Plasma Levels of snoRNAs are Associated with Platelet Activation in Patients with Peripheral Artery Disease


Abstract
:1. Introduction
2. Results
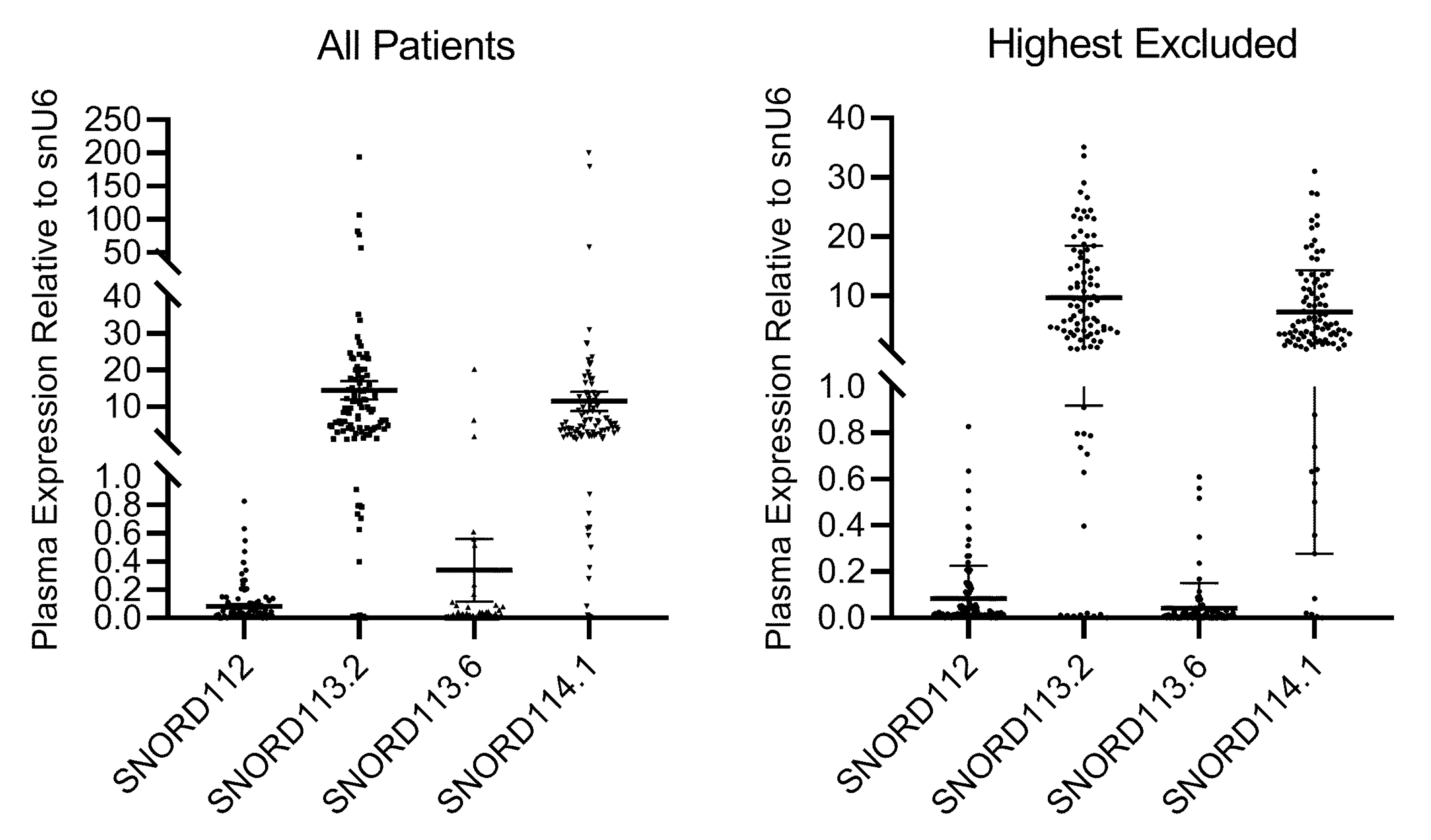
2.1. SnoRNA Plasma Levels
2.2. Clinical Endpoints
2.3. Classical Risk Factors
2.4. SnoRNA Levels and Smoking
2.5. SnoRNA levels and Platelet Function
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Blood Sampling
4.3. Determination of P-Selectin Expression and Glycoprotein (GP) IIb/IIIa Activation
4.4. Determination of Monocyte-Platelet Aggregates (MPA)
4.5. Vasodilator-Stimulated Phosphoprotein (VASP) Phosphorylation Assay
4.6. Clinical Endpoints
4.7. RNA Isolation and SnoRNA Measurements
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| RNA | ribonucleic acid |
| PAD | peripheral artery disease |
| snoRNA | small nucleolar RNA |
| MI | myocardial infarction |
| TIA | transient ischemic attack |
| STEMI | ST-elevated myocardial infarction |
| PCI | percutaneous coronary intervention |
| snRNAs | small nuclear RNA |
| rRNAs | ribosomal RNA |
| Lpa | lipoprotein a |
| WBC | white blood cell count |
| PRI | platelet reactivity index |
| MPA | monocyte-platelet aggregate |
| VASP | Vasodilator-stimulated phosphoprotein |
References
- Hiatt, W.R.; Goldstone, J.; Smith, S.C., Jr.; McDermott, M.; Moneta, G.; Oka, R.; Newman, A.B.; Pearce, W.H.; American Heart Association Writing Group 1. Atherosclerotic Peripheral Vascular Disease Symposium II: Nomenclature for vascular diseases. Circulation 2008, 118, 2826–2829. [Google Scholar] [CrossRef] [PubMed]
- Hiatt, W.R.; Armstrong, E.J.; Larson, C.J.; Brass, E.P. Pathogenesis of the limb manifestations and exercise limitations in peripheral artery disease. Circ. Res. 2015, 116, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Jukema, J.W.; Verschuren, J.J.; Ahmed, T.A.; Quax, P.H. Restenosis after PCI. Part 1: Pathophysiology and risk factors. Nat. Rev. Cardiol. 2011, 9, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Price, J.F.; Russell, M.J.; Smith, F.B.; van Wijk, M.C.; Fowkes, F.G. Improved prediction of fatal myocardial infarction using the ankle brachial index in addition to conventional risk factors: The Edinburgh Artery Study. Circulation 2004, 110, 3075–3080. [Google Scholar] [CrossRef] [PubMed]
