Proteomic Analysis of Cyclic Ketamine Compounds Ability to Induce Neural Differentiation in Human Adult Mesenchymal Stem Cells




Abstract
:
1. Introduction
2. Results
2.1. Microscopy
2.2. iTRAQ Proteome Comparisons of Chemically Induced hADSCs toward Neural Lineage
2.3. Interaction Network Analysis of CK Treated ADSC Proteomes
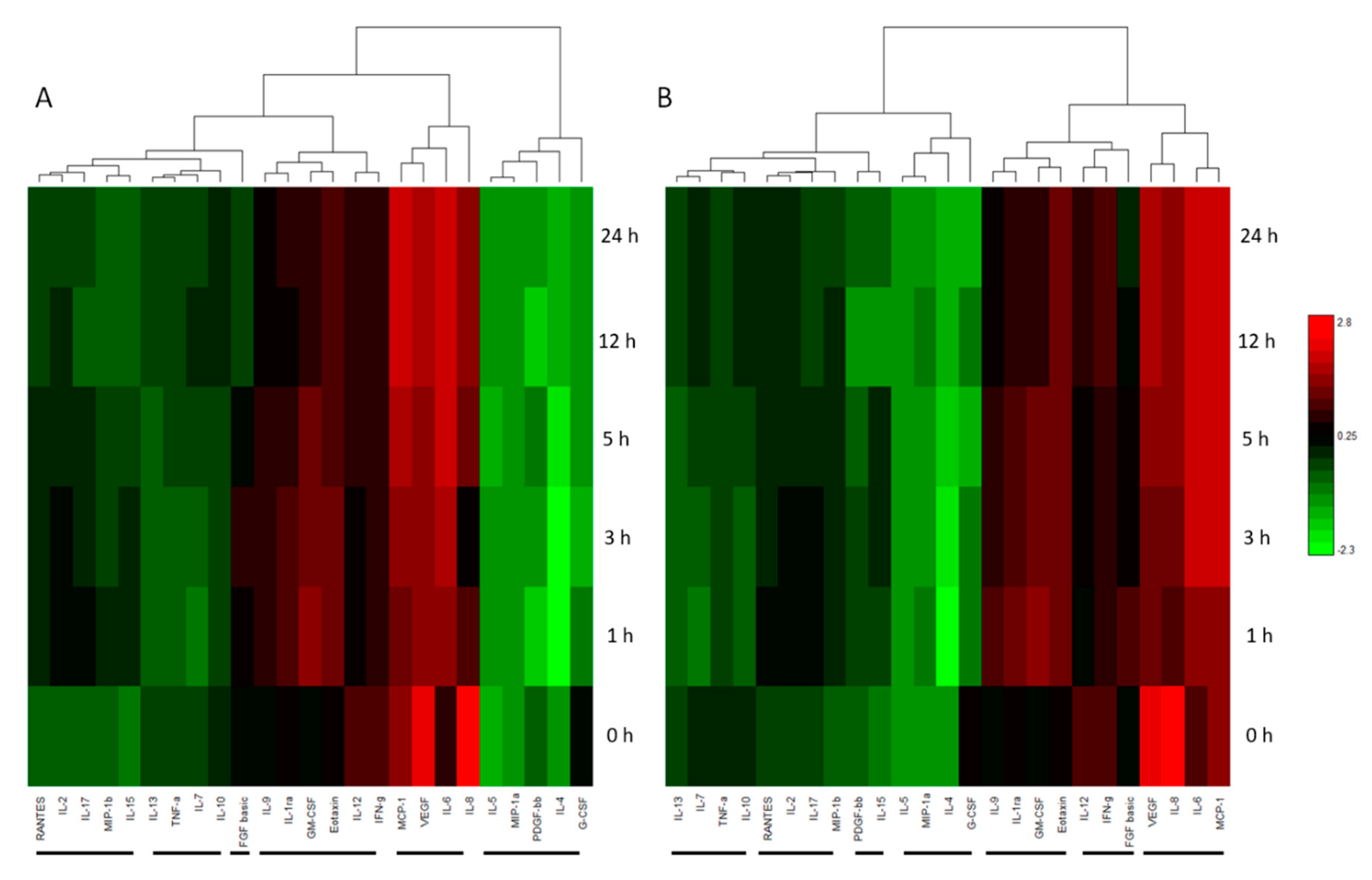
2.4. Cytokine Levels
3. Discussion
3.1. Neurogenic Related Roles of Identified Proteins
3.2. Neurogenic Roles of Cytokines
4. Materials and Methods
4.1. Cell Culture
4.1.1. Human Adipose Derived Stem Cells Harvest and Cell Culture
4.1.2. Chemical Induction for Differentiation
4.1.3. Glioblastoma Cell Culture
4.2. Microscopy
Cell Counts
4.3. Protein Extraction
4.4. iTRAQ
4.5. Bioplex
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
Abbreviations
| AECK | S-aminoethyl-l-cysteine ketamine |
| BHA | butylated hydroxyanisole |
| BME | beta mercaptoethanol |
| CK | cyclic ketamine |
| DMSO | dimethylsulfoxide |
| hADSCs | human adipose derived stem cells |
| iTRAQ | Isobaric tag for relative and absolute quantitation |
| LK | lanthionine ketamine |
| LKEE | lanthionine ketamine ethyl ester |
| MSC | mesenchymal stromal/stem cells |
| MSMS | tandem mass spectrometry |
| PBS | phosphate buffered saline |
References
- Caplan, A.I.; Bruder, S.P. Mesenchymal stem cells: Building blocks for molecular medicine in the 21st century. Trends Mol. Med. 2001, 7, 259–264. [Google Scholar] [CrossRef]
- Da Silva Meirelles, L.; Chagastelles, P.C.; Nardi, N.B. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J. Cell Sci. 2006, 119, 2204–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco Lambert, A.P.; Fraga Zandonai, A.; Bonatto, D.; Cantarelli Machado, D.; Pêgas Henriques, J.A. Differentiation of human adipose-derived adult stem cells into neuronal tissue: Does it work? Differentiation 2009, 77, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Woodbury, D.; Schwarz, E.J.; Prockop, D.J.; Black, I.B. Adult rat and human bone marrow stromal cells differentiate into neurons. J. Neurosci. Res. 2000, 61, 364–370. [Google Scholar] [CrossRef] [Green Version]
- Mareschi, K.; Novara, M.; Rustichelli, D.; Ferrero, I.; Guido, D.; Carbone, E.; Medico, E.; Madon, E.; Vercelli, A.; Fagioli, F. Neural differentiation of human mesenchymal stem cells: Evidence for expression of neural markers and eag K+ channel types. Exp. Hematol. 2006, 34, 1563–1572. [Google Scholar] [CrossRef]
- Santos, J.; Milthorpe, B.K.; Herbert, B.R.; Padula, M.P. Proteomic Analysis of Human Adipose Derived Stem Cells during Small Molecule Chemical Stimulated Pre-neuronal Differentiation. Int. J. Stem Cells 2017. [Google Scholar] [CrossRef]
- Krabbe, C.; Zimmer, J.; Meyer, M. Neural transdifferentiation of mesenchymal stem cells—A critical review. APMIS 2005, 113, 831–844. [Google Scholar] [CrossRef]
- Hensley, K. Lanthionine-Related Compounds for the Treatment of Inflammatory Diseases. U.S. Patent 7683055B2, 23 March 2010. [Google Scholar]
