation of unlocalized nanos mRNA in the Drosophila embryo. Genes Dev. 1996, 10, 2600–2609. [Google Scholar] [CrossRef] [PubMed]
- Semotok, J.L.; Cooperstock, R.L.; Pinder, B.D.; Vari, H.K.; Lipshitz, H.D.; Smibert, C.A. Smaug recruits the CCR4/POP2/NOT deadenylase complex to trigger maternal transcript localization in the early Drosophila embryo. Curr. Biol. 2005, 15, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Tadros, W.; Goldman, A.L.; Babak, T.; Menzies, F.; Vardy, L.; Orr-Weaver, T.; Hughes, T.R.; Westwood, J.T.; Smilbert, C.A.; Lipshitz, H.D. SMAUG is a major regulator of maternal mRNA destabilization in Drosophila and its translation is activated by the PAN GU kinase. Dev. Cell 2007, 12, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, S.; Anders, S.; Janga, S.C.; Huber, W.; Alonso, C.R. Genome-wide analysis of mRNA decay patterns during early Drosophila development. Genome Biol. 2010, 11, R93. [Google Scholar] [CrossRef] [PubMed]
- Jeske, M.; Moritz, B.; Anders, A.; Wahle, E. Smaug assembles an ATP-dependent stable complex repressing nanos mRNA translation at multiple levels. Embo J. 2011, 30, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, P.M. The Drosophila pumilio gene: An unusually long transcription unit and an unusual protein. Development 1992, 114, 221–232. [Google Scholar] [PubMed]
- Zamore, P.D.; Williamson, J.R.; Lehmann, R. The Pumilio protein binds RNA through a conserved domain that defines a new class of RNA-binding proteins. RNA 1997, 3, 1421–1433. [Google Scholar] [PubMed]
- Gamberi, C.; Peterson, D.S.; He, L.; Gottlieb, E. An anterior function for the Drosophila posterior determinant Pumilio. Development 2002, 129, 2699–2710. [Google Scholar] [PubMed]
- Gerber, A.P.; Luschnig, S.; Krasnow, M.A.; Brown, P.O.; Herschlag, D. Genome-wide identification of mRNAs associated with the translational regulator PUMILIO in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2006, 103, 4487–4492. [Google Scholar] [CrossRef] [PubMed]
- De Renzis, S.; Elemento, O.; Tavazoie, S.; Wieschaus, E.F. Unmasking activation of the zygotic genome using chromosomal deletions in the Drosophila embryo. PLoS Biol. 2007, 5, 1036–1051. [Google Scholar] [CrossRef]
- Sonoda, J.; Wharton, R.P. Drosophila Brain Tumor is a translational repressor. Genes Dev. 2001, 15, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Loedige, I.; Stotz, M.; Qamar, S.; Kramer, K.; Hennig, J.; Schubert, T.; Loffler, P.; Langst, G.; Merkl, R.; Urlaub, H.; et al. The NHL domain of BRAT is an RNA-binding domain that directly contacts the hunchback mRNA for regulation. Gene Dev. 2014, 28, 749–764. [Google Scholar] [CrossRef] [PubMed]
- Laver, J.D.; Li, X.; Ray, D.; Cook, K.B.; Hahn, N.A.; Nabeel-Shah, S.; Kekis, M.; Luo, H.; Marsolais, A.J.; Fung, K.Y.Y.; et al. Brain tumor is a sequence-specific RNA-binding protein that directs maternal mRNA clearance during the Drosophila maternal-to-zygotic transition. Genome Biol. 2015, 16, 94. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ly, M.; Lugowski, A.; Laver, J.D.; Lipshitz, H.D.; Smibert, C.A.; Rissland, O.S. ME31B globally represses maternal mRNAs by two distinct mechanisms during the Drosophila maternal-to-zygotic transition. Elife 2017, 6, e27891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallie, D.R. The cap and poly(A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 1991, 5, 2108–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouget, C.; Papin, C.; Boureux, A.; Meunier, A.C.; Franco, B.; Robine, N.; Lai, E.C.; Pelisson, A.; Simonelig, M. Maternal mRNA deadenylation and decay by the piRNA pathway in the early Drosophila embryo. Nature 2010, 467, 1128–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barckmann, B.; Pierson, S.; Dufourt, J.; Papin, C.; Armenise, C.; Port, F.; Grentzinger, T.; Chambeyron, S.; Baronian, G.; Desvignes, J.P.; et al. Aubergine iCLIP Reveals piRNA-Dependent Decay of mRNAs Involved in Germ Cell Development in the Early Embryo. Cell Rep. 2015, 12, 1205–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Li, X.; Claycomb, J.M.; Lipshitz, H.D. The Smaug RNA-Binding Protein Is Essential for microRNA Synthesis During the Drosophila Maternal-to-Zygotic Transition. G3 Genes Genomes Genet. 