- Diehm, C.; Lange, S.; Darius, H.; Pittrow, D.; von Stritzky, B.; Tepohl, G.; Haberl, R.L.; Allenberg, J.R.; Dasch, B.; Trampisch, H.J. Association of low ankle brachial index with high mortality in primary care. Eur. Heart J. 2006, 27, 1743–1749. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, S.; Jurisic, M.; Kopp, C.W.; Koppensteiner, R.; Huber, K.; Wojta, J.; Gremmel, T. Circulating microRNAs identify patients at increased risk of in-stent restenosis after peripheral angioplasty with stent implantation. Atherosclerosis 2018, 269, 197–203. [Google Scholar] [CrossRef]
- Stather, P.W.; Sylvius, N.; Wild, J.B.; Choke, E.; Sayers, R.D.; Bown, M.J. Differential microRNA expression profiles in peripheral arterial disease. Circ. Cardiovasc. Genet. 2013, 6, 490–497. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Hakansson, K.E.J.; Sollie, O.; Simons, K.H.; Quax, P.H.A.; Jensen, J.; Nossent, A.Y. Circulating Small Non-coding RNAs as Biomarkers for Recovery After Exhaustive or Repetitive Exercise. Front. Physiol. 2018, 9, 1136. [Google Scholar] [CrossRef]
- Hakansson, K.E.J.; Goossens, E.A.C.; Trompet, S.; van Ingen, E.; de Vries, M.R.; van der Kwast, R.; Ripa, R.S.; Kastrup, J.; Hohensinner, P.J.; Kaun, C.; et al. Genetic associations and regulation of expression indicate an independent role for 14q32 snoRNAs in Human Cardiovascular Disease. Cardiovasc. Res. 2018. [Google Scholar] [CrossRef]
- Kiss, T. Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs. EMBO J. 2001, 20, 3617–3622. [Google Scholar] [CrossRef] [PubMed]
- Wezel, A.; Welten, S.M.; Razawy, W.; Lagraauw, H.M.; de Vries, M.R.; Goossens, E.A.; Boonstra, M.C.; Hamming, J.F.; Kandimalla, E.R.; Kuiper, J.; et al. Inhibition of MicroRNA-494 Reduces Carotid Artery Atherosclerotic Lesion Development and Increases Plaque Stability. Ann. Surg. 2015, 262, 841–847, discussion 847–848. [Google Scholar] [CrossRef] [PubMed]
- Welten, S.M.; Bastiaansen, A.J.; de Jong, R.C.; de Vries, M.R.; Peters, E.A.; Boonstra, M.C.; Sheikh, S.P.; La Monica, N.; Kandimalla, E.R.; Quax, P.H.; et al. Inhibition of 14q32 MicroRNAs miR-329, miR-487b, miR-494 and miR-495 increases neovascularization and blood flow recovery after ischemia. Circ. Res. 2014, 115, 696–708. [Google Scholar] [CrossRef] [PubMed]
- Welten, S.M.; Goossens, E.A.; Quax, P.H.; Nossent, A.Y. The multifactorial nature of microRNAs in vascular remodelling. Cardiovasc. Res. 2016, 110, 6–22. [Google Scholar] [CrossRef] [PubMed]
