Emetine Synergizes with Cisplatin to Enhance Anti-Cancer Efficacy against Lung Cancer Cells

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
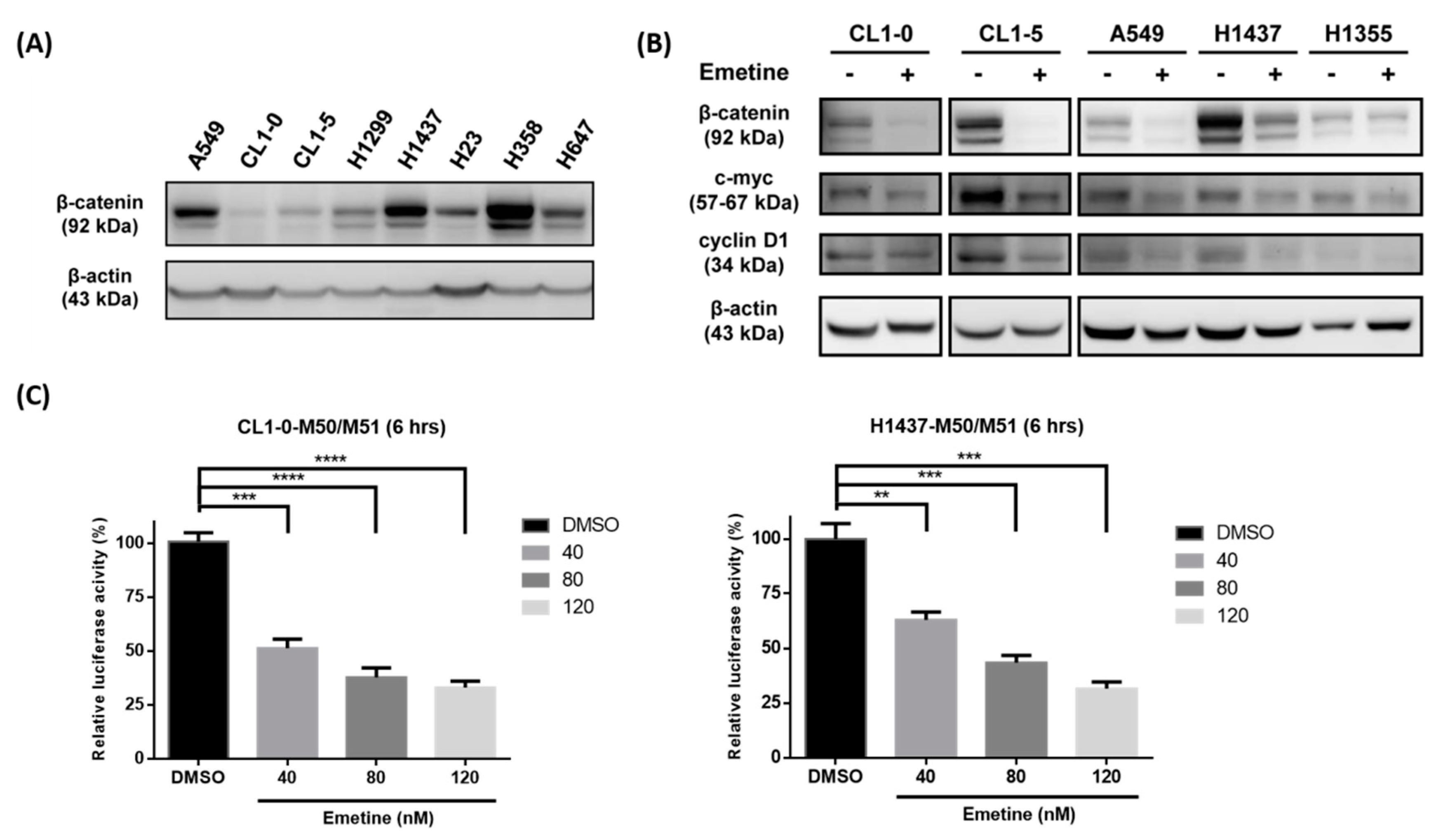
2.1. Emetine Inhibits the Wnt/β-catenin Pathway, c-myc and Cyclin D1 in Human NSCLC Cells
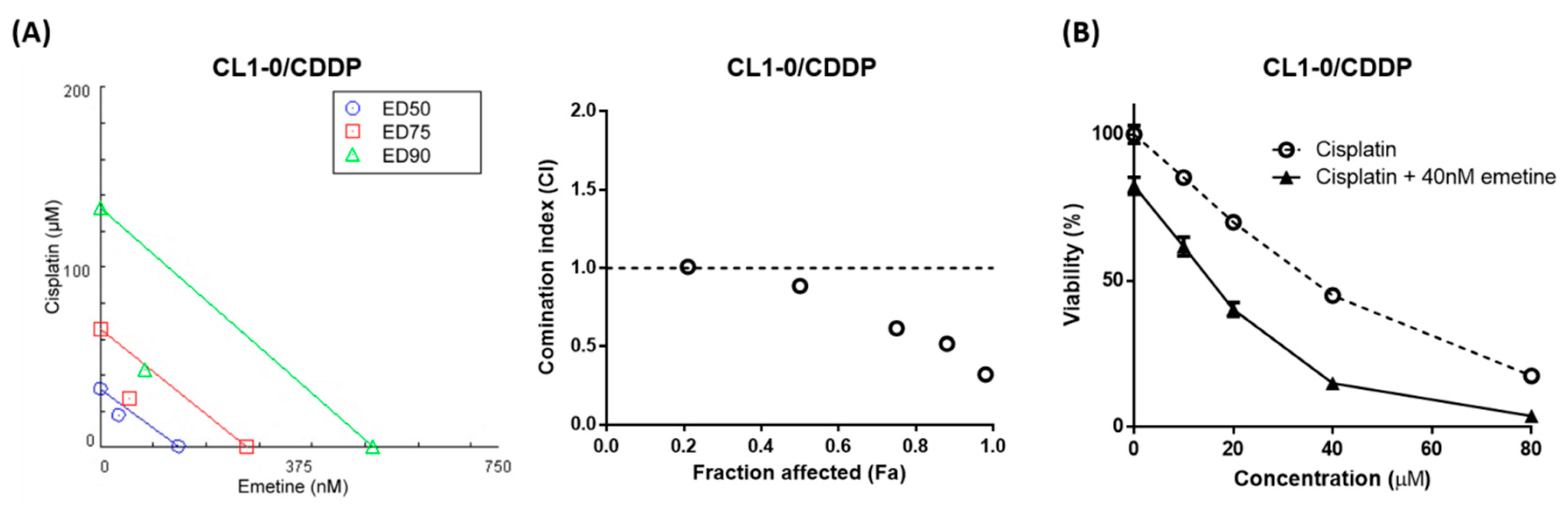
2.2. Emetine Synergizes with Cisplatin to Enhance its Anticancer Efficacy in Human NSCLC Cells
2.3. Low-Dose Emetine Sensitizes Lung Cancer Cells to Cisplatin
2.4. Nuclear β-catenin and Stemness-Related Markers are Upregulated in the Cisplatin-resistant Subpopulation of CL1-0 cells
2.5. Emetine Enhances The Anticancer Efficacy of Cisplatin and Synergizes with Cisplatin in the Cisplatin-Resistant Subpopulation of CL1-0 Cells
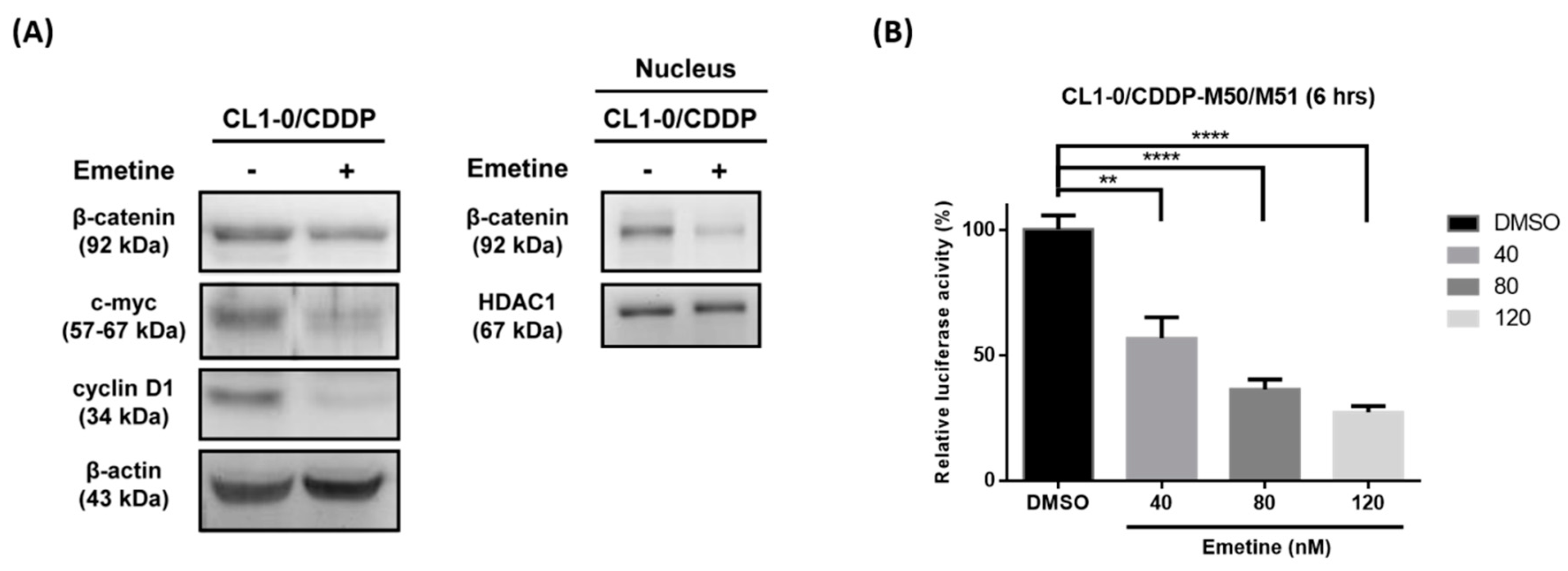
2.6. Emetine Inhibits the Wnt/β-catenin Pathway in the Cisplatin-Resistant Subpopulation of CL1-0 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Drug Treatment
4.2. Drugs
4.3. Establishment of the Cisplatin-Resistant Cell Subpopulations
4.4. Cell Viability Assay
4.5. Isobologram and the Combination Index (CI)
4.6. Western Blot Analysis
4.7. TCF/LEF Luciferase Assay
4.8. Spheroid Formation Ability
4.9. Immunocytochemical (ICC) Staining
4.10. Nuclear Extraction
4.11. RNA Extraction and Real-Time Quantitative Polymerase Chain Reaction
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Tieche, C.C.; Peng, R.W.; Dorn, P.; Froment, L.; Schmid, R.A.; Marti, T.M. Prolonged pemetrexed pretreatment augments persistence of cisplatin-induced DNA damage and eliminates resistant lung cancer stem-like cells associated with EMT. BMC Cancer 2016, 16, 125. [Google Scholar] [CrossRef] [PubMed]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar] [CrossRef] [PubMed]
- Giaccone, G.; Splinter, T.A.; Debruyne, C.; Kho, G.S.; Lianes, P.; van Zandwijk, N.; Pennucci, M.C.; Scagliotti, G.; van Meerbeeck, J.; van Hoesel, Q.; et al. Randomized study of paclitaxel-cisplatin versus cisplatin-teniposide in patients with advanced non-small-cell lung cancer. The European Organization for Research and Treatment of Cancer Lung Cancer Cooperative Group. J. Clin. Oncol. 1998, 16, 2133–2141. [Google Scholar] [CrossRef]
