Involvement of the PD-1/PD-L1 Co-Inhibitory Pathway in the Pathogenesis of the Inflammatory Stage of Early-Onset Preeclampsia

 ,
,
Abstract
:1. Introduction
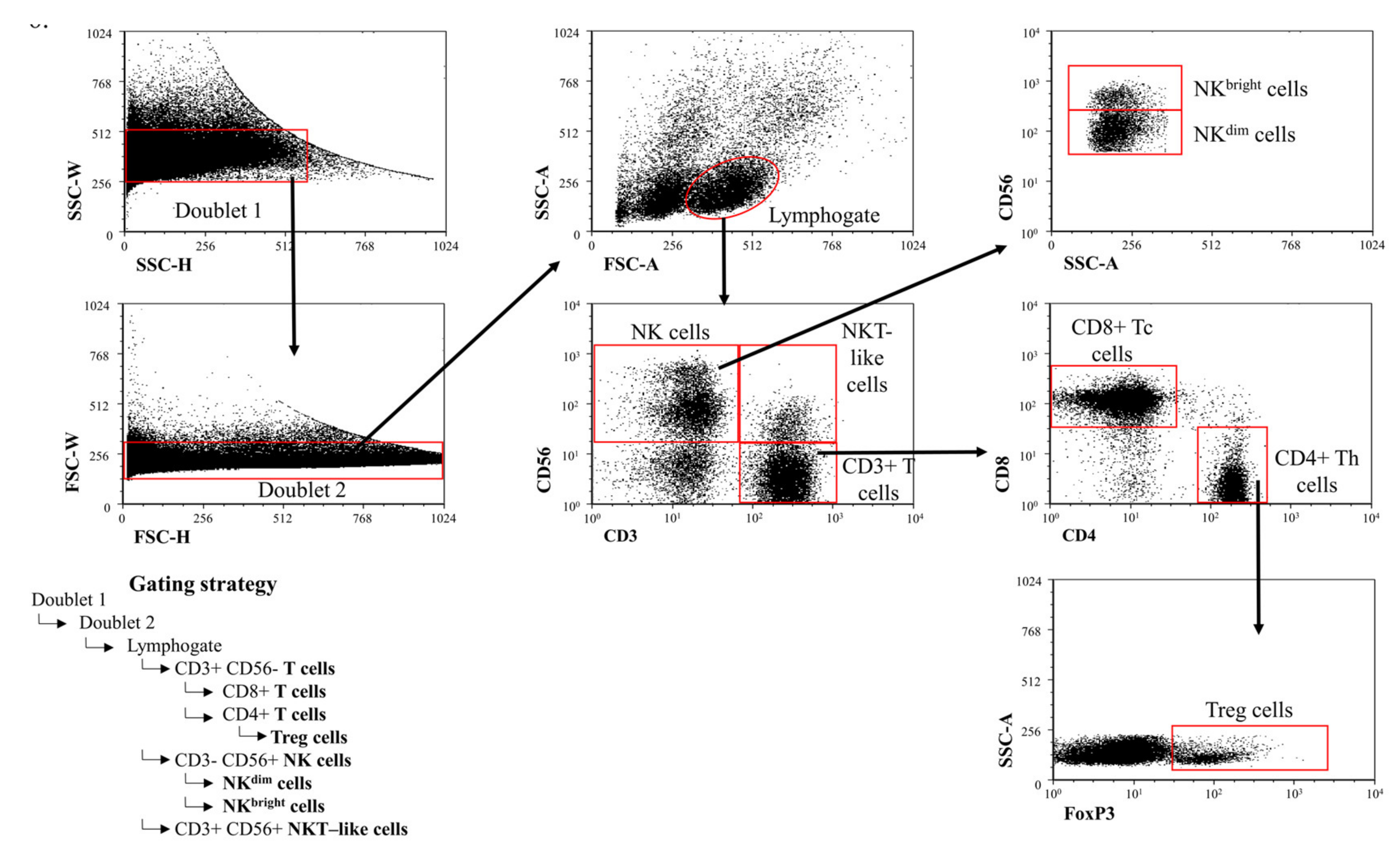
2. Results
2.1. The Ratio of Different Lymphocyte Subpopulations in the Peripheral Blood of 3rd-Trimester Healthy Pregnant Women and in Women with Early-Onset Preeclampsia
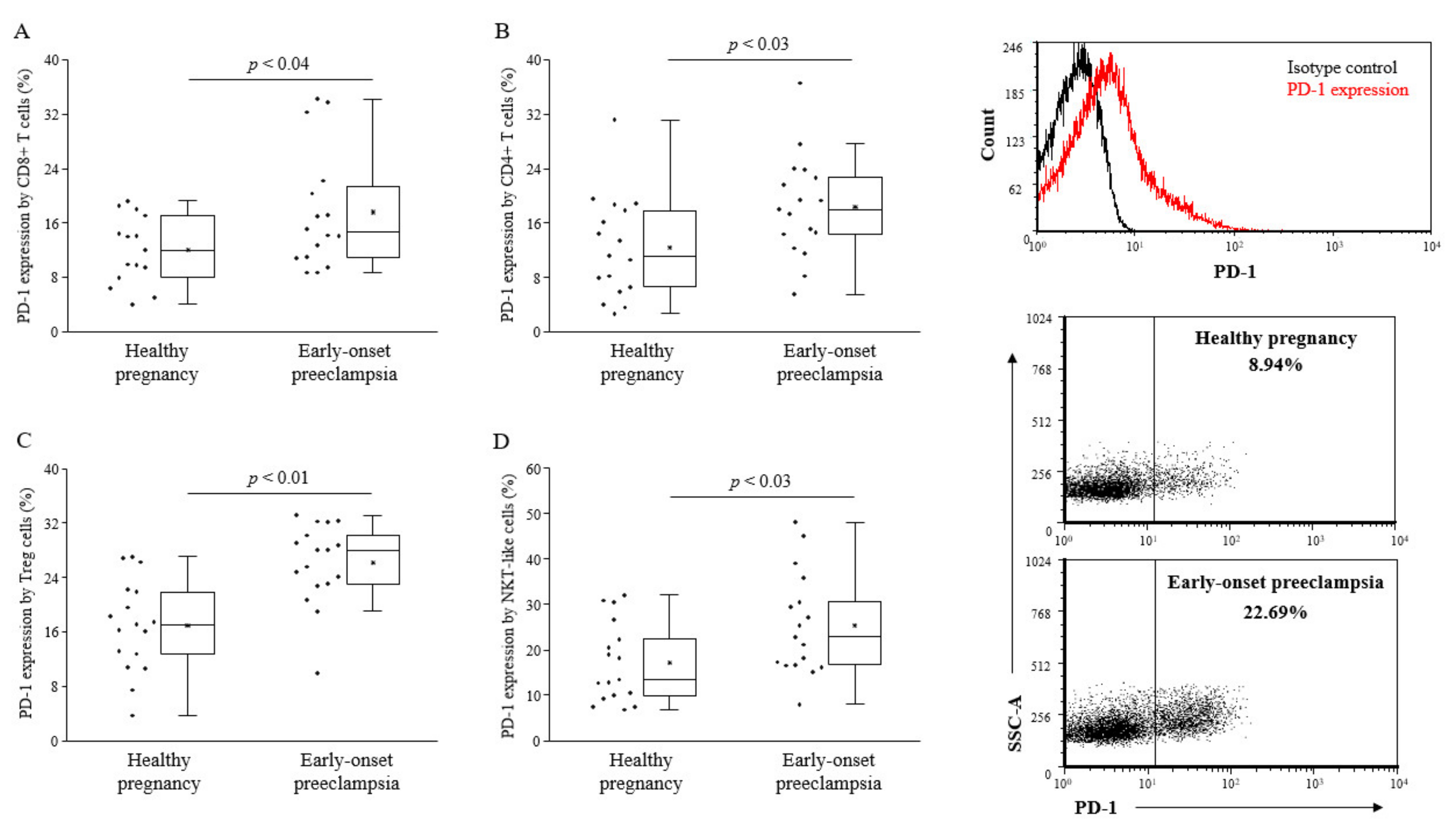
2.2. PD-1 Expression by Different Lymphocyte Subsets of the Innate and the Adaptive Immunity of 3rd-Trimester Healthy Pregnant Women and in Women with Early-Onset Preeclampsia
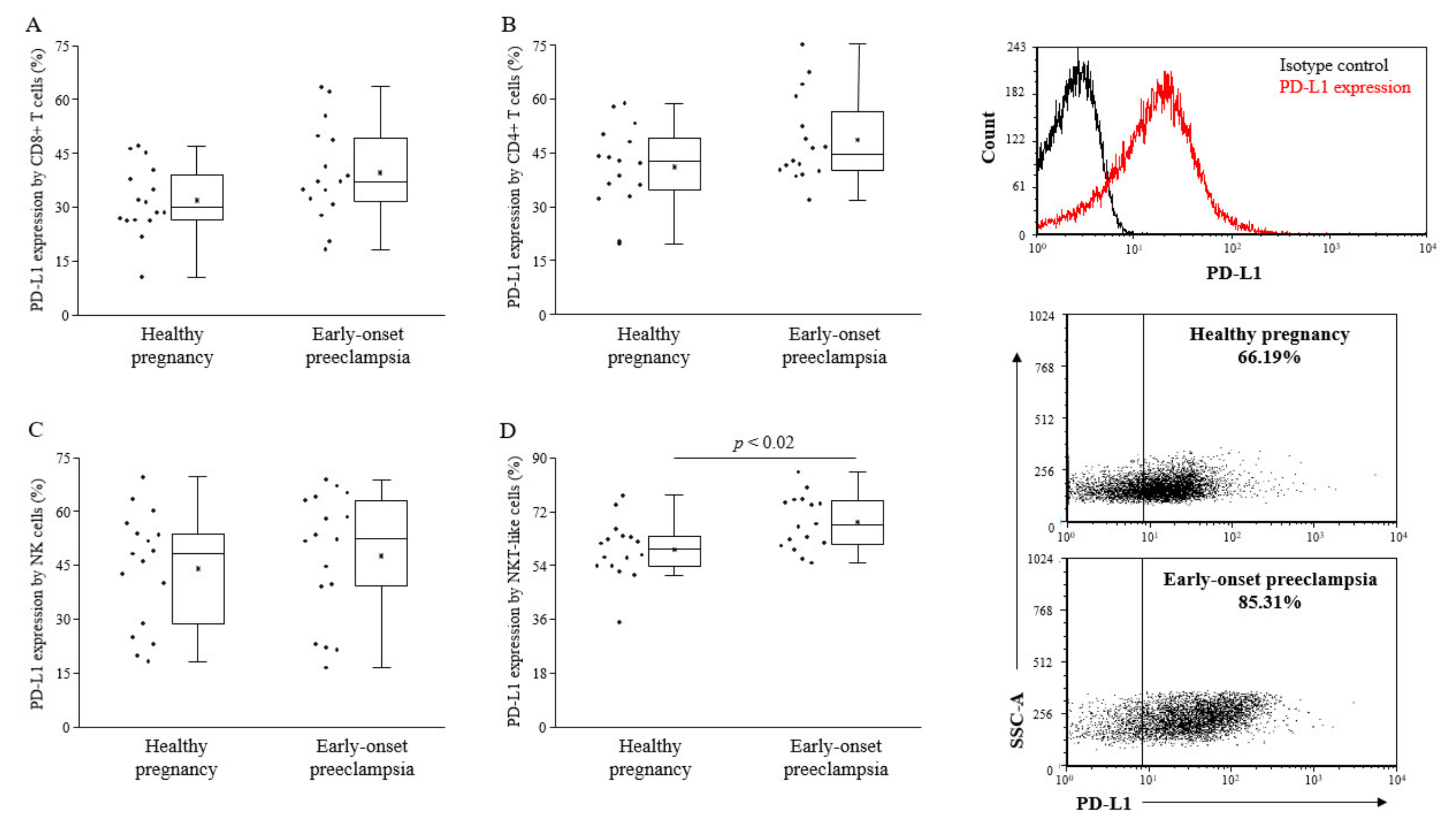
2.3. PD-L1 Expression by Different Lymphocyte Subsets of the Innate and the Adaptive Immunity of 3rd-Trimester Healthy Pregnant Women and in Women with Early-Onset Preeclampsia
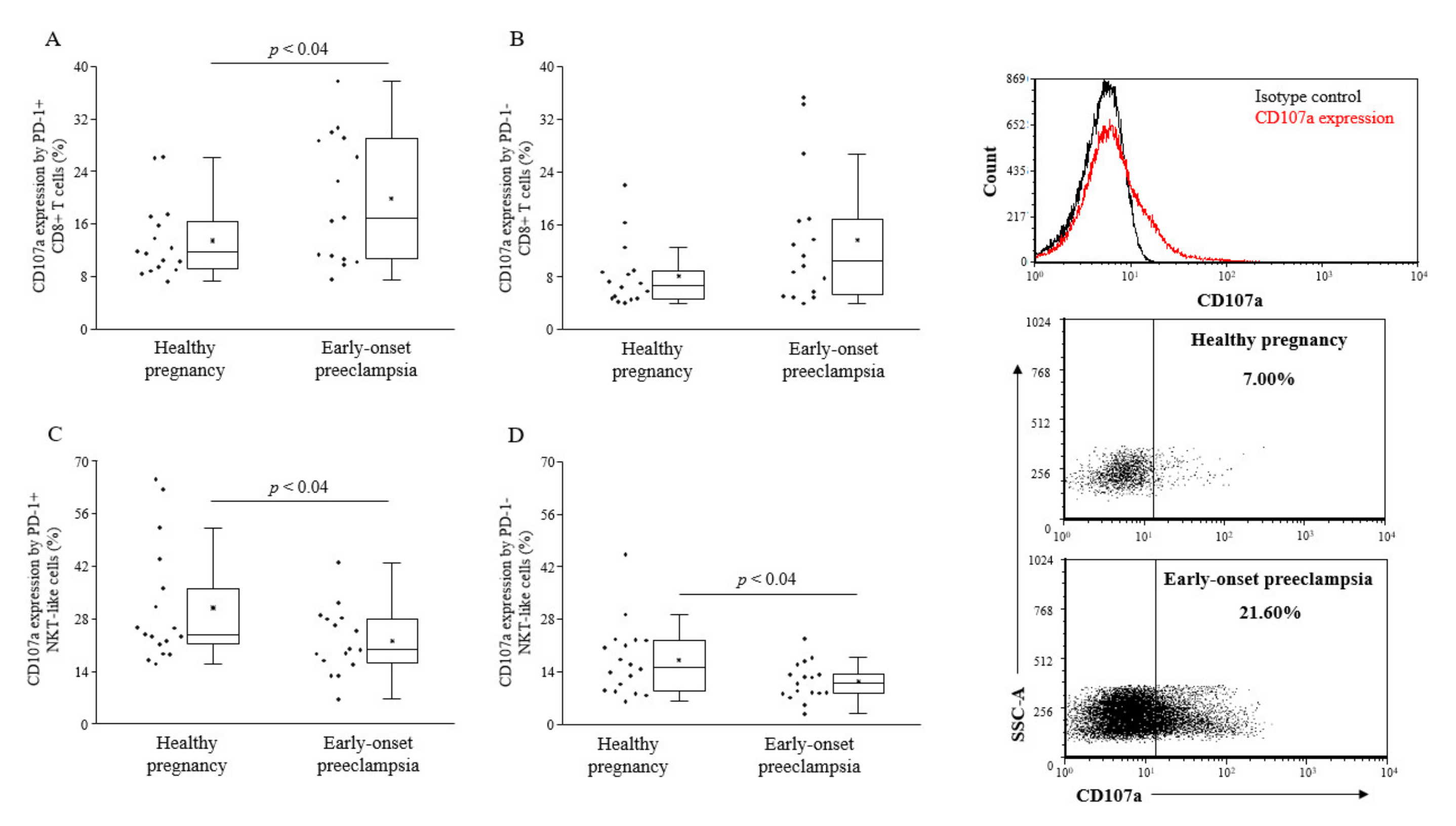
2.4. Cytotoxicity of PD-1 Expressing Lymphocytes in the Peripheral Blood of 3rd-Trimester Healthy Pregnant Women and Women in with Early-Onset Preeclampsia
2.5. PD-1 and NKG2D Co-Expression by Lymphocytes in the Peripheral Blood of 3rd-Trimester Healthy Pregnant Women and in Women with Early-Onset Preeclampsia
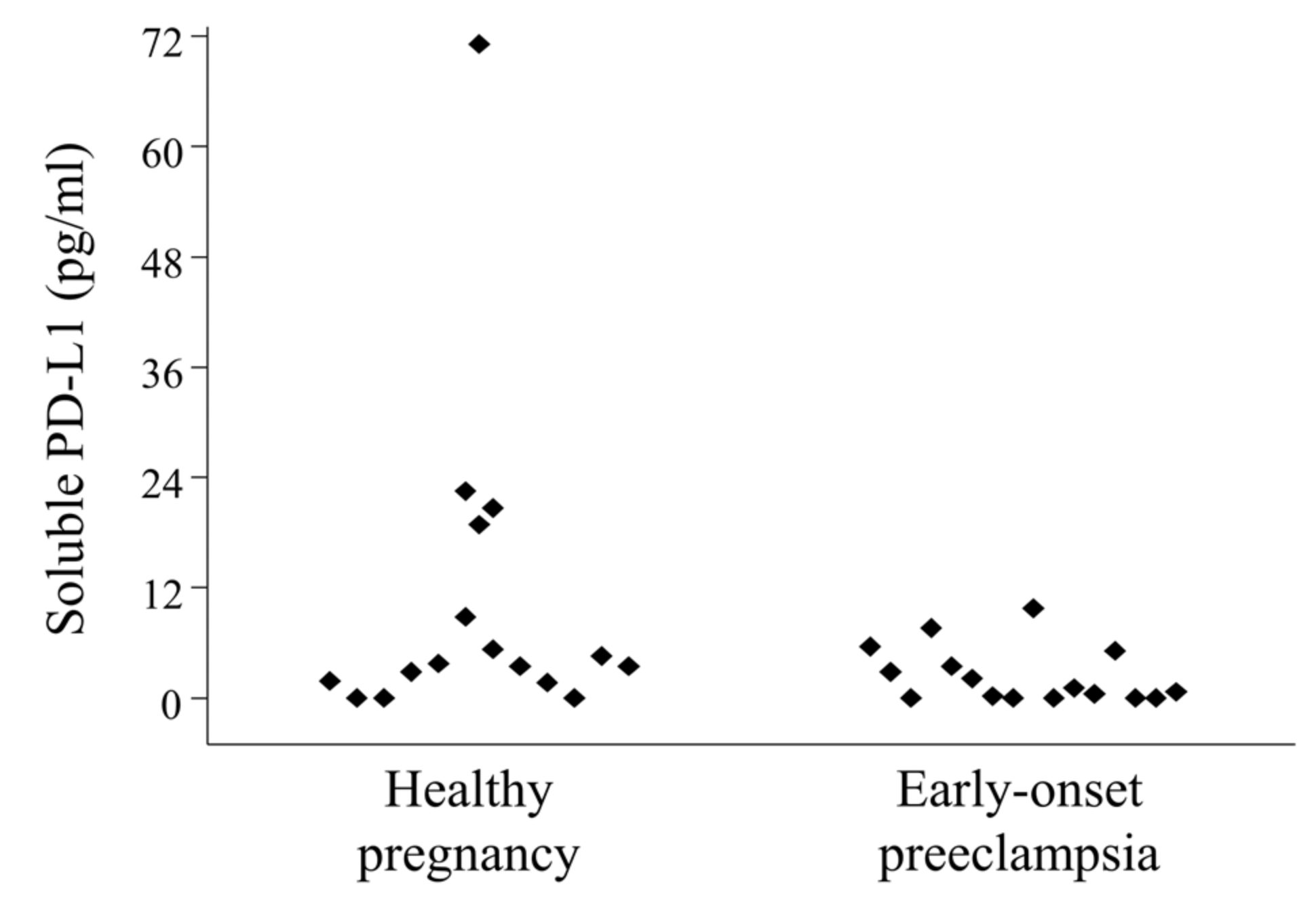
2.6. Serum-Soluble Programmed Death-Ligand 1 (sPD-L1) Levels in 3rd-Trimester Healthy Pregnant Women and in Women with Early-Onset Preeclampsia
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Lymphocyte Separation, Cryopreservation and Thawing
4.3. Antibodies
4.4. Labeling of Lymphocytes and Flow Cytometric Analysis
4.5. CD107a Functional Assay
4.6. FoxP3 Intracellular Labeling
4.7. Determination of Serum Levels of Soluble PD-L1
4.8. Statistical Analysis
4.9. Ethical Approval
