Estimation of Forces on Actin Filaments in Living Muscle from X-ray Diffraction Patterns and Mechanical Data



Abstract
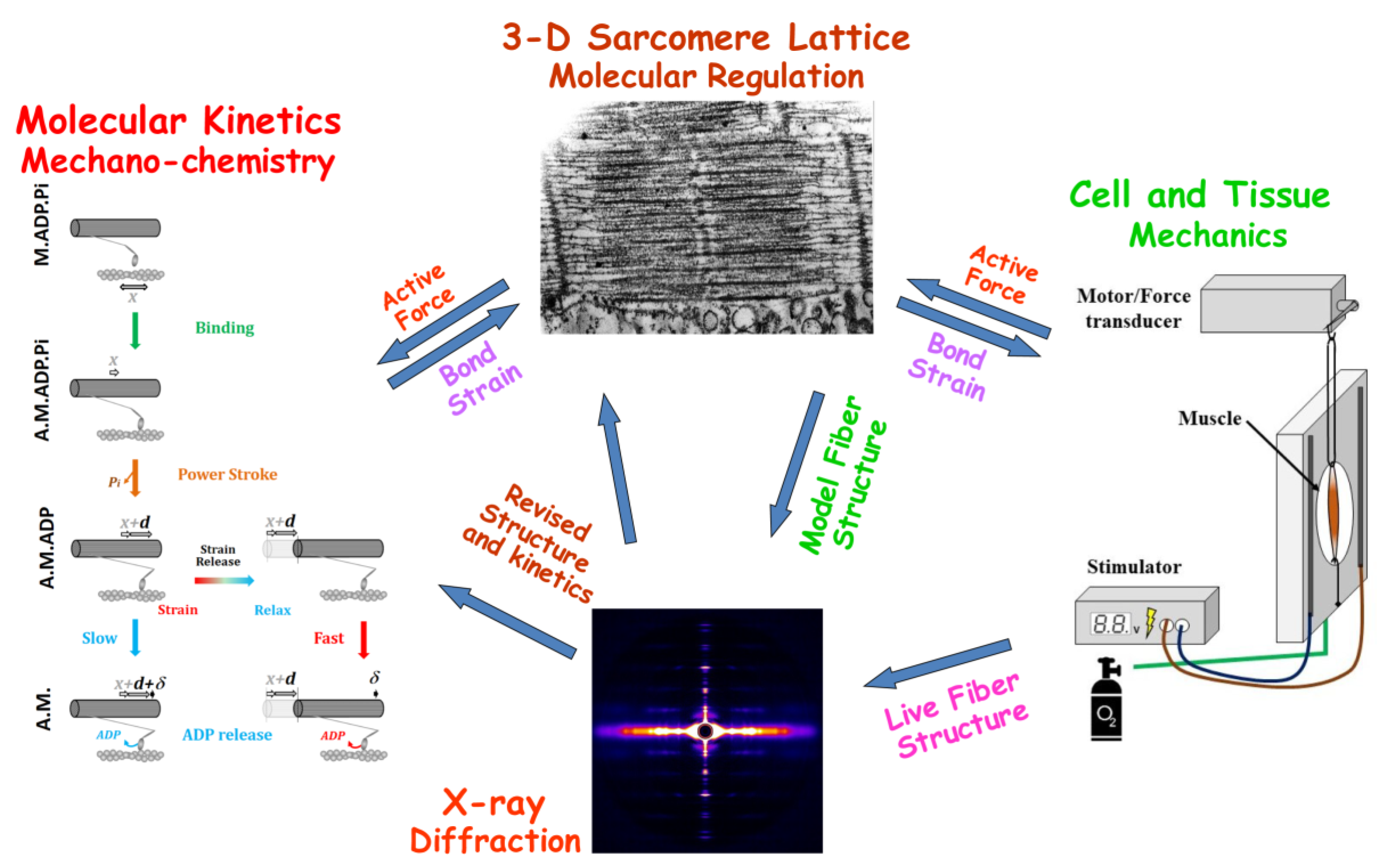
:1. Introduction
2. Simulation of X-ray Diffraction Patterns
2.1. Simulation of Contraction in Muscle Fibers—The MUSICO Platform
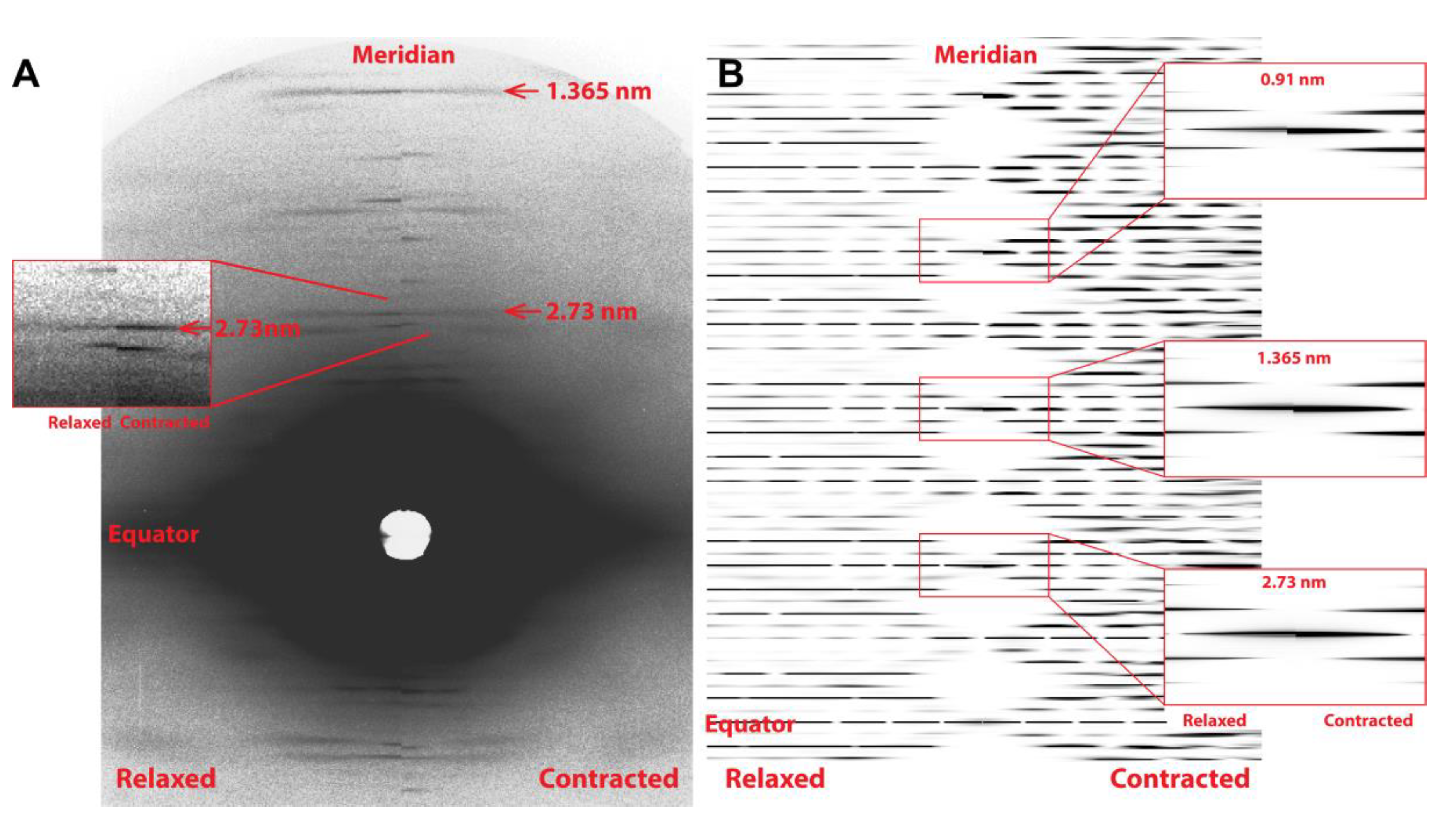
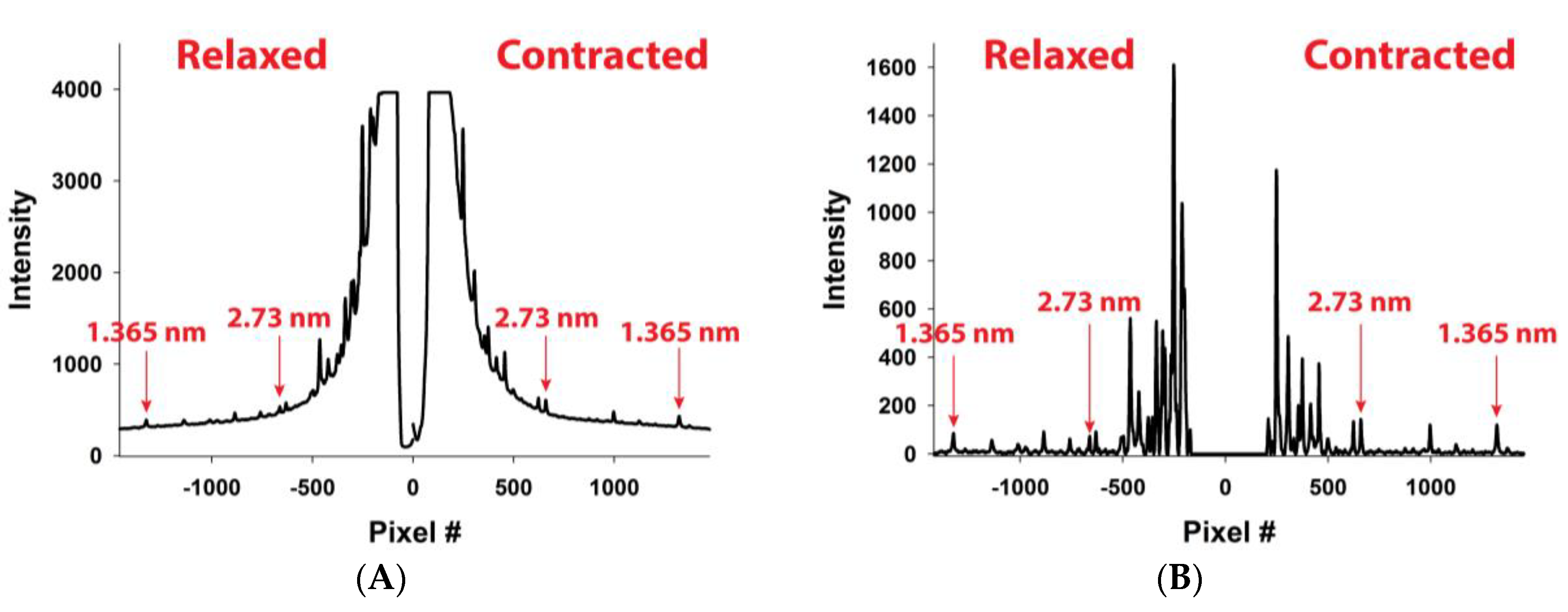
2.2. MUSICO Predictions of X-ray Diffraction Patterns
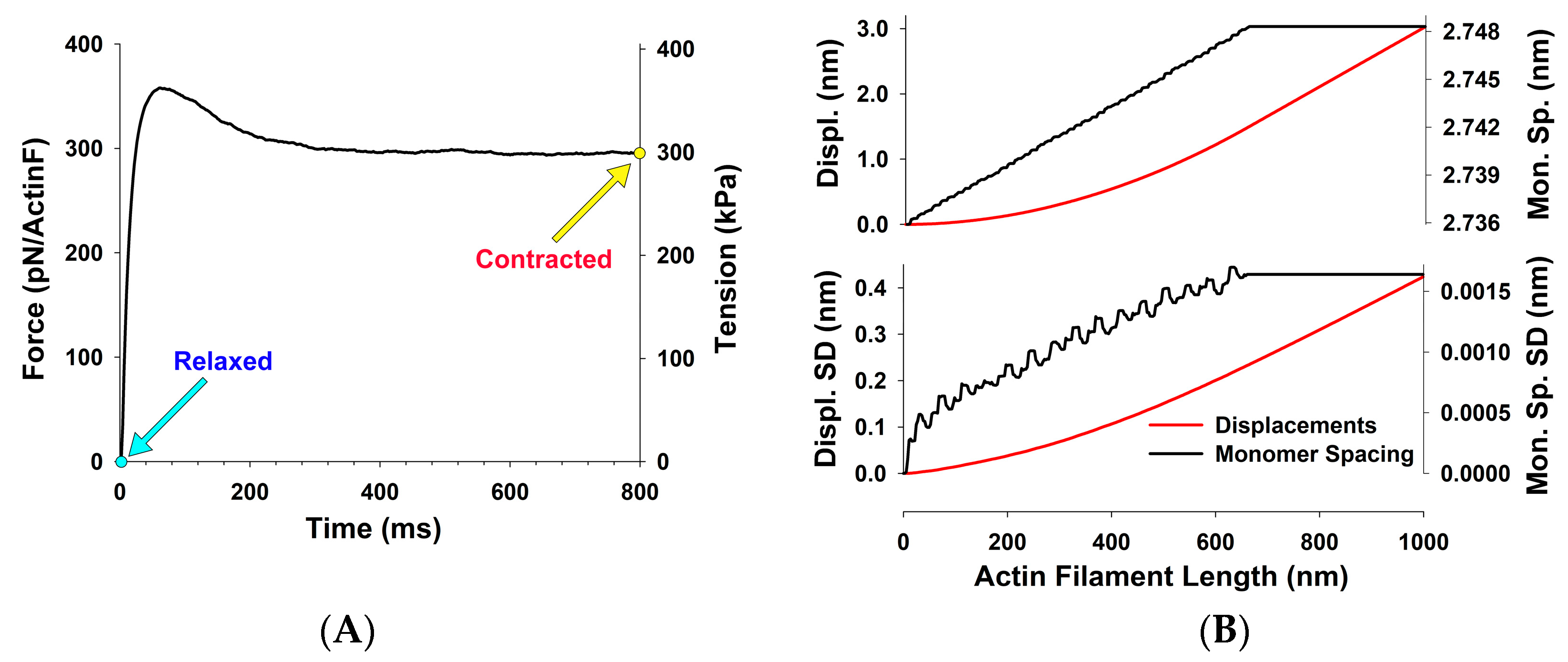
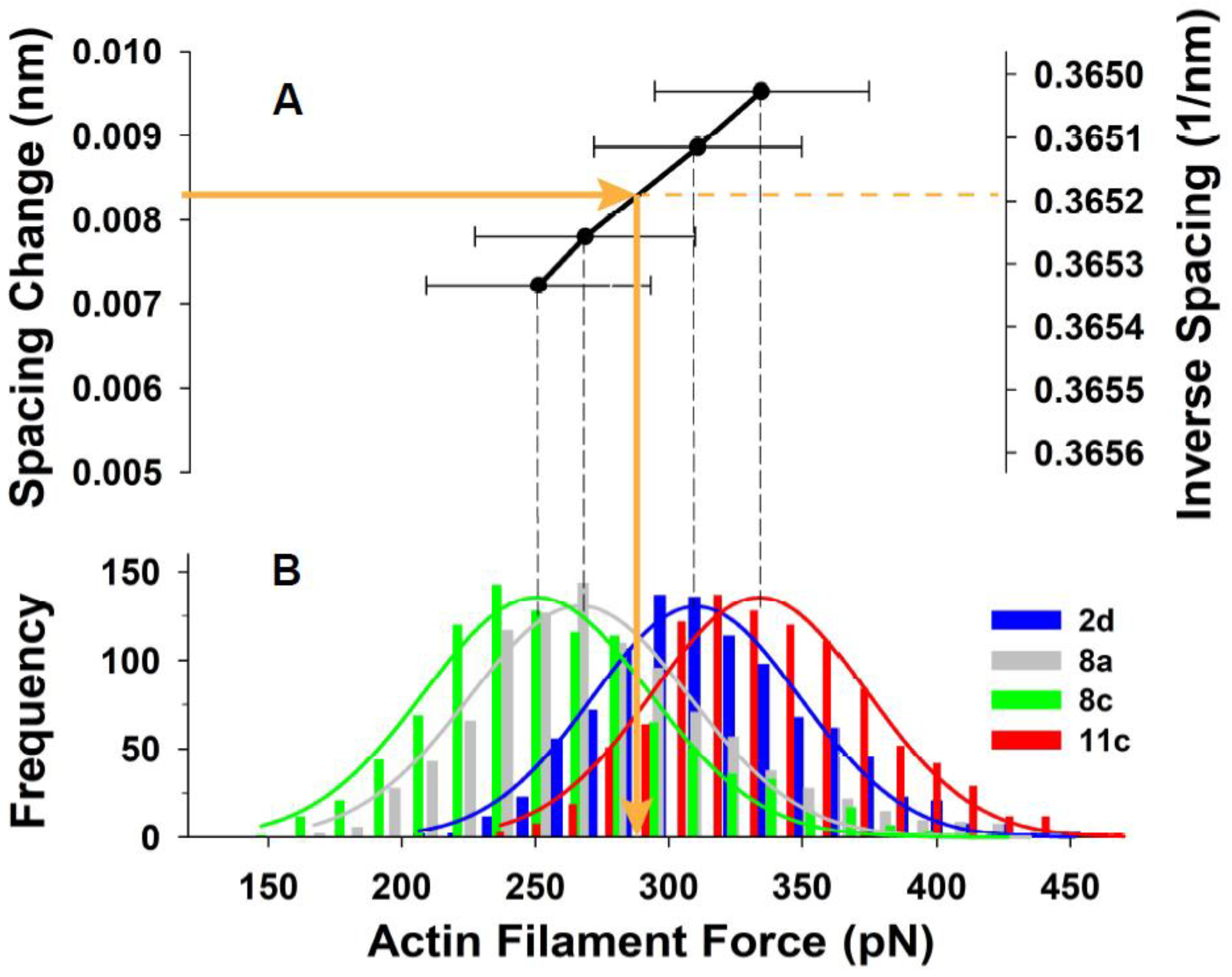
2.3. Estimation of Thin Filament Forces in Contracting Muscle
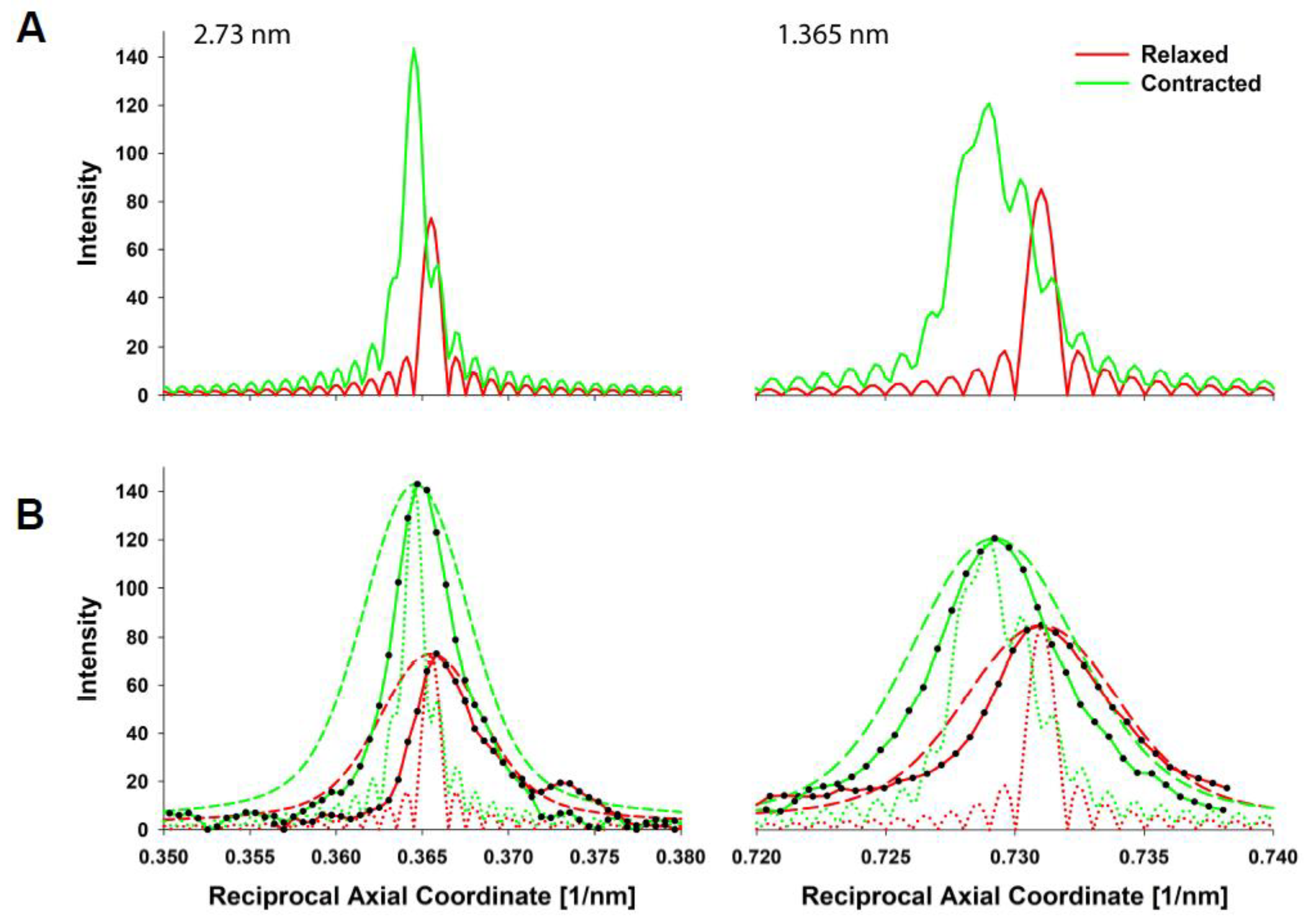
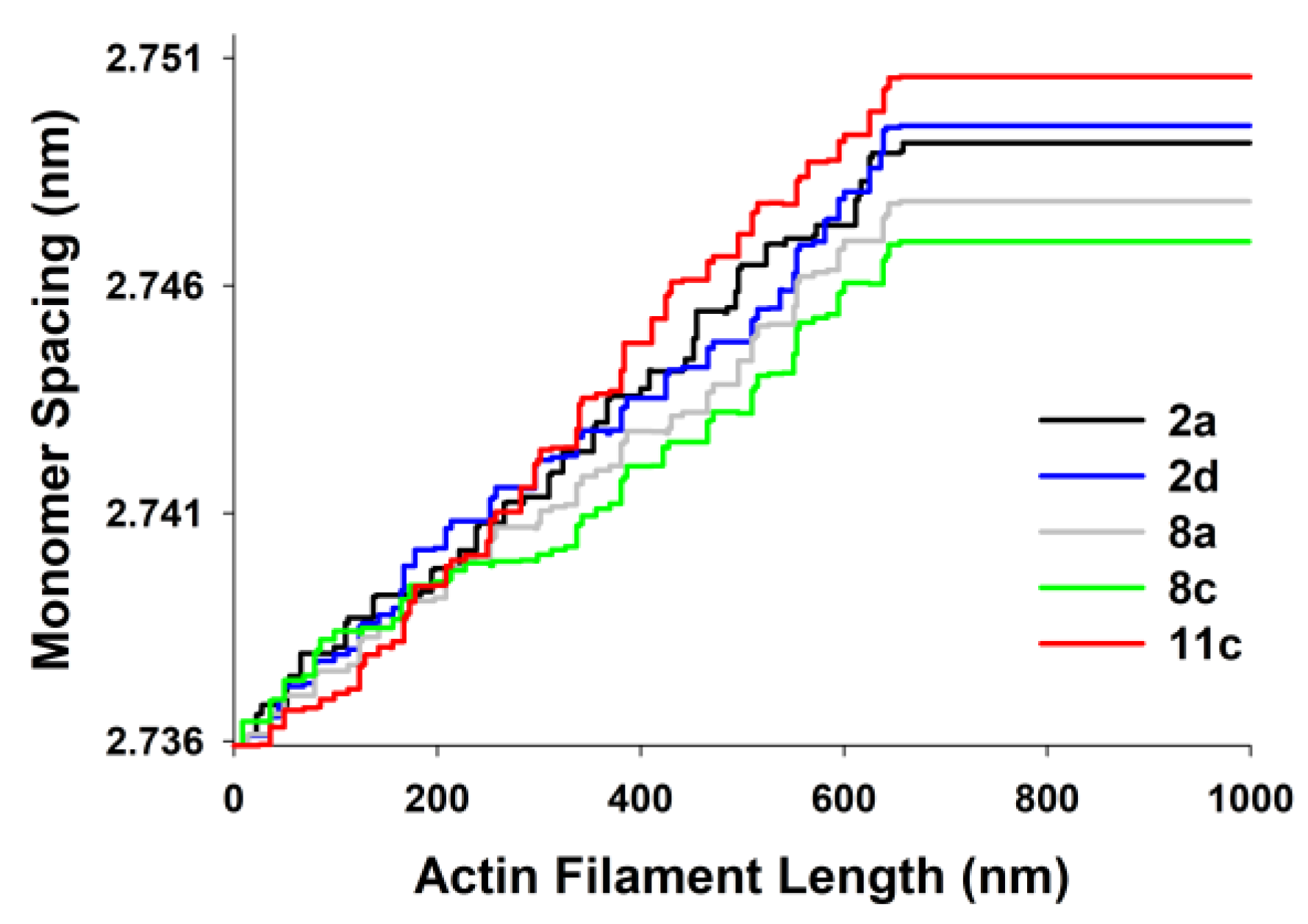
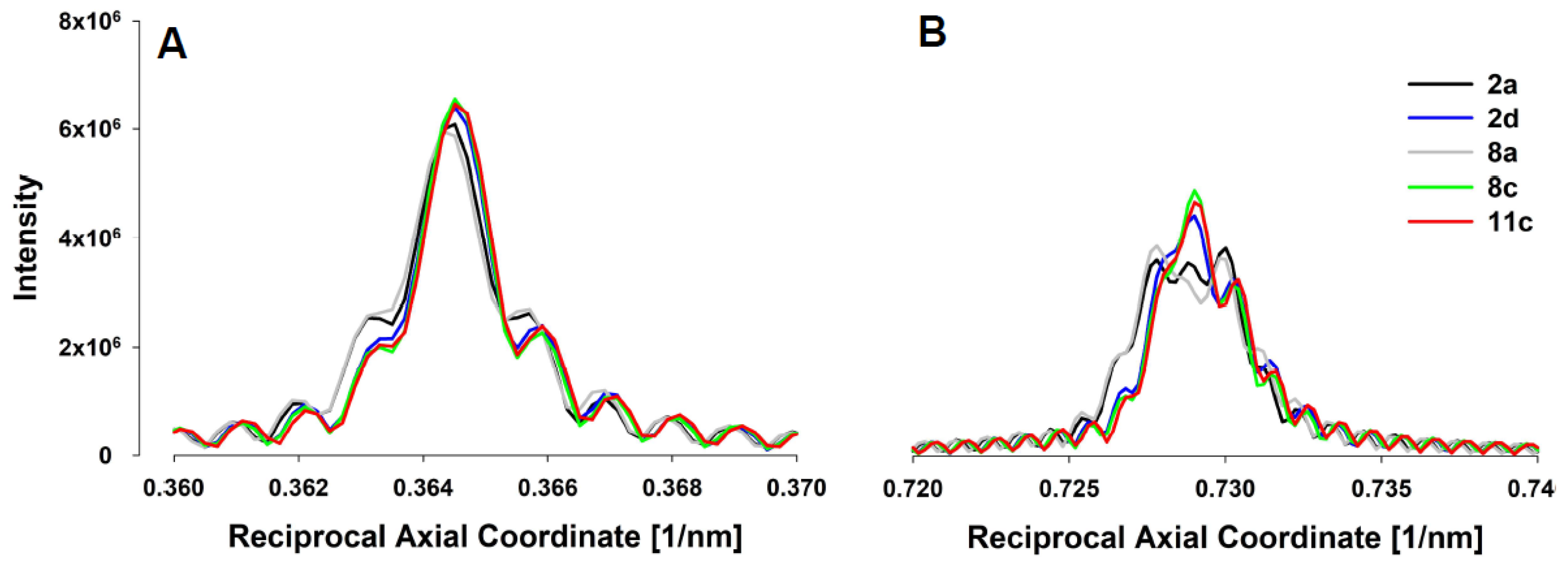
3. Results
4. Discussion
Limitations of the Study
5. Materials and Methods
5.1. MUSICO Model Parameters
5.1.1. Sarcomere Geometry and Myofilament Elasticity
5.1.2. Three-State Crossbridge Model Parameters
5.2. Conversion between Tension and Isometric Force per Actin Filament
5.3. X-ray Diffraction Experiments
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | Three dimensional |
| 2D | Two dimensional |
| MUSICO | Muscle Simulation Code |
| ATP | Adenosine tri-phosphate |
| ADP | Adenosine di-phosphate |
| Pi | Inorganic phosphate |
| ATP-ase | Rate of ATP consumption |
| ActinF | Actin filament |
| SD | Standard deviation |
References
- Alenghat, F.J.; Ingber, D.E. Mechanotransduction: All signals point to cytoskeleton, matrix, and integrins. Sci. Signal. 2002, 2002, pe6. [Google Scholar] [CrossRef]