- Schiller, J.H.; Harrington, D.; Belani, C.P.; Langer, C.; Sandler, A.; Krook, J.; Zhu, J.; Johnson, D.H. Comparison of four chemotherapy regimens for advanced non-small-cell lung cancer. N. Engl. J. Med. 2002, 346, 92–98. [Google Scholar] [CrossRef]
- Gao, J.; Meng, Q.; Zhao, Y.; Chen, X.; Cai, L. EHD1 confers resistance to cisplatin in non-small cell lung cancer by regulating intracellular cisplatin concentrations. BMC Cancer 2016, 16, 470. [Google Scholar] [CrossRef]
- Peng, Y.Y.; He, Y.H.; Chen, C.; Xu, T.; Li, L.; Ni, M.M.; Meng, X.M.; Huang, C.; Li, J. NLRC5 regulates cell proliferation, migration and invasion in hepatocellular carcinoma by targeting the Wnt/beta-catenin signaling pathway. Cancer Lett. 2016, 376, 10–21. [Google Scholar] [CrossRef]
- Hubaux, R.; Thu, K.L.; Lam, W.L. Re: The Wnt signaling pathway in non-small cell lung cancer. J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef]
- Chen, X.; Song, X.; Yue, W.; Chen, D.; Yu, J.; Yao, Z.; Zhang, L. Fibulin-5 inhibits Wnt/beta-catenin signaling in lung cancer. Oncotarget 2015, 6, 15022–15034. [Google Scholar] [CrossRef]
- Fang, L.; Cai, J.; Chen, B.; Wu, S.; Li, R.; Xu, X.; Yang, Y.; Guan, H.; Zhu, X.; Zhang, L.; et al. Aberrantly expressed miR-582-3p maintains lung cancer stem cell-like traits by activating Wnt/beta-catenin signalling. Nat. Commun. 2015, 6, 8640. [Google Scholar] [CrossRef] [PubMed]
- Akiri, G.; Cherian, M.M.; Vijayakumar, S.; Liu, G.; Bafico, A.; Aaronson, S.A. Wnt pathway aberrations including autocrine Wnt activation occur at high frequency in human non-small-cell lung carcinoma. Oncogene 2009, 28, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Jiang, Y.; Yang, X.; Wang, S.; Xie, C.; Li, X.; Li, Y.; Chen, Y.; Wang, X.; Meng, Y.; et al. Wnt/beta-catenin pathway mediates (−)−Epigallocatechin-3-gallate (EGCG) inhibition of lung cancer stem cells. Biochem. Biophys. Res. Commun. 2017, 482, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Shih, Y.L.; Lin, S.P.; Hsieh, T.Y.; Lin, Y.W. YC-1 Antagonizes Wnt/beta-Catenin Signaling Through the EBP1 p42 Isoform in Hepatocellular Carcinoma. Cancers (Basel) 2019, 11, 661. [Google Scholar] [CrossRef]