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MAIT cell | Mucosal associated invariant T cell |
| NK cell | Natural Killer cell |
| NKT-like cell | Natural Killer T-like cell |
| PD-1 | Programmed death-1 receptor |
| PD-L1 | Programmed death-ligand 1 |
| sPD-L1 | Soluble programmed death-ligand 1 |
| Treg cell | Regulatory T cell |
References
- Arck, P.C.; Hecher, K. Fetomaternal immune cross-talk and its consequences for maternal and offspring’s health. Nat. Med. 2013, 99, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Shepard, M.T.; Bonney, E.A. PD-1 Regulates T cell proliferation in a tissue and subset-specific manner during normal mouse pregnancy. Immunol. Invest. 2013, 42, 385–408. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.K.; Johnson, J.G. Molecules involved in T-cell costimulation. Curr. Opin. Immunol. 1993, 5, 361–367. [Google Scholar] [CrossRef]
- Becker, T.C.; Wherry, E.J.; Boone, D.; Murali-Krishna, K.; Antia, R.; Ma, A.; Ahmed, R. Interleukin 15 is required for proliferative renewal of virus-specific memory CD8 T cells. J. Exp. Med. 2002, 195, 1541–1548. [Google Scholar] [CrossRef] [PubMed]
- Agata, Y.; Kawasaki, A.; Nishimura, H.; Ishida, Y.; Tsubata, T.; Yagita, H.; Honjo, T. Expression of the PD-1 antigen on the surface of stimulated mouse T and B lymphocytes. Int. Immunol. 1996, 8, 765–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riella, L.V.; Paterson, A.M.; Sharpe, A.H.; Chandraker, A. Role of the PD-1 pathway in the immune response. Am. J. Transplant. 2012, 12, 2575–2587. [Google Scholar] [CrossRef] [PubMed]