- Lauffenburger, D.A.; Horwitz, A.F. Cell migration: A physically integrated molecular process. Cell 1996, 84, 359–369. [Google Scholar] [CrossRef]
- Sheetz, M.P.; Felsenfeld, D.P.; Galbraith, C.G. Cell migration: Regulation of force on extracellular-matrix-integrin complexes. Trends Cell Biol. 1998, 8, 51–54. [Google Scholar] [CrossRef]
- Wang, N.; Butler, J.P.; Ingber, D.E. Mechanotransduction across the cell surface and through the cytoskeleton. Science 1993, 260, 1124–1127. [Google Scholar] [CrossRef]
- Evans, E. Looking inside molecular bonds at biological interfaces with dynamic force spectroscopy. Biophys. Chem. 1999, 82, 83–97. [Google Scholar] [CrossRef]
- Evans, E. Probing the relation between force—Lifetime—and chemistry in single molecular bonds. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 105–128. [Google Scholar] [CrossRef]
- Merkel, R.; Nassoy, P.; Leung, A.; Ritchie, K.; Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature 1999, 397, 50–53. [Google Scholar] [CrossRef]
- Mijailovich, S.M.; Stojanovic, B.; Nedic, D.; Svicevic, M.; Geeves, M.A.; Irving, T.C.; Granzier, H. Nebulin and Titin Modulate Crossbridge Cycling and Length Dependent Calcium Sensitivity. J. Gen. Physiol. 2019, 151, 680–704. [Google Scholar] [CrossRef] [PubMed]
- Linari, M.; Dobbie, I.; Reconditi, M.; Koubassova, N.; Irving, M.; Piazzesi, G.; Lombardi, V. The stiffness of skeletal muscle in isometric contraction and rigor: The fraction of myosin heads bound to actin. Biophys. J. 1998, 74, 2459–2473. [Google Scholar] [CrossRef]
