Engineering of Fatty Acid Synthases (FASs) to Boost the Production of Medium-Chain Fatty Acids (MCFAs) in Mucor circinelloides

 , ,
, ,  and
and
Abstract
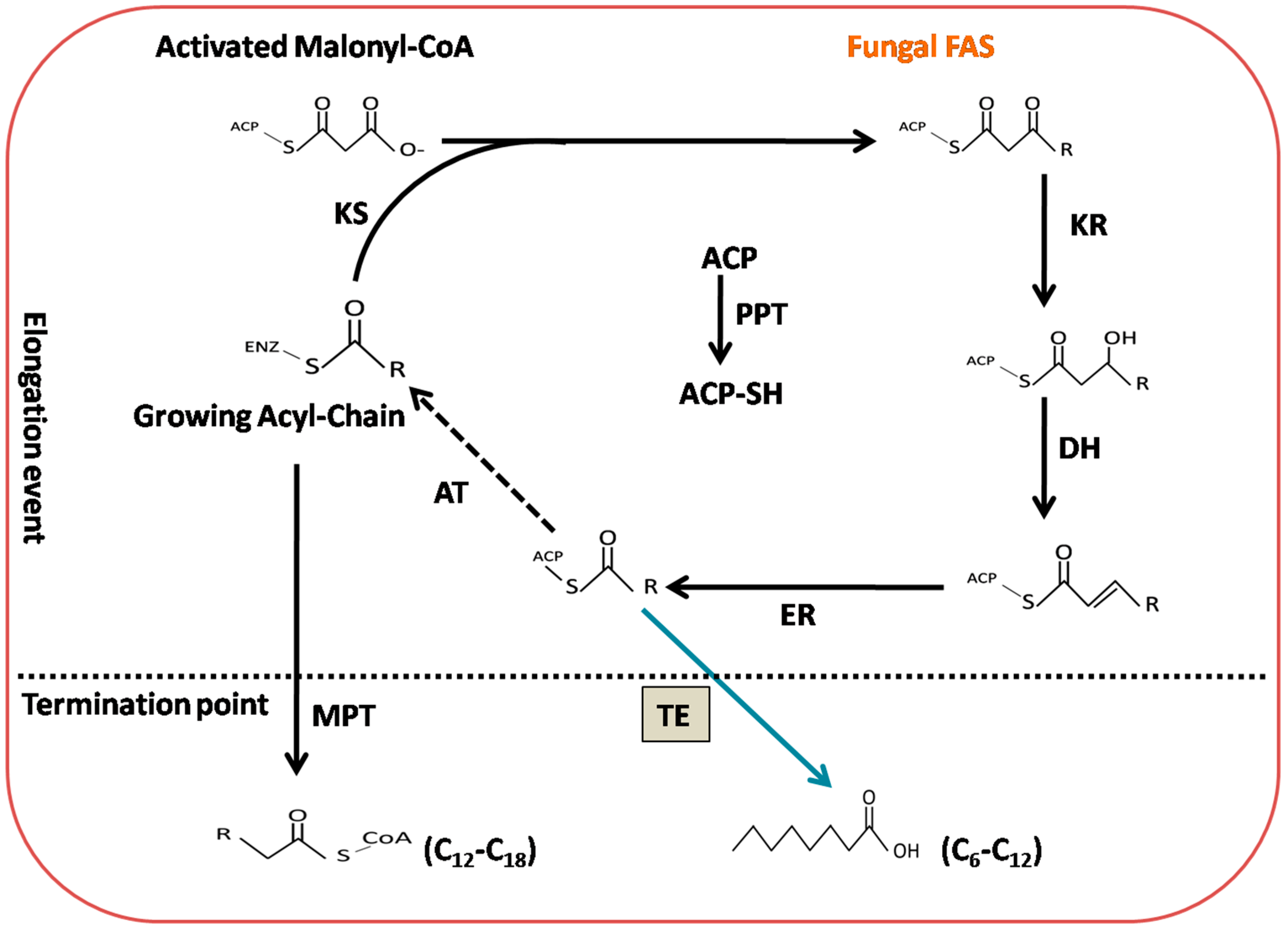
:1. Introduction
2. Results
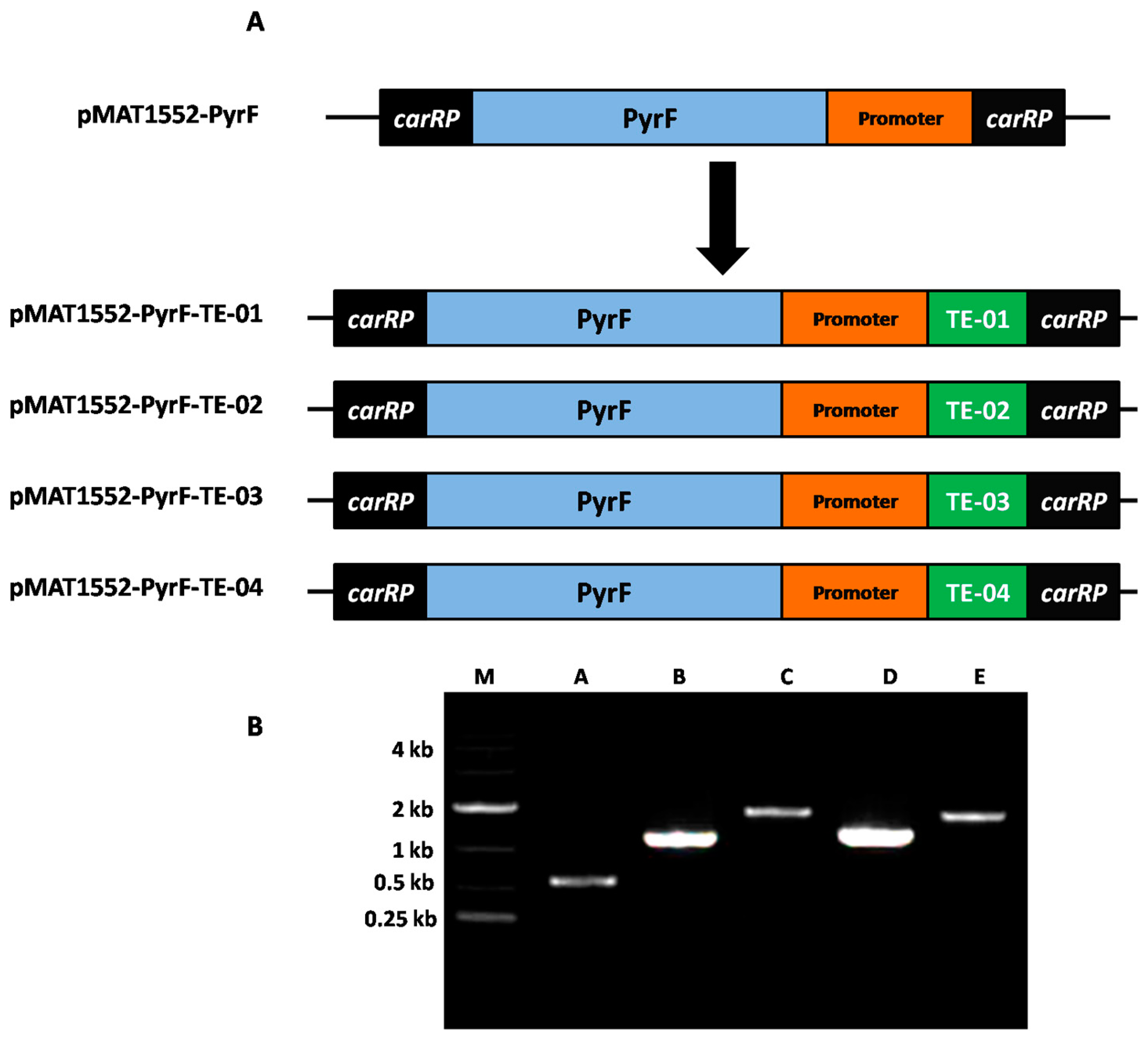
2.1. Generation of Diverse TE Gene-Over-Expressing Strains by Genetic Manipulation
2.2. Expression Levels of Different TE Genes in TE-Over-Expressing Strains
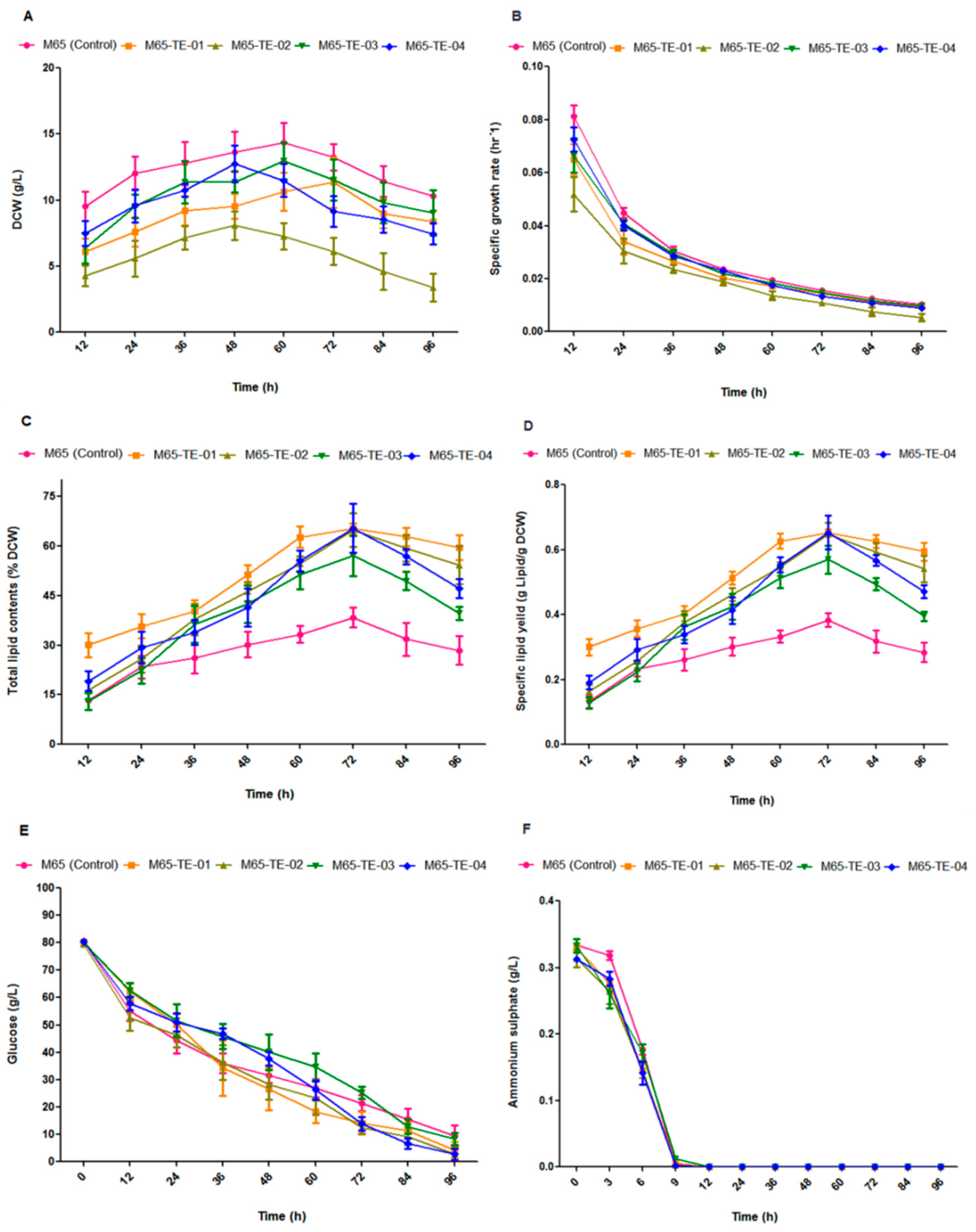
2.3. Cell Growth and Lipid Yield from the Engineered Fungal Strains
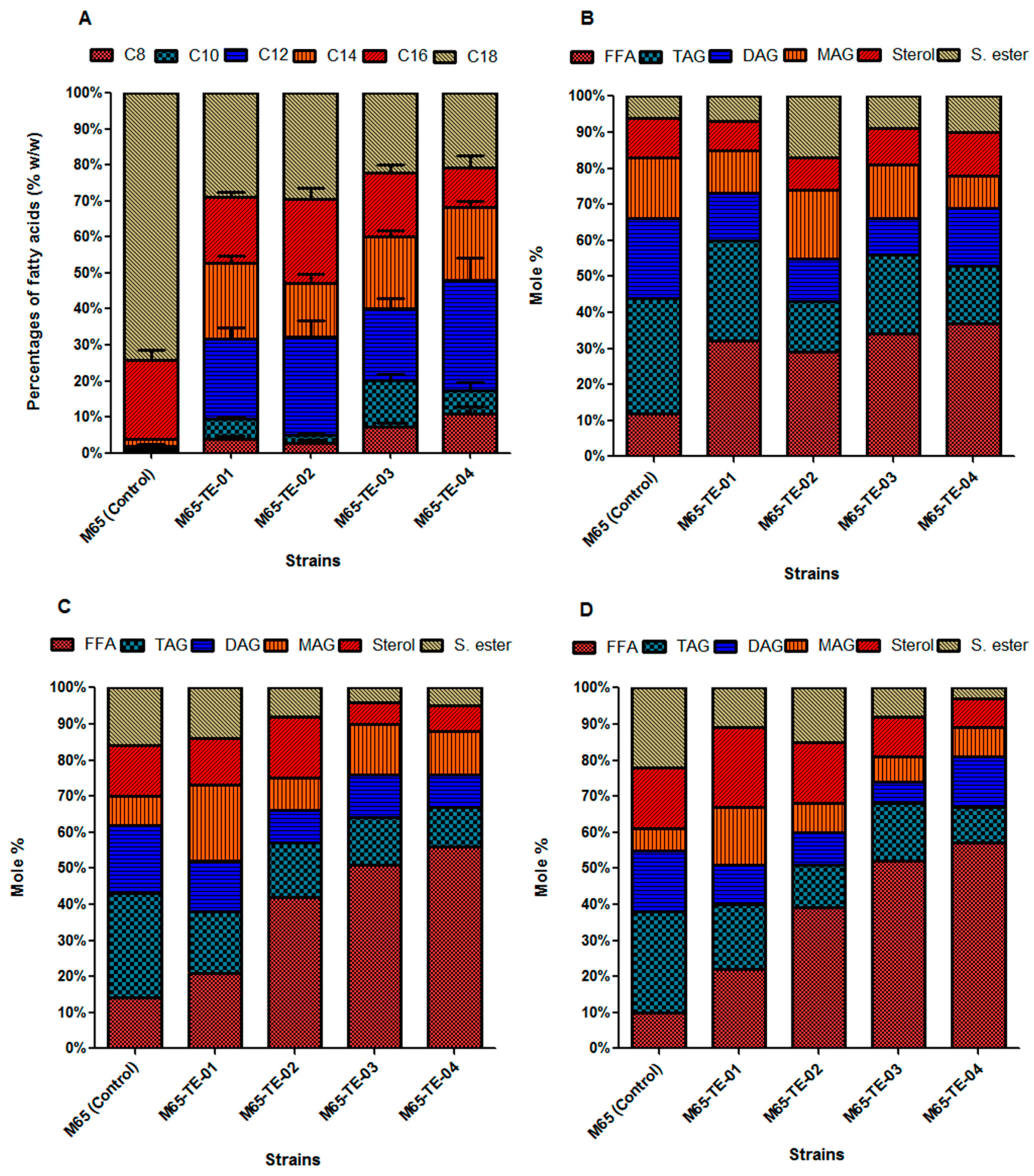
2.4. Production of De Novo Fatty Acids in M. Circinelloides M65
2.5. Integration of De Novo Medium-Chain Fatty Acids (MCFAs) into Diverse Lipid Classes
3. Discussion
4. Materials and Methods
4.1. Fungal Strains, Cultivation, and Transformation Conditions