- Hensley, K.; Christov, A.; Kamat, S.; Zhang, X.C.; Jackson, K.W.; Snow, S.; Post, J. Proteomic identification of binding partners for the brain metabolite lanthionine ketimine (LK) and documentation of LK effects on microglia and motoneuron cell cultures. J. Neurosci. 2010, 30, 2979–2988. [Google Scholar] [CrossRef]
- Hensley, K.; Venkova, K.; Christov, A. Emerging biological importance of central nervous system lanthionines. Molecules 2010, 15, 5581–5594. [Google Scholar] [CrossRef]
- Morrison, R.S.; Sharma, A.; de Vellis, J.; Bradshaw, R.A. Basic fibroblast growth factor supports the survival of cerebral cortical neurons in primary culture. Proc. Natl. Acad. Sci. USA 1986, 83, 7537. [Google Scholar] [CrossRef]
- Taverner, T.; Karpievitch, Y.V.; Polpitiya, A.D.; Brown, J.N.; Dabney, A.R.; Anderson, G.A.; Smith, R.D. DanteR: An extensible R-based tool for quantitative analysis of -omics data. Bioinformatics 2012, 28, 2402–2406. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Kilroy, G.E.; Foster, S.J.; Wu, X.; Ruiz, J.; Sherwood, S.; Heifetz, A.; Ludlow, J.W.; Stricker, D.M.; Potiny, S.; Green, P.; et al. Cytokine profile of human adipose-derived stem cells: Expression of angiogenic, hematopoietic, and pro-inflammatory factors. J. Cell. Physiol. 2007, 212, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, D.; Ricci, G.; Dupre, S.; Pecci, L.; Costa, M.; Matarese, R.M.; Pensa, B.; Antonucci, A.; Solinas, S.P.; Fontana, M. Sulfur-containing cyclic ketimines and imino acids. A novel family of endogenous products in the search for a role. Eur. J. Biochem. 1991, 202, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnabé, G.F.; Schwindt, T.T.; Calcagnotto, M.E.; Motta, F.L.; Martinez, G., Jr.; de Oliveira, A.C.; Keim, L.M.N.; D’Almeida, V.; Mendez-Otero, R.; Mello, L.E. Chemically-Induced RAT Mesenchymal Stem Cells Adopt Molecular Properties of Neuronal-Like Cells but Do Not Have Basic Neuronal Functional Properties. PLoS ONE 2009, 4, e5222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardini, M.; Ricci, G.; Vesci, L.; Pecci, L.; Cavallini, D. Bovine brain ketimine reductase. Biochim. Biophys. Acta 1988, 957, 286–292. [Google Scholar] [CrossRef]
- Fontana, M.; Ricci, G.; Solinas, S.P.; Antonucci, A.; Serao, I.; Duprè, S.; Cavallini, D. [35S]Lanthionine ketimine binding to bovine brain membranes. Biochem. Biophys. Res. Commun. 1990, 171, 480–486. [Google Scholar] [CrossRef]
- Ricci, G.; Vesci, L.; Nardini, M.; Arduini, A.; Storto, S.; Rosato, N.; Cavallini, D. Detection of 2H-1,4-thiazine-5,6-dihydro-3,5-dicarboxylic acid (lanthionine ketimine) in the bovine brain by a fluorometric assay. Biochim. Biophys. Acta 1989, 990, 211–215. [Google Scholar] [CrossRef]
- Hensley, K.; Venkova, K.; Christov, A.; Gunning, W.; Park, J. Collapsin response mediator protein-2: An emerging pathologic feature and therapeutic target for neurodisease indications. Mol. Neurobiol. 2011, 43, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Solinas, S.P.; Pecci, L.; Montefoschi, G.; Cavallini, D. The reducing activity of S-aminoethylcysteine ketimine and similar sulfur-containing ketimines. Biochem. Biophys. Res. Commun. 1992, 183, 481–486. [Google Scholar] [CrossRef]