- Wang, N.; Naruse, K.; Stamenovic, D.; Fredberg, J.J.; Mijailovich, S.M.; Tolic-Norrelykke, I.M.; Polte, T.; Mannix, R.; Ingber, D.E. Mechanical behavior in living cells consistent with the tensegrity model. Proc. Natl. Acad. Sci. USA 2001, 98, 7765–7770. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Tolic-Norrelykke, I.M.; Chen, J.; Mijailovich, S.M.; Butler, J.P.; Fredberg, J.J.; Stamenovic, D. Cell prestress. I. Stiffness and prestress are closely associated in adherent contractile cells. Am. J. Physiol. Cell Physiol. 2002, 282, C606–C616. [Google Scholar] [CrossRef] [PubMed]
- Huxley, H.E.; Stewart, A.; Sosa, H.; Irving, T. X-ray diffraction measurements of the extensibility of actin and myosin filaments in contracting muscle. Biophys. J. 1994, 67, 2411–2421. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Sugimoto, Y.; Tanaka, H.; Ueno, Y.; Takezawa, Y.; Amemiya, Y. X-ray diffraction evidence for the extensibility of actin and myosin filaments during muscle contraction. Biophys. J. 1994, 67, 2422–2435. [Google Scholar] [CrossRef]
- Prodanovic, M.; Irving, T.C.; Mijailovich, S.M. X-ray diffraction from nonuniformly stretched helical molecules. J. Appl. Crystallogr. 2016, 49, 784–797. [Google Scholar] [CrossRef] [PubMed]