4.2. Plasmids Construction
4.3. Preparation for Genomic DNA and Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
4.4. Determination of the Dry Cell Weight (DCW) and Lipid Accumulation
4.5. Determination of Glucose and Nitrogen Contents in the Culture Medium
4.6. Separation of Lipid Classes
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FAS | fatty acid synthase |
| MCFAs | medium-chain fatty acids |
| LCFAs | long chain fatty acids |
| DCW | dry cell weight |
| M.C | Mucor circinelloides |
| FFA | free fatty acid |
| SCO | single cell oil |
| TEs | thioesterases |
| TAG | triacylglycerol |
| DAG | diacylglycerol |
| MAG | monoacylglycerol |
| Tx | taxonomy |
| GC | gas chromatography |
| FAMEs | fatty acid methyl esters |
References
- Sarria, S.; Kruyer, N.S.; Yahya, P.P. Microbial synthesis of medium-chain chemicals from renewable. Nat. Biotechnol. 2017, 35, 1158–1166. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.; Verstrepen, K.J.; Dijck, P.V.; Thevelein, J.M.; Delvaux, F.R. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef]
- Knothe, G. “Designer” biodiesel: Optimizing fatty ester composition to improve fuel properties. Energy Fuels 2008, 22, 1358–1364. [Google Scholar] [CrossRef]
- Choi, Y.J.; Lee, S.Y. Microbial production of short-chain alkanes. Nature 2013, 502, 571–574. [Google Scholar] [CrossRef]
- Chang, Y.W.; Lee, D.; Bae, S.Y. Preparation of polyethylene-octene elastomer/clay nanocomposite and microcellular foam processed in supercritical carbon dioxide. Polym. Int. 2006, 55, 184–189. [Google Scholar] [CrossRef]