- Marcoux, F.W.; Goodrich, J.E.; Dominick, M.A. Ketamine prevents ischemic neuronal injury. Brain Res. 1988, 452, 329–335. [Google Scholar] [CrossRef]
- Nada, S.E.; Tulsulkar, J.; Raghavan, A.; Hensley, K.; Shah, Z.A. A derivative of the CRMP2 binding compound lanthionine ketimine provides neuroprotection in a mouse model of cerebral ischemia. Neurochem. Int. 2012, 61, 1357–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilby, C.R.; Bader, J.F.; Salfen, B.E.; Youngquist, R.S.; Murphy, C.N.; Garverick, H.A.; Crooker, B.A.; Lucy, M.C. Plasma GH, IGF-I, and conception rate in cattle treated with low doses of recombinant bovine GH. Theriogenology 1999, 51, 1285–1296. [Google Scholar] [CrossRef]
- Bikkavilli, R.K.; Malbon, C.C. Mitogen-activated protein kinases and Wnt/β-catenin signaling: Molecular conversations among signaling pathways. Commun. Integr. Biol. 2009, 2, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Sheen, V.L.; Feng, Y.; Graham, D.; Takafuta, T.; Shapiro, S.S.; Walsh, C.A. Filamin A and Filamin B are co-expressed within neurons during periods of neuronal migration and can physically interact. Hum. Mol. Genet. 2002, 11, 2845–2854. [Google Scholar] [CrossRef] [Green Version]
- Nagano, T.; Morikubo, S.; Sato, M. Filamin A and FILIP (Filamin A-Interacting Protein) Regulate Cell Polarity and Motility in Neocortical Subventricular and Intermediate Zones during Radial Migration. J. Neurosci. 2004, 24, 9648–9657. [Google Scholar] [CrossRef] [PubMed]
- Vadodaria, K.C.; Brakebusch, C.; Suter, U.; Jessberger, S. Stage-Specific Functions of the Small Rho GTPases Cdc42 and Rac1 for Adult Hippocampal Neurogenesis. J. Neurosci. 2013, 33, 1179–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gualdoni, S.; Albertinazzi, C.; Corbetta, S.; Valtorta, F.; de Curtis, I. Normal levels of Rac1 are important for dendritic but not axonal development in hippocampal neurons. Biol. Cell 2007, 99, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Jo, H.Y.; Kim, M.H.; Cha, Y.Y.; Choi, S.W.; Shim, J.H.; Kim, T.J.; Lee, K.Y. H2O2-dependent hyperoxidation of peroxiredoxin 6 (Prdx6) plays a role in cellular toxicity via up-regulation of iPLA2 activity. J. Biol. Chem. 2008, 283, 33563–33568. [Google Scholar] [CrossRef]
- Rhee, S.G.; Chae, H.Z.; Kim, K. Peroxiredoxins: A historical overview and speculative preview of novel mechanisms and emerging concepts in cell signaling. Free Radic. Biol. Med. 2005, 38, 1543–1552. [Google Scholar] [CrossRef]
- Simzar, S.; Ellyin, R.; Shau, H.; Sarafian, T. Contrasting Antioxidant and Cytotoxic Effects of Peroxiredoxin I and II in PC12 and NIH3T3 Cells. Neurochem. Res. 2000, 25, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, M.K.; Lekli, I.; Ray, D.; Yodoi, J.; Das, D.K. Redox regulation of cell survival by the thioredoxin superfamily: An implication of redox gene therapy in the heart. Antioxid. Redox Signal. 2009, 11, 2741–2758. [Google Scholar] [CrossRef] [PubMed]