- Mijailovich, S.M.; Kayser-Herold, O.; Stojanovic, B.; Nedic, D.; Irving, T.C.; Geeves, M.A. Three-dimensional stochastic model of actin-myosin binding in the sarcomere lattice. J. Gen. Physiol. 2016, 148, 459–488. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.; Geeves, M.A.; Sleep, J.; Mijailovich, S.M. Towards a unified theory of muscle contraction. I: Foundations. Ann. Biomed. Eng. 2008, 36, 1624–1640. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.; Mijailovich, S.M. Toward a unified theory of muscle contraction. II: Predictions with the mean-field approximation. Ann. Biomed. Eng. 2008, 36, 1353–1371. [Google Scholar] [CrossRef] [PubMed]
- Mijailovich, S.M.; Kayser-Herald, O.; Moss, R.L.; Geeves, M.A. Stochastic Model of Contraction and Thin Filament Regulation in Exact 3D Sarcomere Geometry. Biophys. J. 2008, 94, 404a. [Google Scholar]
- Mijailovich, S.M.; Kayser-Herald, O.; Moss, R.L.; Geeves, M.A. Thin Filament Regulation of Relaxation in 3D Multi-Sarcomere Geometry. Biophys. J. 2009, 96, 201a. [Google Scholar] [CrossRef]
- Mijailovich, S.M.; Stojanovic, B.; Nedic, D.; Geeves, M.A. Activation and Relaxation Kinetics in Skeletal and Cardiac Muscles. Biophys. J. 2015, 108, 337a–338a. [Google Scholar] [CrossRef]
- Mijailovich, S.M.; Stojanovic, B.; Nedic, D.; Svicevic, M.; Gilbert, R.J.; Geeves, M.A.; Irving, T. Modulation of Crossbridge Cycling Kinetics and Length Dependent Calcium Sensitivity by Titin and Nebulin. Biophys. J. 2013, 104, 310a. [Google Scholar] [CrossRef]