- Welten, S.M.J.; de Jong, R.C.M.; Wezel, A.; de Vries, M.R.; Boonstra, M.C.; Parma, L.; Jukema, J.W.; van der Sluis, T.C.; Arens, R.; Bot, I.; et al. Inhibition of 14q32 microRNA miR-495 reduces lesion formation, intimal hyperplasia and plasma cholesterol levels in experimental restenosis. Atherosclerosis 2017, 261, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Nossent, A.Y.; Eskildsen, T.V.; Andersen, L.B.; Bie, P.; Bronnum, H.; Schneider, M.; Andersen, D.C.; Welten, S.M.; Jeppesen, P.L.; Hamming, J.F.; et al. The 14q32 microRNA-487b targets the antiapoptotic insulin receptor substrate 1 in hypertension-induced remodeling of the aorta. Ann. Surg. 2013, 258, 743–751, discussion 752–743. [Google Scholar] [CrossRef]
- Welten, G.M.; Schouten, O.; Hoeks, S.E.; Chonchol, M.; Vidakovic, R.; van Domburg, R.T.; Bax, J.J.; van Sambeek, M.R.; Poldermans, D. Long-term prognosis of patients with peripheral arterial disease: A comparison in patients with coronary artery disease. J. Am. Coll. Cardiol. 2008, 51, 1588–1596. [Google Scholar] [CrossRef]
- Gremmel, T.; Xhelili, E.; Steiner, S.; Koppensteiner, R.; Kopp, C.W.; Panzer, S. Response to antiplatelet therapy and platelet reactivity to thrombin receptor activating peptide-6 in cardiovascular interventions: Differences between peripheral and coronary angioplasty. Atherosclerosis 2014, 232, 119–124. [Google Scholar] [CrossRef]
- Xi, S.; Xu, H.; Shan, J.; Tao, Y.; Hong, J.A.; Inchauste, S.; Zhang, M.; Kunst, T.F.; Mercedes, L.; Schrump, D.S. Cigarette smoke mediates epigenetic repression of miR-487b during pulmonary carcinogenesis. J. Clin. Investig. 2013, 123, 1241–1261. [Google Scholar] [CrossRef]
- Gremmel, T.; Steiner, S.; Seidinger, D.; Koppensteiner, R.; Panzer, S.; Kopp, C.W. Smoking promotes clopidogrel-mediated platelet inhibition in patients receiving dual antiplatelet therapy. Thromb. Res. 2009, 124, 588–591. [Google Scholar] [CrossRef]
- Bliden, K.P.; Dichiara, J.; Lawal, L.; Singla, A.; Antonino, M.J.; Baker, B.A.; Bailey, W.L.; Tantry, U.S.; Gurbel, P.A. The association of cigarette smoking with enhanced platelet inhibition by clopidogrel. J. Am. Coll. Cardiol. 2008, 52, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Badrnya, S.; Schrottmaier, W.C.; Kral, J.B.; Yaiw, K.C.; Volf, I.; Schabbauer, G.; Soderberg-Naucler, C.; Assinger, A. Platelets mediate oxidized low-density lipoprotein-induced monocyte extravasation and foam cell formation. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 571–580. [Google Scholar] [CrossRef] [PubMed]