- Habicht, A.; Dada, S.; Jurewicz, M.; Fife, B.T.; Yagita, H.; Azuma, M.; Sayegh, M.H.; Guleria, I. A link between PDL1 and T regulatory cells in fetomaternal tolerance. J. Immunol. 2017, 179, 5211–5219. [Google Scholar] [CrossRef]
- Chikuma, S. Basics of PD-1 in self-tolerance, infection, and cancer immunity. Int. J. Clin. Oncol. 2016, 21, 448–455. [Google Scholar] [CrossRef]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef]
- Meggyes, M.; Szanto, J.; Lajko, A.; Farkas, B.; Varnagy, A.; Tamas, P.; Hantosi, E.; Miko, E.; Szereday, L. The possible role of CD8+/Vα7.2+/CD161++ T (MAIT) and CD8+/Vα7.2+/CD161 lo T (MAIT-like) cells in the pathogenesis of early-onset pre-eclampsi. Am. J. Reprod. Immunol. 2018, 79, e12805. [Google Scholar] [CrossRef]
- Petroff, M.G.; Perchellet, A. B7 family molecules as regulators of the maternal immune system in pregnancy. Am. J. Reprod. Immunol. 2010, 63, 506–519. [Google Scholar] [CrossRef] [PubMed]
- Petroff, M.G.; Chen, L.; Phillips, T.A.; Azzola, D.; Sedlmayr, P.; Hunt, J.S. B7 family molecules are favorably positioned at the human maternal-fetal interface. Biol. Reprod. 2003, 68, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Veras, E.; Kurman, R.J.; Wang, T.-L.; Shih, I.-M. PD-L1 expression in human placentas and gestational trophoblastic diseases. Int. J. Gynecol. Pathol. 2016, 36, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Sibai, B.; Dekker, G.; Kupferminc, M. Pre-eclampsia. Lancet 2005, 365, 785–799. [Google Scholar] [CrossRef]
- Raymond, D.; Peterson, E. A critical review of early-onset and late-onset preeclampsia. Obstet. Gynecol. Surv. 2011, 66, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Latest advances in understanding preeclampsia. Science 2005, 308, 1592–1594. [Google Scholar] [CrossRef] [PubMed]