- Mijailovich, S.M.; Nedic, D.; Stojanovic, B.; Geeves, M.A. Effect of Nonlinear Crossbridge Elasticity on Kinetics of Sarcomeric Contraction. Biophys. J. 2014, 106, 158a. [Google Scholar] [CrossRef]
- Gautel, M. The sarcomeric cytoskeleton: Who picks up the strain? Curr. Opin. Cell Biol. 2011, 23, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Hoshijima, M. Mechanical stress-strain sensors embedded in cardiac cytoskeleton: Z disk, titin, and associated structures. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1313–H1325. [Google Scholar] [CrossRef]
- Linke, W.A.; Kruger, M. The giant protein titin as an integrator of myocyte signaling pathways. Physiology 2010, 25, 186–198. [Google Scholar] [CrossRef]
- Gautel, M. The sarcomere and the nucleus: Functional links to hypertrophy, atrophy and sarcopenia. Adv. Exp. Med. Biol. 2008, 642, 176–191. [Google Scholar]
- Kontrogianni-Konstantopoulos, A.; Ackermann, M.A.; Bowman, A.L.; Yap, S.V.; Bloch, R.J. Muscle giants: Molecular scaffolds in sarcomerogenesis. Physiol. Rev. 2009, 89, 1217–1267. [Google Scholar] [CrossRef]
- Higuchi, H.; Yanagida, T.; Goldman, Y.E. Compliance of thin filaments in skinned fibers of rabbit skeletal muscle. Biophys. J. 1995, 69, 1000–1010. [Google Scholar] [CrossRef]
- Park-Holohan, S.; Linari, M.; Reconditi, M.; Fusi, L.; Brunello, E.; Irving, M.; Dolfi, M.; Lombardi, V.; West, T.G.; Curtin, N.A.; et al. Mechanics of myosin function in white muscle fibres of the dogfish, Scyliorhinus canicula. J. Physiol. 2012, 590, 1973–1988. [Google Scholar] [CrossRef]