- van Ingen, E.; Foks, A.C.; Kröner, M.J.; Kuiper, J.; Quax, P.H.A.; Bot, I.; Nossent, A.Y. Antisense Oligonucleotide Inhibition of MicroRNA-494 Halts Atherosclerotic Plaque Progression and Promotes Plaque Stabilization. Mol. Ther. Nucleic Acids 2019, 18, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, L.C.; Simon, L.M.; Montoya, R.T.; Holinstat, M.; Chen, E.S.; Bergeron, A.; Kong, X.; Nagalla, S.; Mohandas, N.; Cohen, D.E.; et al. Racial differences in human platelet PAR4 reactivity reflect expression of PCTP and miR-376c. Nat. Med. 2013, 19, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Jorjani, H.; Kehr, S.; Jedlinski, D.J.; Gumienny, R.; Hertel, J.; Stadler, P.F.; Zavolan, M.; Gruber, A.R. An updated human snoRNAome. Nucleic. Acids. Res. 2016, 44, 5068–5082. [Google Scholar] [CrossRef] [PubMed]
- Gremmel, T.; Steiner, S.; Seidinger, D.; Koppensteiner, R.; Panzer, S.; Kopp, C.W. In vivo and protease-activated receptor-1-mediated platelet activation but not response to antiplatelet therapy predict two-year outcomes after peripheral angioplasty with stent implantation. Thromb. Haemost. 2014, 111, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Gremmel, T.; Steiner, S.; Seidinger, D.; Koppensteiner, R.; Panzer, S.; Kopp, C.W. Comparison of methods to evaluate clopidogrel-mediated platelet inhibition after percutaneous intervention with stent implantation. Thromb. Haemost. 2009, 101, 333–339. [Google Scholar]
- Gremmel, T.; Koppensteiner, R.; Panzer, S. Comparison of Aggregometry with Flow Cytometry for the Assessment of Agonists—Induced Platelet Reactivity in Patients on Dual Antiplatelet Therapy. PLoS ONE 2015, 10, e0129666. [Google Scholar] [CrossRef]
- Gremmel, T.; Ay, C.; Riedl, J.; Kopp, C.W.; Eichelberger, B.; Koppensteiner, R.; Panzer, S. Platelet-specific markers are associated with monocyte-platelet aggregate formation and thrombin generation potential in advanced atherosclerosis. Thromb. Haemost. 2016, 115, 615–621. [Google Scholar] [CrossRef]
- Gremmel, T.; Panzer, S. Clinical, genetic and confounding factors determine the dynamics of the in vitro response/non response to clopidogrel. Thromb. Haemost. 2011, 106, 211–218. [Google Scholar] [CrossRef]
- Cavaille, J.; Seitz, H.; Paulsen, M.; Ferguson-Smith, A.C.; Bachellerie, J.P. Identification of tandemly-repeated C/D snoRNA genes at the imprinted human 14q32 domain reminiscent of those at the Prader-Willi/Angelman syndrome region. Hum. Mol. Genet. 2002, 11, 1527–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| snoRNA | Hypertension | n | Mean | ∆ | p |
| SNORD112 | Yes | 8 | 0.07 | 0.01 | 0.99 |
| No | 93 | 0.08 | |||
| SNORD113.2 | Yes | 7 | 9.37 | 0.36 | 0.90 |
| No | 87 | 9.72 | |||
| SNORD113.6 | Yes | 7 | 0.08 | 0.04 | 0.34 |
| No | 85 | 0.04 | |||
| SNORD114.1 | Yes | 8 | 9.59 | 2.48 | 0.37 |