- Perez Estrada, C.; Covacu, R.; Sankavaram, S.R.; Svensson, M.; Brundin, L. Oxidative stress increases neurogenesis and oligodendrogenesis in adult neural progenitor cells. Stem Cells Dev. 2014, 23, 2311–2327. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H.; Lim, H.J.; Yoon, S.; Seong, J.K.; Bae, D.S.; Rhee, S.G.; Bae, Y.S. Ahnak Protein Activates Protein Kinase C (PKC) through Dissociation of the PKC-Protein Phosphatase 2A Complex. J. Biol. Chem. 2008, 283, 6312–6320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amagai, M. A Mystery of AHNAK//Desmoyokin Still Goes On. J. Investig. Dermatol. 2004, 123, xiv–xv. [Google Scholar] [CrossRef]
- Thomson, S.; Petti, F.; Sujka-Kwok, I.; Mercado, P.; Bean, J.; Monaghan, M.; Seymour, S.; Argast, G.; Epstein, D.; Haley, J. A systems view of epithelial–mesenchymal transition signaling states. Clin. Exp. Metast. 2011, 28, 137–155. [Google Scholar] [CrossRef]
- Borgonovo, B.; Cocucci, E.; Racchetti, G.; Podini, P.; Bachi, A.; Meldolesi, J. Regulated exocytosis: A novel, widely expressed system. Nat. Cell Biol. 2002, 4, 955–962. [Google Scholar] [CrossRef]
- Benaud, C.; Gentil, B.J.; Assard, N.; Court, M.; Garin, J.; Delphin, C.; Baudier, J. AHNAK interaction with the annexin 2/S100A10 complex regulates cell membrane cytoarchitecture. J. Cell Biol. 2004, 164, 133–144. [Google Scholar] [CrossRef]
- Gerasimenko, J.V.; Flowerdew, S.E.; Voronina, S.G.; Sukhomlin, T.K.; Tepikin, A.V.; Petersen, O.H.; Gerasimenko, O.V. Bile Acids Induce Ca2+ Release from Both the Endoplasmic Reticulum and Acidic Intracellular Calcium Stores through Activation of Inositol Trisphosphate Receptors and Ryanodine Receptors. J. Biol. Chem. 2006, 281, 40154–40163. [Google Scholar] [CrossRef]
- Ritter, B.; Modregger, J.; Paulsson, M.; Plomann, M. PACSIN 2, a novel member of the PACSIN family of cytoplasmic adapter proteins. FEBS Lett. 1999, 454, 356–362. [Google Scholar]
- Lee, I.H.; You, J.O.; Ha, K.S.; Bae, D.S.; Suh, P.-G.; Rhee, S.G.; Bae, Y.S. AHNAK-mediated Activation of Phospholipase C-γ1 through Protein Kinase C. J. Biol. Chem. 2004, 279, 26645–26653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, J.; Sobue, K. Localization and characterization of gelsolin in nervous tissues: Gelsolin is specifically enriched in myelin-forming cells. J. Neurosci. 1994, 14, 1038–1052. [Google Scholar] [CrossRef] [PubMed]
- Harms, C.; Bosel, J.; Lautenschlager, M.; Harms, U.; Braun, J.S.; Hortnagl, H.; Dirnagl, U.; Kwiatkowski, D.J.; Fink, K.; Endres, M. Neuronal gelsolin prevents apoptosis by enhancing actin depolymerization. Mol. Cell. Neurosci. 2004, 25, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Carro, E. Gelsolin as therapeutic target in Alzheimer’s disease. Expert Opin. Ther. Targets 2010, 14, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Lynch, G.; Baudry, M. Brain spectrin, calpain and long-term changes in synaptic efficacy. Brain Res. Bull. 1987, 18, 809–815. [Google Scholar] [CrossRef]