- Tsaturyan, A.K.; Koubassova, N.; Ferenczi, M.A.; Narayanan, T.; Roessle, M.; Bershitsky, S.Y. Strong binding of myosin heads stretches and twists the actin helix. Biophys. J. 2005, 88, 1902–1910. [Google Scholar] [CrossRef]
- Kojima, H.; Ishijima, A.; Yanagida, T. Direct measurement of stiffness of single actin filaments with and without tropomyosin by in vitro nanomanipulation. Proc. Natl. Acad. Sci. USA 1994, 91, 12962–12966. [Google Scholar] [CrossRef] [PubMed]
- Isambert, H.; Venier, P.; Maggs, A.C.; Fattoum, A.; Kassab, R.; Pantaloni, D.; Carlier, M.F. Flexibility of actin filaments derived from thermal fluctuations. Effect of bound nucleotide, phalloidin, and muscle regulatory proteins. J. Biol. Chem. 1995, 270, 11437–11444. [Google Scholar] [CrossRef] [PubMed]
- Kiss, B.; Lee, E.J.; Ma, W.; Li, F.; Tonino, P.; Mijailovich, S.M.; Irving, T.H.G. Nebulin stiffens the thin filament and augments cross-bridge interaction in skeletal muscle. Proc. Natl. Acad. Sci. USA 2018, 115, 10369–10374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunello, E.; Caremani, M.; Melli, L.; Linari, M.; Fernandez-Martinez, M.; Narayanan, T.; Irving, M.; Piazzesi, G.; Lombardi, V.; Reconditi, M. The contributions of filaments and cross-bridges to sarcomere compliance in skeletal muscle. J. Physiol. 2014, 592, 3881–3899. [Google Scholar] [CrossRef] [PubMed]