2016, 6, 3541–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laver, J.D.; Marsolais, A.J.; Smibert, C.A.; Lipshitz, H.D. Regulation and Function of Maternal Gene Products During the Maternal-to-Zygotic Transition in Drosophila. Curr. Top. Dev. Biol. 2015, 113, 43–84. [Google Scholar] [PubMed]
- Niinuma, S.; Tomari, Y. ATP is dispensable for both miRNA- and Smaug-mediated deadenylation reactions. RNA 2017, 23, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol. Cell Pharmacol. 2011, 3, 83–92. [Google Scholar] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bushati, N.; Stark, A.; Brennecke, J.; Cohen, S.M. Temporal reciprocity of miRNAs and their targets during the maternal-to-zygotic transition in Drosophila. Curr. Biol. 2008, 18, 501–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, E.; Liu, M.; Hartley, R.S.; Sheets, M.D.; Dahlberg, J.E. Deadenylation of maternal mRNAs mediated by miR-427 in Xenopus laevis embryos. RNA 2009, 15, 2351–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laue, K.; Rajshekar, S.; Courtney, A.J.; Lewis, Z.A.; Goll, M.G. The maternal to zygotic transition regulates genome-wide heterochromatin establishment in the zebrafish embryo. Nat. Commun. 2019, 10, 1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazzini, A.A.; del Viso, F.; Moreno-Mateos, M.A.; Johnstone, T.G.; Vejnar, C.E.; Qin, Y.D.; Yao, J.; Khokha, M.K.; Giraldez, A.J. Codon identity regulates mRNA stability and translation efficiency during the maternal-to-zygotic transition. EMBO J. 2016, 35, 2087–2103. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Coller, J. A Universal Code for mRNA Stability? Trends Genet. 2016, 32, 687–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishima, Y.; Tomari, Y. Codon Usage and 30 UTR Length Determine Maternal mRNA Stability in Zebrafish. Mol. Cell 2016, 61, 874–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, B.A.; O’Farrell, P.H. The three postblastoderm cell cycles of Drosophila embryogenesis are regulated in G2 by string. Cell 1990, 62, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Edgar, B.A.; Datar, S.A. Zygotic degradation of two maternal Cdc25 mRNAs terminates Drosophila’s early cell cycle program. Genes Dev. 1996, 10, 1966–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, B.; He, C.H.; Zhang, F.; Votruba, S.M.; Tadros, W.; Westwood, J.T.; Smibert, C.A.; Lipshitz, H.D.; Theurkauf, W.E. An essential role for the RNA-binding protein Smaug during the Drosophila maternal-to-zygotic transition. Development 2009, 136, 923–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.M.; Li, J.M.; Elemento, O.; Tavazoie, S.; Wieschaus, E.F. Coupling of zygotic transcription to mitotic control at the Drosophila mid-blastula transition. Development 2009, 136, 2101–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.; Baldwin, D.A.; Schultz, R.M. Transcript profiling during preimplantation mouse development. Dev. Biol. 2004, 272, 483–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Dumelie, J.G.; Li, X.; Cheng, M.H.; Yang, Z.; Laver, J.D.; Siddiqui, N.U.; Westwood, J.T.; Morris, Q.; Lipshitz, H.D.; et al. Global regulation of mRNA translation and stability in the early Drosophila embryo by the Smaug RNA-binding protein. Genome Biol. 2014, 15, R4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, H.; Yoshitake, N. Developmental stages of ovarian follicles of the silkworm, Bombyx mori L. J. Morphol. 