- Goodman, S.R.; Zimmer, W.E.; Clark, M.B.; Zagon, I.S.; Barker, J.E.; Bloom, M.L. Brain spectrin: Of mice and men. Brain Res. Bull. 1995, 36, 593–606. [Google Scholar] [CrossRef]
- Pasquini, L.A.; Millet, V.; Hoyos, H.C.; Giannoni, J.P.; Croci, D.O.; Marder, M.; Liu, F.T.; Rabinovich, G.A.; Pasquini, J.M. Galectin-3 drives oligodendrocyte differentiation to control myelin integrity and function. Cell Death Differ. 2011, 18, 1746–1756. [Google Scholar] [CrossRef] [Green Version]
- Comte, I.; Kim, Y.; Young, C.C.; van der Harg, J.M.; Hockberger, P.; Bolam, P.J.; Poirier, F.; Szele, F.G. Galectin-3 maintains cell motility from the subventricular zone to the olfactory bulb. J. Cell Sci. 2011, 124, 2438–2447. [Google Scholar] [CrossRef] [Green Version]
- Pesheva, P.; Kuklinski, S.; Schmitz, B.; Probstmeier, R. Galectin-3 promotes neural cell adhesion and neurite growth. J. Neurosci. Res. 1998, 54, 639–654. [Google Scholar] [CrossRef]
- Deverman, B.E.; Patterson, P.H. Cytokines and CNS development. Neuron 2009, 64, 61–78. [Google Scholar] [CrossRef]
- Guo, C.J.; Douglas, S.D.; Lai, J.P.; Pleasure, D.E.; Li, Y.; Williams, M.; Bannerman, P.; Song, L.; Ho, W.Z. Interleukin-1beta stimulates macrophage inflammatory protein-1alpha and -1beta expression in human neuronal cells (NT2-N). J. Neurochem. 2003, 84, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Biber, K.; Zuurman, M.W.; Dijkstra, I.M.; Boddeke, H.W. Chemokines in the brain: Neuroimmunology and beyond. Curr. Opin. Pharmacol. 2002, 2, 63–68. [Google Scholar] [CrossRef]
- Pease, J. Tails of the unexpected—An atypical receptor for the chemokine RANTES/CCL5 expressed in brain. Br. J. Pharmacol. 2009, 149, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Chao, C.C.; Hegg, C.C.; Thayer, S.; Peterson, P.K. Morphine inhibits human microglial cell production of, and migration towards, RANTES. J. Psychopharmacol. 2000, 14, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Keswani, S.C.; Polley, M.; Pardo, C.A.; Griffin, J.W.; McArthur, J.C.; Hoke, A. Schwann cell chemokine receptors mediate HIV-1 gp120 toxicity to sensory neurons. Ann. Neurol. 2003, 54, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Dejda, A.; Sokolowska, P.; Nowak, J.Z. Neuroprotective potential of three neuropeptides PACAP, VIP and PHI. Pharmacol. Rep. 2005, 57, 307–320. [Google Scholar]
- Huang, Y.S.; Cheng, S.N.; Chueh, S.H.; Tsai, Y.L.; Liou, N.H.; Guo, Y.W.; Liao, M.H.; Shen, L.H.; Chen, C.C.; Liu, J.C.; et al. Effects of interleukin-15 on neuronal differentiation of neural stem cells. Brain Res. 2009, 1304, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Kramer, J.M.; Gaffen, S.L. Interleukin-17: A new paradigm in inflammation, autoimmunity, and therapy. J. Periodontol. 2007, 78, 1083–1093. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Confidence Cutoff | Proteins Detected | Proteins before Grouping | Distinct Peptides | Spectra Identified | % Total Spectra |
|---|---|---|---|---|---|