- Takezawa, Y.; Sugimoto, Y.; Wakabayashi, K. Extensibility of the actin and myosin filaments in various states of skeletal muscle as studied by X-ray diffraction. Adv. Exp. Med. Biol. 1998, 453, 309–316. [Google Scholar] [PubMed]
- Bordas, J.; Svensson, A.; Rothery, M.; Lowy, J.; Diakun, G.P.; Boesecke, P. Extensibility and symmetry of actin filaments in contracting muscles. Biophys. J. 1999, 77, 3197–3207. [Google Scholar] [CrossRef] [Green Version]
- Burgoyne, T.; Muhamad, F.; Luther, P.K. Visualization of cardiac muscle thin filaments and measurement of their lengths by electron tomography. Cardiovasc. Res. 2008, 77, 707–712. [Google Scholar] [CrossRef]
- Luther, P.K.; Bennett, P.M.; Knupp, C.; Craig, R.; Padron, R.; Harris, S.P.; Patel, J.; Moss, R.L. Understanding the organisation and role of myosin binding protein C in normal striated muscle by comparison with MyBP-C knockout cardiac muscle. J. Mol. Biol. 2008, 384, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Millman, B.M. The filament lattice of striated muscle. Physiol. Rev. 1998, 78, 359–391. [Google Scholar] [CrossRef]
- Matsubara, I.; Elliott, G.F. X-ray diffraction studies on skinned single fibres of frog skeletal muscle. J. Mol. Biol. 1972, 72, 657–669. [Google Scholar] [CrossRef]
- Duke, T.A. Molecular model of muscle contraction. Proc. Natl. Acad. Sci. USA 1999, 96, 2770–2775. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp. No: | Relax | Isometric | Del H | Strain | Mean Force | SD | Tension |
|---|---|---|---|---|---|---|---|
| nm | nm | nm | % | pN/ActinF | pN/ActinF | kPa | |