- Borzychowski, A.M.; Sargent, I.L.; Redman, C.W.G. Inflammation and pre-eclampsia. Semin. Fetal Neonatal Med. 2006, 11, 309–316. [Google Scholar] [CrossRef]
- Miko, E.; Szereday, L.; Barakonyi, A.; Jarkovich, A.; Varga, P.; Szekeres-Bartho, J. The role of invariant NKT cells in pre-eclampsia. Am. J. Reprod. Immunol. 2008, 60, 118–126. [Google Scholar] [CrossRef]
- Miko, E.; Szereday, L.; Barakonyi, A.; Jarkovich, A.; Varga, P.; Szekeres-Bartho, J. Immunoactivation in preeclampsia: Vdelta2+ and regulatory T cells during the inflammatory stage of disease. J. Reprod. Immunol. 2009, 80, 100–108. [Google Scholar] [CrossRef]
- Molvarec, A.; Shiozaki, A.; Ito, M.; Toldi, G.; Stenczer, B.; Szarka, A.; Nakashima, A.; Vásárhelyi, B.; Rigó, J.; Saito, S. Increased prevalence of peripheral blood granulysin-producing cytotoxic T lymphocytes in preeclampsia. J. Reprod. Immunol. 2011, 91, 56–63. [Google Scholar] [CrossRef]
- Toldi, G.; Rigó, J.; Stenczer, B.; Vásárhelyi, B.; Molvarec, A. Increased prevalence of IL-17-producing peripheral blood lymphocytes in pre-eclampsia. Am. J. Reprod. Immunol. 2011, 66, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Molvarec, A.; Czegle, I.; Szijártó, J.; Rigó, J. Increased circulating interleukin-17 levels in preeclampsia. J. Reprod. Immunol. 2015, 112, 53–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latchman, Y.; Wood, C.R.; Chernova, T.; Chaudhary, D.; Borde, M.; Chernova, I.; Iwai, Y.; Long, A.J.; Brown, J.A.; Nunes, R.; et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat. Immunol. 2001, 2, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Darmochwal-Kolarz, D.; Suzuki, D.; Sakai, M.; Ito, M.; Shima, T.; Shiozaki, A.; Rolinski, J.; Saito, S. Proportion of peripheral blood and decidual CD4(+) CD25(bright) regulatory T cells in pre-eclampsia. Clin. Exp. Immunol. 2007, 149, 139–145. [Google Scholar] [CrossRef]