| >2.0 (99) | 2108 | 2774 | 35,891 | 176,734 | 67.3 |
| >1.3 (95) | 2430 | 3204 | 36,993 | 178,574 | 68.0 |
| >0.47 (66) | 2741 | 5246 | 38,381 | 180,467 | 68.8 |
| Cutoff applied: >0.05 (10%) | 3491 | 15,271 | 41,011 | 184,078 | 70.1 |
| AECK Neural | Accession | Name | GO | Biological Process | Peptides (95%) | Fold Change | p-Value |
|---|---|---|---|---|---|---|---|
| O75369 | Filamin-B | GO:0030154 | cell differentiation | 129 | 1.230453968 | 2.44 × 10−15 | |
| P00367 | Glutamate dehydrogenase 1, mitochondrial | GO:0006537 | glutamate biosynthetic process | 12 | 1.339429975 | 0.01273619 | |
| P09429 | High mobility group protein B1 | GO:0031175 | neuron projection development | 16 | 1.15634203 | 0.03726548 | |
| P10599 | Thioredoxin | GO:0008283 | cell proliferation | 7 | 1.57030201 | 0.04286075 | |
| P15144 | Aminopeptidase N | GO:0030154 | cell differentiation | 32 | 1.223500967 | 1.10 × 10−5 | |
| P15559 | NAD(P)H dehydrogenase (quinone) 1 | GO:0007271 | synaptic transmission, cholinergic | 9 | 1.816967964 | 0.002394901 | |
| Q00610 | Clathrin heavy chain 1 | GO:0048011 | nerve growth factor receptor signalling pathway | 69 | 1.205008984 | 7.17 × 10−7 | |
| Q01082 | Spectrin beta chain, brain 1 | GO:0007411 | axon guidance | 54 | 1.284289956 | 3.63 × 10−10 | |
| Q06830 | Peroxiredoxin-1 | GO:0008283 | cell proliferation | 25 | 1.434638977 | 0.008896183 | |
| Q09666 | Transforming protein RhoA | GO:0007399 | nervous system development | 16 | 1.167513967 | 0.019935589 | |
| Q09666 | Neuroblast differentiation-associated protein AHNAK | GO:0007399 | nervous system development | 285 | 1.527696013 | 1.40 × 10−45 | |
| Q13813 | Spectrin alpha chain, brain | GO:0007411 | axon guidance | 71 | 1.29076004 | 6.27 × 10−14 | |
| P63000 | Ras-related C3 botulinum toxin substrate 1 | GO:0048011 | nerve growth factor receptor signaling pathway | 7 | 1.664183 | 0.006588 | |
| Q9P0L0 | Vesicle-associated membrane protein-associated protein A | GO:0031175 | neuron projection development | 10 | 1.441416979 | 0.006864889 |
| LK Neural | Accession | Name | GO | Biological Process | Peptides (95%) | Fold Change | p-Value |
|---|---|---|---|---|---|---|---|
| O75369 | Filamin-B | GO:0030154 | cell differentiation | 129 | 1.372761965 | 0.0149013 | |
| P06396 | Gelsolin | GO:0060271 | cilium morphogenesis | 23 | 1.32772994 | 0.001008915 | |
| P10599 | Thioredoxin | GO:0008283 | cell proliferation | 4 | 1.76563704 | 0.02659229 | |
| P11142 | Heat shock cognate 71 kDa protein | GO:0007269 | neurotransmitter secretion | 73 | 1.381860971 | 0.00224432 | |
| P11413 | Glucose-6-phosphate 1-dehydrogenase | GO:0001816 | cytokine production | 23 | 1.242061019 | 0.00224432 | |
| P15144 | Aminopeptidase N | GO:0030154 | cell differentiation | 32 | 1.44699502 | 0.019935589 | |
| P17931 | Galectin-3 | GO:0030154 | cell differentiation | 7 | 1.757151961 | 0.006587825 | |
| Q01082 | Spectrin beta chain, brain 1 | GO:0007411 | axon guidance | 54 | 1.506716013 | 6.27 × 10−14 | |