1984, 179, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Nohata, J.; Guo, H.; Li, S.; Liu, J.; Guo, Y.; Yamamoto, K.; Kadono-Okuda, K.; Liu, C.; Arunkumar, K.P.; et al. A comprehensive analysis of the chorion locus in silkmoth. Sci. Rep. 2015, 5, 16424. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.W. The transformation of the Synaptonemal Complex into the ‘elimination chromatin’ in Bombyx mori oocytes. Chromosoma 1977, 60, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, N. The early embryonic mitosis in normal and cooled eggs of the silkworm, Bombyx mori. J. Morphol. 1978, 158, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, N. Fertilization and the first cleavage mitosis in insects. Dev. Growth Differ. 2001, 43, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.R.; Qin, S.; Xu, P.Z.; Zhang, G.Z. Identifying potential maternal genes of Bombyx mori using digital gene expression profiling. PLoS ONE 2018, 13, e0192745. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, M.; Jouraku, A.; Toyoda, A.; Yokoi, K.; Minakuchi, Y.; Katsuma, S.; Fujiyama, A.; Kiuchi, T.; Yamamoto, K.; Shimada, T. High-quality genome assembly of the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2019, 107, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Cheng, D.; Duan, J.; Wang, G.; Cheng, T.; Zha, X.; Liu, C.; Zhao, P.; Dai, F.; Zhang, Z.; et al. Microarray-based gene expression profiles in multiple tissues of the domesticated silkworm, Bombyx mori. Genome Biol. 2007, 8, R162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Dong, Z.; Duan, J.; Wang, G.; Wang, L.; Li, Y.; Xiang, Z.; Xia, Q. Genome-wide identification and immune response analysis of serine protease inhibitor genes in the silkworm, Bombyx mori. PLoS ONE 2012, 7, e31168. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Goldsmith, M.R.; Hunsley, J.R.; Kafatos, F.C. Specific protein synthesis in cellular differentiation. Production of eggshell proteins by silkmoth follicular cells. J. Cell Biol. 1972, 55, 653–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blau, H.M.; Kafatos, F.C. Secretory kinetics in the follicular cells of silkmoths during eggshell formation. J. Cell Biol. 1978, 78, 131–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadel, M.R.; Goldsmith, M.R.; Goplerud, J.; Kafatos, F.C. Specific protein synthesis in cellular differentiation. V. A secretory defect of chorion formation in the Grcol mutant of Bombyx mori. Dev. Biol. 1980, 75, 41–58. [Google Scholar] [CrossRef]
- Machado, E.; Swevers, L.; Sdralia, N.; Medeiros, M.N.; Mello, F.G.; Iatrou, K. Prostaglandin signaling and ovarian follicle development in the silkmoth, Bombyx mori. Insect Biochem. Mol. Biol. 2007, 37, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Nadel, M.R.; Kafatos, F.C. Specific protein synthesis in cellular differentiation. IV. The chorion proteins of Bombyx mori and their program of synthesis. Dev. Biol. 1980, 75, 26–40. [Google Scholar] [CrossRef]
- Swevers, L.; Iatrou, K. Early establishment and autonomous implementation of a developmental program controlling silkmoth chorion gene expression. Dev. Biol. 1992, 150, 12–22. [Google Scholar] [CrossRef]