- Darmochwal-Kolarz, D.; Saito, S.; Rolinski, J.; Tabarkiewicz, J.; Kolarz, B.; Leszczynska-Gorzelak, B.; Oleszczuk, J. Activated T lymphocytes in pre-eclampsia. Am. J. Reprod. Immunol. 2007, 58, 39–45. [Google Scholar] [CrossRef]
- Prins, J.R.; Boelens, H.M.; Heimweg, J.; Van der Heide, S.; Dubois, A.E.; Van Oosterhout, A.J.; Erwich, J.J.H.M. Preeclampsia is associated with lower percentages of regulatory T cells in maternal blood. Hypertens. Pregnancy. 2009, 28, 300–311. [Google Scholar] [CrossRef]
- Steinborn, A.; Haensch, G.M.; Mahnke, K.; Schmitt, E.; Toermer, A.; Meuer, S.; Sohn, C. Distinct subsets of regulatory T cells during pregnancy: Is the imbalance of these subsets involved in the pathogenesis of preeclampsia? Clin. Immunol. 2008, 129, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Toldi, G.; Svec, P.; Vásárhelyi, B.; Mészáros, G.; Rigó, J.; Tulassay, T.; Treszl, A. Decreased number of FoxP3+ regulatory T cells in preeclampsia. Acta Obstet. Gynecol. Scand. 2008, 87, 1229–1233. [Google Scholar] [CrossRef]
- Toldi, G.; Vásárhelyi, Z.E.; Rigó, J.; Orbán, C.; Tamássy, Z.; Bajnok, A.; Shima, T.; Saito, S.; Molvarec, A. Prevalence of regulatory T-cell subtypes in preeclampsia. Am. J. Reprod. Immunol. 2015, 74, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Zhang, Y.; Liu, Z.; Sun, G.; Mor, G.; Liao, A. The PD-1/PD-L1 inhibitory pathway is altered in pre-eclampsia and regulates T cell responses in pre-eclamptic rats. Sci. Rep. 2016, 6, 27683. [Google Scholar] [CrossRef] [Green Version]