| Q09666 | Neuroblast differentiation-associated protein AHNAK | GO:0007399 | nervous system development | 285 | 1.659075022 | 2.00 × 10−25 | |
| Q13813 | Spectrin alpha chain, brain | GO:0007411 | axon guidance | 71 | 1.395959973 | 6.27 × 10−14 | |
| Q92974 | Rho guanine nucleotide exchange factor 2 | GO:0048011 | nerve growth factor receptor signaling pathway | 9 | 1.163854957 | 0.033079 | |
| P63000 | Ras-related C3 botulinum toxin substrate 1 | GO:0048011 | nerve growth factor receptor signaling pathway | 7 | 1.616315960 | 0.006153737 | |
| Q9P0L0 | Vesicle-associated membrane protein-associated protein A | GO:0031175 | neuron projection development | 10 | 1.531931043 | 0.039667 |
| AECK Stress | Accession | Name | GO | Biological Process | Peptides (95%) | Fold Change | p-Value |
|---|---|---|---|---|---|---|---|
| P01892 | HLA class I histocompatibility antigen, A-2 alpha chain | GO:0060333 | interferon-gamma-mediated signaling pathway | 4 | 1.4692 | 0.01600371 | |
| P04083 | Annexin A1 | GO:0006954 | inflammatory response | 45 | 1.3066 | 0.00108737 | |
| P04264 | Keratin, type II cytoskeletal 1 | GO:0006979 | response to oxidative stress | 13 | 2.0287 | 0.00100892 | |
| P06396 | Gelsolin | GO:0006921 | cellular component disassembly involved in apoptosis | 25 | 1.3732 | 2.05 × 10−13 | |
| P09429 | High mobility group protein B1 | GO:0002437 | inflammatory response to antigenic stimulus positive regulation of apoptosis | 16 | 1.1563 | 0.00754318 | |
| P11413 | Glucose-6-phosphate 1-dehydrogenase | GO:0034599 | cellular response to oxidative stress | 23 | 1.2994 | 0.00129321 | |
| P15121 | Aldose reductase | GO:0006950 | response to stress | 9 | 1.3973 | 0.00414161 | |
| P16070 | CD44 antigen | GO:0060333 | interferon-gamma-mediated signaling pathway | 15 | 1.3484 | 8.89 × 10−5 | |
| P30044 | Peroxiredoxin-5, mitochondrial | GO:0034614 | cellular response to reactive oxygen species inflammatory response | 9 | 1.3299 | 0.00063648 | |
| P35611 | Alpha-adducin | GO:0006921 | cellular component disassembly involved in apoptosis | 12 | 1.5308 | 4.06 × 10−12 | |
| P51572 | B-cell receptor-associated protein 31 | GO:0006921 | cellular component disassembly involved in apoptosis | 7 | 1.4834 | 0.00252419 | |
| P61586 | Transforming protein RhoA | GO:0050772 | positive regulation of axonogenesis | 16 | 1.1675 | 0.00013793 | |
| P63000 | Ras-related C3 botulinum toxin substrate 1 | GO:0008624 | induction of apoptosis by extracellular signals | 7 | 1.6642 | 3.48 × 10−6 | |
| P63000 | Ras-related C3 botulinum toxin substrate 1 | GO:0006954 | inflammatory response | 7 | 1.6642 | 0.00016355 | |
| P63241 | Eukaryotic translation initiation factor 5A-1 | GO:0006917 | induction of apoptosis | 20 | 1.4873 | 6.36 × 10−12 | |
| Q02952 | A-kinase anchor protein 12 | GO:0030819 | positive regulation of cAMP biosynthetic process | 17 | 1.3261 | 0.0067851 | |
| Q03135 | Caveolin-1 | GO:0009267 | cellular response to starvation; inactivation of MAPK activity; positive regulation of calcium ion transport into cytosol; positive regulation of canonical Wnt receptor signaling pathway; response to hypoxia | 7 | 3.1097 | 0.02121297 | |