| No | 90 | 7.11 | |||
| Hyperlipidemia | n | Mean | ∆ | p | |
| SNORD112 | Yes | 8 | 0.07 | 0.01 | 0.39 |
| No | 93 | 0.08 | |||
| SNORD113.2 | Yes | 7 | 9.37 | 0.36 | 0.53 |
| No | 87 | 9.72 | |||
| SNORD113.6 | Yes | 7 | 0.08 | 0.04 | 0.43 |
| No | 85 | 0.04 | |||
| SNORD114.1 | Yes | 8 | 9.59 | 2.48 | 0.51 |
| No | 90 | 7.11 | |||
| Diabetes Mellitus | n | Mean | ∆ | p | |
| SNORD112 | Yes | 65 | 0.074 | 0.03 | 0.21 |
| No | 36 | 0.10 | |||
| SNORD113.2 | Yes | 61 | 9.15 | 1.56 | 0.24 |
| No | 33 | 10.71 | |||
| SNORD113.6 | Yes | 58 | 0.03 | 0.03 | 0.90 |
| No | 34 | 0.06 | |||
| SNORD114.1 | Yes | 65 | 6.97 | 1.01 | 0.94 |
| No | 33 | 7.98 | |||
| Smoking | n | Mean | ∆ | p | |
| SNORD112 | Yes | 57 | 0.08 | 0.01 | 0.41 |
| No | 44 | 0.09 | |||
| SNORD113.2 | Yes | 51 | 10.32 | 1.35 | 0.35 |
| No | 43 | 8.97 | |||
| SNORD113.6 | Yes | 52 | 0.05 | 0.02 | 0.80 |
| No | 40 | 0.03 | |||
| SNORD114.1 | Yes | 56 | 8.68 | 3.2 | 0.031 |
| No | 42 | 5.48 |
| snoRNA | Risk Factor | B | p |
|---|---|---|---|
| SNORD112 | Lpa | −0.228 | 0.025 |
| SNORD113.2 | Triglycerides | 0.219 | 0.034 |
| SNORD113.2 | WBC | −0.746 | 0.098 |
| SNORD113.6 | WBC | 0.012 | 0.026 |
| Risk Factor | SNORD112 | n | Mean | ∆ | p |
| Lpa | Low | 72.47 | 21.67 | 0.038 | |
| High | 50.80 | ||||
| SNORD113.2 | n | Mean | ∆ | p | |
| Triglycerides | Low | 155.49 | 58.07 | 0.27 | |
| High | 213.56 | ||||
| WBC | Low | 9.19 | 0.70 | 0.10 | |
| High | 8.49 | ||||
| SNORD114.1 | n | Mean | ∆ | p | |
| WBC | Low | 9.12 | 0.73 | 0.057 | |
| High | 8.39 |
| Parameter of Platelet Function | SNORD113.2 | n | Mean | ∆ | p |
| In vivo P-Selectin | Low | 46 | 3.60 | 0.31 | 0.07 |
| High | 49 | 3.29 | |||
| ADP-inducible P-selectin | Low | 46 | 15.23 | 0.27 | 0.50 |
| High | 49 | 14.96 | |||
| In vivo activated GPIIb/IIIa | Low | 46 | 2.99 | 0.21 | 0.18 |
| High | 49 | 2.78 | |||
| ADP-inducible GPIIb/IIIa | Low | 46 | 12.02 | 0.25 | 0.69 |
| High | 49 | 11.77 | |||
| In vivo MPA | Low | 38 | 30.29 | 6.82 | 0.039 |
| High | 42 | 23.47 | |||
| ADP-inducible MPA | Low | 38 | 54.67 | 9.58 | 0.015 |
| High | 42 | 45.09 | |||
| PRI | Low | 48 | 49.34 | 7.54 | 0.09 |
| High | 50 | 41.80 | |||
| Parameter of Platelet Function | SNORD114.1 | n | Mean | ∆ | p |
| In vivo P-Selectin | Low | 49 | 3.60 | 0.31 | 0.11 |
| High | 48 | 3.29 | |||
| ADP-inducible P-selectin | Low | 49 | 16.65 | 2.46 | 0.17 |
| High | 48 | 14.20 | |||
| In vivo activated GPIIb/IIIa | Low | 49 | 2.92 | 0.05 | 0.51 |
| High | 48 | 2.87 | |||