- Nagato, T.; Ohkuri, T.; Ohara, K.; Hirata, Y.; Kishibe, K.; Komabayashi, Y.; Ueda, S.; Takahara, M.; Kumai, T.; Ishibashi, K.; et al. Programmed death-ligand 1 and its soluble form are highly expressed in nasal natural killer/T-cell lymphoma: A potential rationale for immunotherapy. Cancer Immunol. Immunother. 2017, 66, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Xing, Y.; Zhang, Z.; Huang, J.; Chen, Y. Effect of soluble PD-L1 released by lung cancer cells in regulating the function of T lymphocytes. Zhonghua Zhong Liu Za Zhi 2013, 35, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Lang, J. Soluble PD-1 and PD-L1: Predictive and prognostic significance in cancer. Oncotarget 2017, 8, 97671–97682. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.; Chen, H.; Wang, W.; Chen, X.-Q.; Geng, Q.-R.; Xia, Z.-J.; Lu, Y. Serum levels of soluble programmed death ligand 1 predict treatment response and progression free survival in multiple myeloma. Oncotarget 2015, 6, 41228–41236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frigola, X.; Inman, B.A.; Lohse, C.M.; Krco, C.J.; Cheville, J.C.; Thompson, R.H.; Leibovich, B.; Blute, M.L.; Dong, H.; Kwon, E.D. Identification of a soluble form of B7-H1 that retains immunosuppressive activity and is associated with aggressive renal cell carcinoma. Clin. Cancer Res. 2011, 17, 1915–1923. [Google Scholar] [CrossRef] [PubMed]