- Surdej, P.; Jacobs-Lorena, M. Developmental regulation of bicoid mRNA stability is mediated by the first 43 nucleotides of the 3’ untranslated region. Mol. Cell Biol. 1998, 18, 2892–2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashirullah, A.; Cooperstock, R.L.; Lipshitz, H.D. Spatial and temporal control of RNA stability. Proc. Natl. Acad. Sci. USA 2001, 98, 7025–7028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadros, W.; Houston, S.A.; Bashirullah, A.; Cooperstock, R.L.; Semotok, J.L.; Reed, B.H.; Lipshitz, H.D. Regulation of maternal transcript destabilization during egg activation in Drosophila. Genetics 2003, 164, 989–1001. [Google Scholar] [PubMed]
- Tadros, W.; Lipshitz, H.D. Setting the stage for development: mRNA translation and stability during oocyte maturation and egg activation in Drosophila. Dev. Dyn. 2005, 232, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Rauwerda, H.; Pagano, J.F.; de Leeuw, W.C.; Ensink, W.; Nehrdich, U.; de Jong, M.; Jonker, M.; Spaink, H.P.; Breit, T.M. Transcriptome dynamics in early zebrafish embryogenesis determined by high-resolution time course analysis of 180 successive, individual zebrafish embryos. BMC Genom. 2017, 18, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathavan, S.; Lee, S.G.; Mak, A.; Miller, L.D.; Murthy, K.R.; Govindarajan, K.R.; Tong, Y.; Wu, Y.L.; Lam, S.H.; Yang, H.; et al. Transcriptome analysis of zebrafish embryogenesis using microarrays. PLoS Genet. 2005, 1, 260–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audic, Y.; Omilli, F.; Osborne, H.B. Postfertilization deadenylation of mRNAs in Xenopus laevis embryos is sufficient to cause their degradation at the blastula stage. Mol. Cell Biol. 1997, 17, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voeltz, G.K.; Steitz, J.A. AUUUA sequences direct mRNA deadenylation uncoupled from decay during Xenopus early development. Mol. Cell Biol. 1998, 18, 7537–7545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamatani, T.; Carter, M.G.; Sharov, A.A.; Ko, M.S. Dynamics of global gene expression changes during mouse preimplantation development. Dev. Cell 2004, 6, 117–131. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Tatsuke, T.; Mon, H.; Li, Z.; Xu, J.; Lee, J.M.; Kusakabe, T. Characterization of Tudor-sn-containing granules in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2013, 43, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Kirino, Y.; Maragkakis, M.; Alexiou, P.; Ohtaki, A.; Murali, R.; Mourelatos, Z.; Kirino, Y. Mitochondrial protein BmPAPI modulates the length of mature piRNAs. Rna Publ. Rna Soc. 2013, 19, 1405–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashiki, N.; Kawamura, N. Behaviors of nucleus, basal bodies and microtubules during eupyrene and apyrene spermiogenesis in the silkworm, Bombyx mori (Lepidoptera). Dev. Growth Differ. 1997, 39, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cheng, T.; Hu, W.; Peng, Z.; Liu, C.; Xia, Q. Genome-wide identification and analysis of JHBP-domain family members in the silkworm Bombyx mori. Mol. Genet. Genom. 2016, 291, 2159–2171. [Google Scholar] [CrossRef] [PubMed]
- Golub, T.R.; Slonim, D.K.; Tamayo, P.; Huard, C.; Gaasenbeek, M.; Mesirov, J.P.; Coller, H.; Loh, M.L.; Downing, J.R.; Caligiuri, M.A.; et al. Molecular classification of cancer: Class discovery and class prediction by gene expression monitoring. Science 1999, 286, 531–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Probe | Description | Protein Length (Amino Acids) | Location (Chr.) | Signal Peptide | NCBI Reference Sequence |
|---|---|---|---|---|---|---|---|
| me31B | BGIBMGA010673 | sw14777 | ATP-dependent RNA helicase me31b | 440 | 12 | – | AK383517 |
| lok | BGIBMGA005370 | sw11876 | cell cycle checkpoint kinase 2 | 528 | 8 | – | AK382539 |
| vri | BGIBMGA013421 | sw05474 | nuclear factor interleukin-3-regulated protein | 364 | 27 | – | AK388388 |
| Egfr | BGIBMGA000602 | sw11771 | epidermal growth factor receptor | 1449 | 1 | 1 | XM_004929742 |