| ADP-inducible GPIIb/IIIa | Low | 49 | 12.13 | 0.03 | 0.90 |
| High | 48 | 12.10 | |||
| In vivo MPA | Low | 44 | 31.18 | 9.23 | 0.015 |
| High | 38 | 21.95 | |||
| ADP-inducible MPA | Low | 44 | 55.22 | 10.57 | 0.010 |
| High | 39 | 44.65 | |||
| PRI | Low | 50 | 48.79 | 5.80 | 0.24 |
| High | 50 | 42.99 |
| SNORD113.2 | ||||
| Parameter of Platelet Function | B | p | BAdjusted | pAdjusted |
| In vivo P-Selectin | −0.023 | 0.055 | −0.026 | 0.033 |
| ADP-inducible P-selectin | −0.078 | 0.557 | −0.112 | 0.406 |
| In vivo GPIIb/IIIa | −0.017 | 0.080 | −0.020 | 0.035 |
| ADP-inducible GPIIb/IIIa | −0.062 | 0.499 | −0.098 | 0.283 |
| In vivo MPA | −0.441 | 0.028 | −0.524 | 0.010 |
| ADP-inducible MPA | −0.653 | 0.002 | −0.709 | 0.001 |
| PRI | −0.600 | 0.029 | −0.596 | 0.034 |
| SNORD114.1 | ||||
| Parameter of Platelet Function | B | p | BAdjusted * | pAdjusted * |
| In vivo P-Selectin | −0.026 | 0.066 | −0.029 | 0.048 |
| ADP-inducible P-selectin | −0.141 | 0.393 | −0.192 | 0.255 |
| In vivo GPIIb/IIIa | −0.006 | 0.613 | −0.012 | 0.296 |
| ADP-inducible GPIIb/IIIa | −0.029 | 0.796 | −0.071 | 0.526 |
| In vivo MPA | −0.398 | 0.087 | −0.352 | 0.137 |
| ADP-inducible MPA | −0.532 | 0.037 | −0.559 | 0.034 |
| PRI | −0.480 | 0.161 | −0.515 | 0.147 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nossent, A.Y.; Ektefaie, N.; Wojta, J.; Eichelberger, B.; Kopp, C.; Panzer, S.; Gremmel, T. Plasma Levels of snoRNAs are Associated with Platelet Activation in Patients with Peripheral Artery Disease. Int. J. Mol. Sci. 2019, 20, 5975. https://doi.org/10.3390/ijms20235975
Nossent AY, Ektefaie N, Wojta J, Eichelberger B, Kopp C, Panzer S, Gremmel T. Plasma Levels of snoRNAs are Associated with Platelet Activation in Patients with Peripheral Artery Disease. International Journal of Molecular Sciences. 2019; 20(23):5975. https://doi.org/10.3390/ijms20235975
Chicago/Turabian StyleNossent, Anne Yaël, Neda Ektefaie, Johann Wojta, Beate Eichelberger, Christoph Kopp, Simon Panzer, and Thomas Gremmel. 2019. "Plasma Levels of snoRNAs are Associated with Platelet Activation in Patients with Peripheral Artery Disease" International Journal of Molecular Sciences 20, no. 23: 5975. https://doi.org/10.3390/ijms20235975
APA StyleNossent, A. Y., Ektefaie, N., Wojta, J., Eichelberger, B., Kopp, C., Panzer, S., & Gremmel, T. (2019). Plasma Levels of snoRNAs are Associated with Platelet Activation in Patients with Peripheral Artery Disease. International Journal of Molecular Sciences, 20(23), 5975. https://doi.org/10.3390/ijms20235975