- Rossille, D.; Gressier, M.; Damotte, D.; Maucort-Boulch, D.; Pangault, C.; Semana, G.; Le Gouill, S.; Haioun, C.; Tarte, K.; Lamy, T.; et al. Groupe ouest-est des leucémies et autres maladies du sang, groupe ouest-est des leucémies et autres maladies du sang. High level of soluble programmed cell death ligand 1 in blood impacts overall survival in aggressive diffuse large B-Cell lymphoma: Results from a French multicenter clinical trial. Leukemia 2014, 28, 2367–2375. [Google Scholar] [CrossRef] [PubMed]
- Finkelmeier, F.; Canli, Ö.; Tal, A.; Pleli, T.; Trojan, J.; Schmidt, M.; Kronenberger, B.; Zeuzem, S.; Piiper, A.; Greten, F.R.; et al. High levels of the soluble programmed death-ligand (sPD-L1) identify hepatocellular carcinoma patients with a poor prognosis. Eur. J. Cancer. 2016, 59, 152–159. [Google Scholar] [CrossRef]
- Zheng, Z.; Bu, Z.; Liu, X.; Zhang, L.; Li, Z.; Wu, A.; Wu, X.; Cheng, X.; Xing, X.; Du, H.; et al. Level of circulating PD-L1 expression in patients with advanced gastric cancer and its clinical implications. Chin. J. Cancer Res. 2014, 26, 104–111. [Google Scholar] [CrossRef]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lymphocyte Subpopulations | Healthy Pregnant Women (n = 17) | Early-Onset Preeclamptic Patients (n = 17) | p-Value |
|---|---|---|---|
| CD3+ T cells | 68.80 ± 9.91 | 65.01 ± 11.53 | NS |
| CD4+ T cells | 38.15 ± 10.77 | 35.76 ± 9.94 | NS |
| CD4+ T cells in CD3+ T cells | 60.12 ± 8.66 | 55.57 ± 9.04 | NS |
| CD8+ T cells | 26.36 ± 13.16 | 23.17 ± 6.79 | NS |
| CD8+ T cells in CD3+ T cells | 32.66 ± 8.60 | 34.46 ± 7.78 | NS |
| Treg cells | 1.95 ± 1.30 | 1.73 ± 0,53 | NS |
| NK cells | 12.08 ± 5.52 | 15.62 ± 9.16 | NS |
| NKdim cells | 10.37 ± 5.56 | 14.08 ± 8.59 | NS |
| NKbright cells | 1.79 ± 0.65 | 1.60 ± 1.12 | NS |
| NKT cells | 2.91 ± 2.79 | 2.80 ± 1.62 | NS |
| Parameter | Healthy Pregnant Women | Early-Onset Preeclamptic Patients |