- Akinboye, E.; Bakare, O. Biological activities of emetine. Open Nat. Prod. J. 2011, 4, 8–15. [Google Scholar] [CrossRef]
- Panettiere, F.; Coltman, C.A., Jr. Experience with emetine hydrochloride (NSC 33669) as an antitumor agent. Cancer 1971, 27, 835–841. [Google Scholar] [CrossRef]
- Moller, M.; Herzer, K.; Wenger, T.; Herr, I.; Wink, M. The alkaloid emetine as a promising agent for the induction and enhancement of drug-induced apoptosis in leukemia cells. Oncol. Rep. 2007, 18, 737–744. [Google Scholar] [CrossRef]
- Moller, M.; Wink, M. Characteristics of apoptosis induction by the alkaloid emetine in human tumour cell lines. Planta Med. 2007, 73, 1389–1396. [Google Scholar] [CrossRef]
- Sun, Q.; Yogosawa, S.; Iizumi, Y.; Sakai, T.; Sowa, Y. The alkaloid emetine sensitizes ovarian carcinoma cells to cisplatin through downregulation of bcl-xL. Int. J. Oncol. 2015, 46, 389–394. [Google Scholar] [CrossRef]
- Davidson, V.J.; Patel, D.; Flanigan, R.; Gupta, G.N.; Foreman, K.E. Emetine reduces the effective dose of cisplatin or carboplatin required to inhibit bladder cancer cell proliferation. Bladder 2017, 4. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, E.B.; Lee, J.; Jung, O.; Ryu, B.J.; Kim, S.H.; Cho, J.Y.; Ryou, C.; Lee, S.Y. Emetine inhibits migration and invasion of human non-small-cell lung cancer cells via regulation of ERK and p38 signaling pathways. Chem. Biol. Interact. 2015, 242, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, M.J.; Grage, T.B.; Bellet, R.E.; Weiss, A.J. A phase I study of emetine hydrochloride (NSC 33669) in solid tumors. Cancer 1973, 31, 1170–1175. [Google Scholar] [CrossRef]
- Siddiqui, S.; Firat, D.; Olshin, S. Phase II study of emetine (NSC-33669) in the treatment of solid tumors. Cancer Chemother. Rep. 1973, 57, 423–428. [Google Scholar] [PubMed]
- Kane, R.C.; Cohen, M.H.; Broder, L.E.; Bull, M.I.; Creaven, P.J.; Fossieck, B.E., Jr. Phase I-II evaluation of emetine (NSC-33669) in the treatment of epidermoid bronchogenic carcinoma. Cancer Chemother. Rep. 1975, 59, 1171–1172. [Google Scholar]
- Hopf, N.J.; Goebel, H.H. Experimental emetine myopathy: Enzyme histochemical, electron microscopic, and immunomorphological studies. Acta Neuropathol. 1993, 85, 414–418. [Google Scholar] [CrossRef]