| 1 (2a) | 2.73015 | 2.73893 | 0.00878 | 0.322 | 309.7 | 41.1 | 313.4 |
| 2 (2d) | 2.73037 | 2.73924 | 0.00887 | 0.325 | 312.8 | 38.9 | 316.6 |
| 3 (8a) | 2.73234 | 2.74014 | 0.00780 | 0.285 | 274.9 | 41.3 | 278.2 |
| 4 (8c) | 2.72991 | 2.73713 | 0.00722 | 0.264 | 254.7 | 42.0 | 257.8 |
| 5 (11c) | 2.73171 | 2.74124 | 0.00953 | 0.349 | 335.9 | 39.9 | 340.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijailovich, S.M.; Prodanovic, M.; Irving, T.C. Estimation of Forces on Actin Filaments in Living Muscle from X-ray Diffraction Patterns and Mechanical Data. Int. J. Mol. Sci. 2019, 20, 6044. https://doi.org/10.3390/ijms20236044
Mijailovich SM, Prodanovic M, Irving TC. Estimation of Forces on Actin Filaments in Living Muscle from X-ray Diffraction Patterns and Mechanical Data. International Journal of Molecular Sciences. 2019; 20(23):6044. https://doi.org/10.3390/ijms20236044
Chicago/Turabian StyleMijailovich, Srboljub M., Momcilo Prodanovic, and Thomas C. Irving. 2019. "Estimation of Forces on Actin Filaments in Living Muscle from X-ray Diffraction Patterns and Mechanical Data" International Journal of Molecular Sciences 20, no. 23: 6044. https://doi.org/10.3390/ijms20236044
APA StyleMijailovich, S. M., Prodanovic, M., & Irving, T. C. (2019). Estimation of Forces on Actin Filaments in Living Muscle from X-ray Diffraction Patterns and Mechanical Data. International Journal of Molecular Sciences, 20(23), 6044. https://doi.org/10.3390/ijms20236044