|---|---|---|
| No. of patients | 17 | 17 |
| Age (years) | 32.29 (26–43) | 28.00 (17–42) |
| Gestational age at birth (week) | 39.14 ± 1.10 | 32.35 ± 3.46 * |
| Gestational age at sampling (week) | 33.29 ± 3.96 | 30.76 ± 2.41 |
| Birth weight (gram) | 3444.29 ± 512.38 | 1631.76 ± 738.56 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meggyes, M.; Miko, E.; Lajko, A.; Csiszar, B.; Sandor, B.; Matrai, P.; Tamas, P.; Szereday, L. Involvement of the PD-1/PD-L1 Co-Inhibitory Pathway in the Pathogenesis of the Inflammatory Stage of Early-Onset Preeclampsia. Int. J. Mol. Sci. 2019, 20, 583. https://doi.org/10.3390/ijms20030583
Meggyes M, Miko E, Lajko A, Csiszar B, Sandor B, Matrai P, Tamas P, Szereday L. Involvement of the PD-1/PD-L1 Co-Inhibitory Pathway in the Pathogenesis of the Inflammatory Stage of Early-Onset Preeclampsia. International Journal of Molecular Sciences. 2019; 20(3):583. https://doi.org/10.3390/ijms20030583
Chicago/Turabian StyleMeggyes, Matyas, Eva Miko, Adrienn Lajko, Beata Csiszar, Barbara Sandor, Peter Matrai, Peter Tamas, and Laszlo Szereday. 2019. "Involvement of the PD-1/PD-L1 Co-Inhibitory Pathway in the Pathogenesis of the Inflammatory Stage of Early-Onset Preeclampsia" International Journal of Molecular Sciences 20, no. 3: 583. https://doi.org/10.3390/ijms20030583
APA StyleMeggyes, M., Miko, E., Lajko, A., Csiszar, B., Sandor, B., Matrai, P., Tamas, P., & Szereday, L. (2019). Involvement of the PD-1/PD-L1 Co-Inhibitory Pathway in the Pathogenesis of the Inflammatory Stage of Early-Onset Preeclampsia. International Journal of Molecular Sciences, 20(3), 583. https://doi.org/10.3390/ijms20030583