- Pan, S.J.; Combs, A.B. Effects of pharmacological interventions on emetine cardiotoxicity in isolated perfused rat hearts. Toxicology 1995, 97, 93–104. [Google Scholar] [CrossRef]
- Pan, S.J.; Combs, A.B. Emetine inhibits glycolysis in isolated, perfused rat hearts. Cardiovasc. Toxicol. 2003, 3, 311–318. [Google Scholar] [CrossRef]
- Barr, M.P.; Gray, S.G.; Hoffmann, A.C.; Hilger, R.A.; Thomale, J.; O’Flaherty, J.D.; Fennell, D.A.; Richard, D.; O’Leary, J.J.; O’Byrne, K.J. Generation and characterisation of cisplatin-resistant non-small cell lung cancer cell lines displaying a stem-like signature. PLoS ONE 2013, 8, e54193. [Google Scholar] [CrossRef]
- Chau, N.M.; Rogers, P.; Aherne, W.; Carroll, V.; Collins, I.; McDonald, E.; Workman, P.; Ashcroft, M. Identification of novel small molecule inhibitors of hypoxia-inducible factor-1 that differentially block hypoxia-inducible factor-1 activity and hypoxia-inducible factor-1alpha induction in response to hypoxic stress and growth factors. Cancer Res. 2005, 65, 4918–4928. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-D.; Kim, Y.-P.; Mohammed, K.A.; Jones, D.K.; Muhammad, I.; Dunbar, D.C.; Nagle, D.G. Terpenoid Tetrahydroisoquinoline Alkaloids Emetine, Klugine, and Isocephaeline Inhibit the Activation of Hypoxia-Inducible Factor-1 in Breast Tumor Cells. J. Nat. Prod. 2005, 68, 947–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardizzoni, A.; Group, F.T.C.M.-A.; Boni, L.; Fossella, F.V.; Schiller, J.H. Cisplatin- Versus Carboplatin-Based Chemotherapy in First-Line Treatment of Advanced Non–Small-Cell Lung Cancer: An Individual Patient Data Meta-analysis. J. Natl. Cancer Inst. 2007, 99, 847–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.S. Chemotherapy Resistance in Lung Cancer. Adv. Exp. Med. Biol. 2016, 893, 189–209. [Google Scholar] [CrossRef]
- Ashburn, T.T.; Thor, K.B. Drug repositioning: Identifying and developing new uses for existing drugs. Nat. Rev. Drug Discov. 2004, 3, 673–683. [Google Scholar] [CrossRef]
- Chong, C.R.; Sullivan, D.J., Jr. New uses for old drugs. Nature 2007, 448, 645–646. [Google Scholar] [CrossRef]