| Su (var) 205 | BGIBMGA006109 | sw05786 | chromobox-like protein 5 | 191 | 4 | – | AK385880 |
| Hp1b-l | BGIBMGA012860 | sw21166 | heterochromatin protein | 179 | 16 | – | XM_012692600 |
| spz | BGIBMGA002397 | sw08256 | spatzle | 277 | 9 | 1 | NM_001114594 |
| tkv | BGIBMGA007355 | sw21102 | bone morphogenetic protein receptor type-1B | 485 | 3 | – | AK385287 |
| CycB | BGIBMGA003747 | sw04030 | cyclin B homolog | 525 | 5 | 1 | AK382330 |
| proPPAE | BGIBMGA013746 | sw20327 | prophenoloxidase activating enzyme precursor | 441 | 28 | 1 | AK383056 |
| asp | BGIBMGA005594 | sw10953 | protein abnormal spindle | 2309 | 17 | – | XM_004921876 |
| PAH | BGIBMGA003866 | sw01062 | phenylalanine hydroxylase | 456 | 1 | – | NM_001287837 |
| aub | BGIBMGA010644 | sw10899 | aubergine protein | 899 | 12 | – | EU143547 |
| Csp (DnaJ-7) | BGIBMGA007808 | sw01087 | dnaJ (Hsp40) homolog 7 | 203 | 15 | – | XM_012692267 |
| SPE | BGIBMGA005172 | – | serine protease easter | 430 | 25 | 1 | XM_012689474 |
| BAEE | BGIBMGA005173 | – | BzArgOEtase | 369 | 25 | 1 | NM_001043379 |
| PPAE | BGIBMGA010546 | sw15390 | prophenoloxidase activating enzyme | 382 | 12 | 1 | AK383498 |
| Sod2 | BGIBMGA007453 | sw21871 | Mn superoxide dismutase | 221 | 3 | – | XM_012690443 |
| esc | BGIBMGA006325 | sw12637 | extra sex combs | 411 | 6 | – | AK385410 |
| Src42A | BGIBMGA004089 | sw11866 | tyrosine-protein kinase Src42A-like | 922 | 19 | – | XM_012693691 |
| Smg | BGIBMGA008249 | sw17394 | Bombyx mori protein Smaug | 599 | 18 | – | AK385418 |
| Eif-4a | BGIBMGA003186 | sw22934 | eukaryotic translation initiation factor 4A | 420 | 4 | – | AK383662 |
| eIF4AIII | BGIBMGA004822 | sw12663 | eukaryotic initiation factor 4A-III | 405 | 25 | – | AK386335 |
| rod | BGIBMGA002655 | sw06197 | rough deal protein | 1817 | 28 | – | XM_004932260 |
| vfl | BGIBMGA012283 | sw05259 | zinc finger protein | 1064 | 1 | – | XM_004933146 |
| bai | BGIBMGA004891 | sw00285 | transmembrane trafficking protein | 205 | 25 | – | AK385774 |
| Nelf-E | BGIBMGA003207 | sw19434 | negative elongation factor E | 264 | 13 | – | AK385219 |
| Pabn2 | BGIBMGA001950 | – | polyadenylate binding protein 2 | 225 | 19 | – | XM_012696483 |
| Bin1 | BGIBMGA011014 | sw11118 | histone deacetylase complex subunit SAP18 | 159 | 23 | – | AK384481 |
| tud | BGIBMGA011857 | sw17672 | maternal protein tudor | 1839 | 11 | – | XM_012695006 |
| Moe | BGIBMGA002544 | sw02031 | moesin/ezrin/radixin homolog 1 | 574 | 9 | – | AK383231 |
| Sel (cnpy1) | BGIBMGA003267 | sw18745 | protein canopy homolog 1 | 242 | 2 | – | AK385660 |
| Hip14 (ZDHHC17) | BGIBMGA001083 | sw14724 | palmitoyltransferase ZDHHC17 | 591 | 13 | – | XM_004927675 |
| mamo | BGIBMGA012517 | sw18043 | zinc finger protein | 798 | 9 | – | XM_012688563 |
| sax | BGIBMGA009134 | sw06935 | activin receptor type-1 | 566 | 20 | 1 | XM_004925975 |
| babo | BGIBMGA000601 | sw20139 | TGF-beta receptor type-1 | 503 | 1 | – | XM_012693543 |
| h | BGIBMGA005390 | sw08720 | protein hairy isoform | 261 | 8 | – | XM_004932202 |
| Chc | BGIBMGA012935 | sw07960 | Bombyx mori clathrin heavy chain | 1681 | 16 | – | AK378376 |
| Snap25 | BGIBMGA005176 | sw12219 | synaptosomal-associated protein 25 | 211 | 25 | – | AK383225 |
| SPE-like | BGIBMGA013797 | sw18366 | serine protease easter-like | 431 | 28 | 1 | AK386026 |
| Src64B | BGIBMGA012094 | sw07585 | tyrosine-protein kinase Src64B | 521 | 11 | – | AK378283 |
| wbl | BGIBMGA012931 | sw05230 | Bombyx mori protein windbeutel | 254 | 16 | 1 | AK381984 |
| Mat89Ba | BGIBMGA007162 | sw18950 | nucleolar protein 6 | 1120 | 21 | – | AK385389 |
| Dif | BGIBMGA010496 | sw17578 | embryonic polarity protein dorsal isoform | 529 | 12 | – | AK386522 |