| Q13813 | Spectrin alpha chain | GO:0006921 | cellular component disassembly involved in apoptosis | 71 | 1.2908 | 0.00131438 | |
| Q15149 | Plectin | GO:0006921 | cellular component disassembly involved in apoptosis | 174 | 1.3019 | 2.20 × 10−9 | |
| Q9NR28 | Diablo homolog, mitochondrial | GO:0008625 | induction of apoptosis via death domain receptors | 6 | 1.2655 | 2.20 × 10−9 |
| LK Stress | Accession | Name | GO | Biological Process | Peptides (95%) | Fold Change | p-Value |
|---|---|---|---|---|---|---|---|
| P02545 | Prelamin-A/C | GO:0006921 | cellular component disassembly involved in apoptosis | 63 | 1.4487 | 3.04 × 10−9 | |
| P06396 | Gelsolin | GO:0006921 | cellular component disassembly involved in apoptosis | 23 | 1.3277 | 0.00031779 | |
| P08670 | Vimentin | GO:0006921 | cellular component disassembly involved in apoptosis | 199 | 1.2634 | 7.44 × 10−5 | |
| P11413 | Glucose-6-phosphate 1-dehydrogenase | GO:0034599 | cellular response to oxidative stress | 23 | 1.2421 | 0.00100042 | |
| P35611 | Alpha-adducin | GO:0006921 | cellular component disassembly involved in apoptosis | 12 | 1.5308 | 0.02401391 | |
| P36776 | Lon protease homolog, mitochondrial | GO:0034599 | cellular response to oxidative stress | 17 | 1.4223 | 0.0050889 | |
| P51572 | B-cell receptor-associated protein 31 | GO:0006921 | cellular component disassembly involved in apoptosis | 7 | 1.5205 | 0.00379271 | |
| Q03135 | Caveolin-1 | GO:0009267 | cellular response to starvation; inactivation of MAPK activity | 7 | 2.5188 | 0.00436605 | |
| Q13813 | Spectrin alpha chain, brain | GO:0006921 | cellular component disassembly involved in apoptosis | 71 | 1.396 | 9.24 × 10−15 | |
| Q15149 | Plectin | GO:0006921 | cellular component disassembly involved in apoptosis | 174 | 1.569 | 2.06 × 10−39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, J.; Milthorpe, B.K.; Padula, M.P. Proteomic Analysis of Cyclic Ketamine Compounds Ability to Induce Neural Differentiation in Human Adult Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 523. https://doi.org/10.3390/ijms20030523
Santos J, Milthorpe BK, Padula MP. Proteomic Analysis of Cyclic Ketamine Compounds Ability to Induce Neural Differentiation in Human Adult Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2019; 20(3):523. https://doi.org/10.3390/ijms20030523
Chicago/Turabian StyleSantos, Jerran, Bruce Kenneth Milthorpe, and Matthew Paul Padula. 2019. "Proteomic Analysis of Cyclic Ketamine Compounds Ability to Induce Neural Differentiation in Human Adult Mesenchymal Stem Cells" International Journal of Molecular Sciences 20, no. 3: 523. https://doi.org/10.3390/ijms20030523
APA StyleSantos, J., Milthorpe, B. K., & Padula, M. P. (2019). Proteomic Analysis of Cyclic Ketamine Compounds Ability to Induce Neural Differentiation in Human Adult Mesenchymal Stem Cells. International Journal of Molecular Sciences, 20(3), 523. https://doi.org/10.3390/ijms20030523