- Fong, W.; To, K.K.W. Drug repurposing to overcome resistance to various therapies for colorectal cancer. Cell. Mol. Life Sci. 2019. [Google Scholar] [CrossRef]
- Stewart, D.J. Wnt signaling pathway in non-small cell lung cancer. J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef]
- Jin, J.; Zhan, P.; Qian, H.; Wang, X.; Katoh, M.; Phan, K.; Chung, J.H.; Lv, T.; Song, Y. Prognostic value of wingless-type proteins in non-small cell lung cancer patients: A meta-analysis. Transl. Lung Cancer Res. 2016, 5, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, T.; Liu, D.; Nakano, J.; Ishikawa, S.; Yokomise, H.; Ueno, M.; Kadota, K.; Huang, C.L. Wnt1 overexpression associated with tumor proliferation and a poor prognosis in non-small cell lung cancer patients. Oncol. Rep. 2008, 19, 203–209. [Google Scholar] [CrossRef]
- Wang, Q.; Zhan, P.; Yu, L.; Song, Y. The correlation between the expression of Wnt1 and prognosis in resected non-small cell lung cancer. Zhongguo Fei Ai Za Zhi 2010, 13, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sun, P.L.; Li, J.Z.; Jheon, S.; Lee, C.T.; Chung, J.H. Aberrant Wnt1/beta-catenin expression is an independent poor prognostic marker of non-small cell lung cancer after surgery. J. Thorac. Oncol. 2011, 6, 716–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, M.; Akiri, G.; Chin, C.; Wisnivesky, J.P.; Beasley, M.B.; Weiser, T.S.; Swanson, S.J.; Aaronson, S.A. Wnt pathway activation predicts increased risk of tumor recurrence in patients with stage I nonsmall cell lung cancer. Ann. Surg. 2013, 257, 548–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.X.; Chiang, A.C.; Zhang, X.H.; Kim, J.Y.; Kris, M.G.; Ladanyi, M.; Gerald, W.L.; Massague, J. WNT/TCF signaling through LEF1 and HOXB9 mediates lung adenocarcinoma metastasis. Cell 2009, 138, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coscio, A.; Chang, D.W.; Roth, J.A.; Ye, Y.; Gu, J.; Yang, P.; Wu, X. Genetic variants of the Wnt signaling pathway as predictors of recurrence and survival in early-stage non-small cell lung cancer patients. Carcinogenesis 2014, 35, 1284–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Yan, J.; Lang, X.; Zhuang, Y. Expression of circ_001569 is upregulated in osteosarcoma and promotes cell proliferation and cisplatin resistance by activating the Wnt/beta-catenin signaling pathway. Oncol. Lett. 2018, 16, 5856–5862. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Q.; Chen, H.; Wang, C. BCL9 promotes epithelial mesenchymal transition and invasion in cisplatin resistant NSCLC cells via beta-catenin pathway. Life Sci. 2018, 208, 284–294. [Google Scholar] [CrossRef]
- He, F.; Wang, H.; Li, Y.; Liu, W.; Gao, X.; Chen, D.; Wang, Q.; Shi, G. SRPX2 knockdown inhibits cell proliferation and metastasis and promotes chemosensitivity in esophageal squamous cell carcinoma. Biomed. Pharmacother. 2019, 109, 671–678. [Google Scholar] [CrossRef]
- Jiang, H.; Xiong, W.; Chen, L.; Lv, Z.; Yang, C.; Li, Y. Knockdown of the long noncoding RNA HOTTIP inhibits cell proliferation and enhances cell sensitivity to cisplatin by suppressing the Wnt/beta-catenin pathway in prostate cancer. J. Cell. Biochem. 2019, 120, 8965–8974. [Google Scholar] [CrossRef]
- Mann, M.; Kumar, S.; Sharma, A.; Chauhan, S.S.; Bhatla, N.; Kumar, S.; Bakhshi, S.; Gupta, R.; Kumar, L. PARP-1 inhibitor modulate beta-catenin signaling to enhance cisplatin sensitivity in cancer cervix. Oncotarget 2019, 10, 4262–4275. [Google Scholar] [CrossRef]
- Milosevic, V.; Kopecka, J.; Salaroglio, I.C.; Libener, R.; Napoli, F.; Izzo, S.; Orecchia, S.; Ananthanarayanan, P.; Bironzo, P.; Grosso, F.; et al. Wnt/IL-1beta/IL-8 autocrine circuitries control chemoresistance in mesothelioma initiating cells by inducing ABCB5. Int. J. Cancer 2019. [Google Scholar] [CrossRef]
- Ruan, X.; Liu, A.; Zhong, M.; Wei, J.; Zhang, W.; Rong, Y.; Liu, W.; Li, M.; Qing, X.; Chen, G.; et al. Silencing LGR6 Attenuates Stemness and Chemoresistance via Inhibiting Wnt/beta-Catenin Signaling in Ovarian Cancer. Mol. Ther. Oncolytics 2019, 14, 94–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Xi, J.; Xu, X.; Peng, B.; Zhang, B. MiR-148a suppressed cell invasion and migration via targeting WNT10b and modulating beta-catenin signaling in cisplatin-resistant colorectal cancer cells. Biomed. Pharmacother. 