| ndl (osp) | BGIBMGA014089 | sw15075 | ovarian serine protease | 1920 | 9 | – | XM_012691651 |
| Nelf-A | BGIBMGA002236 | sw03405 | negative elongation factor A | 581 | 26 | – | XM_012691503 |
| tld | BGIBMGA002518 | sw11946 | tolloid-like protein 1 | 1349 | 9 | 1 | XM_012694771 |
| proSP7 | BGIBMGA012427 | sw09635 | serine protease 7 precursor | 397 | 21 | 1 | AK386200 |
| gammaTub | BGIBMGA013500 | sw02138 | tubulin gamma-1 | 456 | 15 | – | AK377270 |
| Th | BGIBMGA000563 | sw13482 | tyrosine hydroxylase | 561 | 1 | – | AK383721 |
| pie | BGIBMGA001789 | sw19480 | G2/M phase-specific E3 ubiquitin-protein ligase | 757 | 11 | – | XM_004922174 |
| gro | BGIBMGA012449 | sw19514 | groucho-like isoform X1 | 679 | 21 | – | AK382427 |
| hb | BGIBMGA003334 | sw12894 | protein hunchback | 621 | 15 | – | AK385224 |
| pip | BGIBMGA011817 | sw14126 | heparan sulfate 2-O-sulfotransferase pipe | 436 | 11 | – | XM_004931477 |
| spoon (AKAP1) | BGIBMGA006841 | sw04955 | A-kinase anchor protein 1 | 3601 | 10 | – | XM_004924760 |
| snk | BGIBMGA001745 | sw01630 | venom protease-like | 401 | 11 | 1 | XM_004922131 |
| Btk29A | BGIBMGA000972 | sw08339 | tyrosine-protein kinase Btk29A | 610 | 13 | – | XM_012691697 |
| dpp | BGIBMGA010384 | sw00355 | decapentaplegic | 369 | 12 | 1 | XM_012693077 |
| Msp300 (nesprin-1) | BGIBMGA010471 | sw17350 | nesprin-1 | 8514 | 12 | – | XM_012693124 |
| KCNQ | BGIBMGA003731 | sw13728 | potassium voltage-gated channel subfamily KQT member 5 | 751 | 9 | – | XM_012693718 |
| shot | BGIBMGA004414 | sw05774 | Bombyx mori plectin-like | 1325 | 20 | – | XM_012695224 |
| sog | BGIBMGA005348 | sw05769 | dorsal-ventral patterning protein Sog | 927 | 8 | 1 | XM_012695533 |
| Pc | BGIBMGA006904 | sw14515 | polycomb | 281 | 10 | – | AK383962 |
| Dst | BGIBMGA004415 | sw01256 | Bombyx mori dystonin-like | 4811 | 20 | – | XM_012695102 |
| TPH1 | BGIBMGA000642 | sw03004 | tryptophan 5-hydroxylase 1 | 543 | 1 | – | NM_001309589 |
| glo (hnRNPF) | BGIBMGA013473 | sw04558 | heterogeneous nuclear ribonucleoprotein F | 336 | 6 | – | XM_012690736 |
| Cluster | No. of Maternal Genes | Name of Maternal Genes |
|---|---|---|
| 1 | 8 | Sod2, Pabn2, Eif-4a, Bin1, eIF4AIII, Chc, bai, tud |
| 2 | 17 | gammaTub, Nelf-E, lok, Mat89Ba, PPAE, Pc, proPPAE, tld, esc, Btk29A, Src64B, shot, wbl, Smg, spoon(AKAP1), Msp300(nesprin-1), TPH1 |
| 3 | 41 | sog, rod, me31B, vfl, vri, KCNQ, Egfr, Sel(cnpy1), Su(var)205, Hip14(ZDHHC17), Hp1b-l, Nelf-A, spz, mamo, tkv, hb, CycB, babo, dpp, snk, asp, proSP7, PAH, h, aub, glo(hnRNPF), Csp(DnaJ-7), pie, SPE, sax, BAEE, gro, SPE-like, Dif, ndl(osp), Th, Src42A, pip, Dst, Snap25, Moe |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Xu, P.; Pang, H.; Chen, T.; Zhang, G. Expression Analysis of mRNA Decay of Maternal Genes during Bombyx mori Maternal-to-Zygotic Transition. Int. J. Mol. Sci. 2019, 20, 5651. https://doi.org/10.3390/ijms20225651
Zhang M, Xu P, Pang H, Chen T, Zhang G. Expression Analysis of mRNA Decay of Maternal Genes during Bombyx mori Maternal-to-Zygotic Transition. International Journal of Molecular Sciences. 2019; 20(22):5651. https://doi.org/10.3390/ijms20225651
Chicago/Turabian StyleZhang, Meirong, Pingzhen Xu, Huilin Pang, Tao Chen, and Guozheng Zhang. 2019. "Expression Analysis of mRNA Decay of Maternal Genes during Bombyx mori Maternal-to-Zygotic Transition" International Journal of Molecular Sciences 20, no. 22: 5651. https://doi.org/10.3390/ijms20225651
APA StyleZhang, M., Xu, P., Pang, H., Chen, T., & Zhang, G. (2019). Expression Analysis of mRNA Decay of Maternal Genes during Bombyx mori Maternal-to-Zygotic Transition. International Journal of Molecular Sciences, 20(22), 5651. https://doi.org/10.3390/ijms20225651