2019, 109, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.L.; Zhang, Z.H. Long non-coding RNA SNHG1 contributes to cisplatin resistance in non-small cell lung cancer by regulating miR-140-5p/Wnt/beta-catenin pathway. Neoplasma 2019. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, J.; Gao, P.; Su, K.; Wu, H.; Li, J.; Lou, W. Wnt1-inducible signaling protein 1 regulates laryngeal squamous cell carcinoma glycolysis and chemoresistance via the YAP1/TEAD1/GLUT1 pathway. J. Cell. Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yang, Y.; Xian, Y.S. HCRP1 inhibits cell proliferation and invasion and promotes chemosensitivity in esophageal squamous cell carcinoma. Chem. Biol. Interact. 2019, 308, 357–363. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, K.; Wu, J.; Shi, J.; Xue, J.; Li, J.; Chen, J.; Zhu, Y.; Wei, J.; He, J.; et al. Wnt5a Increases Properties of Lung Cancer Stem Cells and Resistance to Cisplatin through Activation of Wnt5a/PKC Signaling Pathway. Stem Cells Int. 2016, 2016, 1690896. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Li, H.; Wang, J. beta-Catenin signaling pathway regulates cisplatin resistance in lung adenocarcinoma cells by upregulating Bcl-xl. Mol. Med. Rep. 2016, 13, 2543–2551. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhao, W.; Zhang, L.; Wang, L.; Wang, J.; Wan, Z.; Hong, Y.; Yu, L. MALAT1-miR-101-SOX9 feedback loop modulates the chemo-resistance of lung cancer cell to DDP via Wnt signaling pathway. Oncotarget 2017, 8, 94317–94329. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Shi, J.; Zhang, K.; Xue, J.; Li, J.; Yang, J.; Chen, J.; Wei, J.; Ren, H.; Liu, X. Sox2 inhibits Wnt-beta-catenin signaling and metastatic potency of cisplatin-resistant lung adenocarcinoma cells. Mol. Med. Rep. 2017, 15, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Xu, H.; Xu, C.; Chen, A.; Chen, L.; Zhou, M.; Haq, I.U.; Wu, X.; Mariyam, Z.; Feng, Q. NEAT1 contributes to the CSC-like traits of A549/CDDP cells via activating Wnt signaling pathway. Chem. Biol. Interact. 2018, 296, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Blakely, C.M.; Watkins, T.B.K.; Wu, W.; Gini, B.; Chabon, J.J.; McCoach, C.E.; McGranahan, N.; Wilson, G.A.; Birkbak, N.J.; Olivas, V.R.; et al. Evolution and clinical impact of co-occurring genetic alterations in advanced-stage EGFR-mutant lung cancers. Nat. Genet. 2017, 49, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Wang, X.; Wang, Y.; Ma, D. Wnt/beta-catenin signaling regulates cancer stem cells in lung cancer A549 cells. Biochem. Biophys. Res. Commun. 2010, 392, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Herlyn, M. The emerging roles of Oct4 in tumor-initiating cells. Am. J. Physiol. Cell. Physiol. 2015, 309, C709–C718. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Gao, Q.; Suo, Z.; Munthe, E.; Solberg, S.; Ma, L.; Wang, M.; Westerdaal, N.A.; Kvalheim, G.; Gaudernack, G. Identification and characterization of cells with cancer stem cell properties in human primary lung cancer cell lines. PLoS ONE 2013, 8, e57020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, G.; Kang, Y.; Dong, Z.; Qian, Q.; Ma, X. N-cadherin expression is associated with acquisition of EMT phenotype and with enhanced invasion in erlotinib-resistant lung cancer cell lines. PLoS ONE 2013, 8, e57692. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.-H.; Chang, S.-Y.; Shih, Y.-L.; Huang, T.-W.; Chang, H.; Lin, Y.-W. Emetine Synergizes with Cisplatin to Enhance Anti-Cancer Efficacy against Lung Cancer Cells. Int. J. Mol. Sci. 2019, 20, 5914. https://doi.org/10.3390/ijms20235914
Wu T-H, Chang S-Y, Shih Y-L, Huang T-W, Chang H, Lin Y-W. Emetine Synergizes with Cisplatin to Enhance Anti-Cancer Efficacy against Lung Cancer Cells. International Journal of Molecular Sciences. 2019; 20(23):5914. https://doi.org/10.3390/ijms20235914
Chicago/Turabian StyleWu, Ti-Hui, Shan-Yueh Chang, Yu-Lueng Shih, Tsai-Wang Huang, Hung Chang, and Ya-Wen Lin. 2019. "Emetine Synergizes with Cisplatin to Enhance Anti-Cancer Efficacy against Lung Cancer Cells" International Journal of Molecular Sciences 20, no. 23: 5914. https://doi.org/10.3390/ijms20235914
APA StyleWu, T. -H., Chang, S. -Y., Shih, Y. -L., Huang, T. -W., Chang, H., & Lin, Y. -W. (2019). Emetine Synergizes with Cisplatin to Enhance Anti-Cancer Efficacy against Lung Cancer Cells. International Journal of Molecular Sciences, 20(23), 5914. https://doi.org/10.3390/ijms20235914