- Cabrales, L.; Calderon, K.; Hinojosa, I.; Valencia, F.; Abidi, N. Synthesis and characterization of polyesters derived from sebacic acid, hexanediol, and hydroquinone. Int. J. Polym. Anal. Charact. 2016, 21, 718–727. [Google Scholar] [CrossRef]
- Malca, S.H.; Scheps, D.; Kuhnel, L.; Venegas-Venegas, E.; Seifert, A.; Nestl, B.M.; Hauer, B. Bacterial CYP153A monooxygenases for the synthesis of omega-hydroxylated fatty acids. Chem. Commun. 2012, 48, 5115–5117. [Google Scholar]
- Liu, X.; Sheng, J.; Curtiss, R., III. Fatty acid production in genetically modified cyanobacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 6899–6904. [Google Scholar] [CrossRef] [Green Version]
- Nagao, K.; Yanagita, T. Medium-chain fatty acids: Functional lipids for the prevention and treatment of the metabolic syndrome. Pharm. Res. 2010, 61, 208–212. [Google Scholar] [CrossRef]
- Torella, J.P.; Ford, T.J.; Kim, S.N.; Chen, A.M.; Way, J.C.; Silver, P.A. Tailored fatty acid synthesis via dynamic control of fatty acid elongation. Proc. Natl. Acad. Sci. USA 2013, 110, 11290–11295. [Google Scholar] [CrossRef] [Green Version]
- Leber, C.; Da-Silva, N.A. Engineering of Saccharomyces cerevisiae for the synthesis of short chain fatty acids. Biotechnol. Bioeng. 2014, 111, 347–358. [Google Scholar] [CrossRef]
- Lian, J.; Zhao, H. Reversal of the β-oxidation cycle in Saccharomyces cerevisiae for production of fuels and chemicals. ACS Synth. Biol. 2015, 4, 332–341. [Google Scholar] [CrossRef]
- Xu, P.; Qiao, K.; Ahn, W.S.; Stephanopoulos, G. Engineering Yarrowia lipolytica as a platform for synthesis of drop-in transportation fuels and oleochemicals. Proc. Natl. Acad. Sci. USA 2016, 113, 10848–10853. [Google Scholar] [CrossRef]
- Dellomonaco, C.; Clomburg, J.M.; Miller, E.N.; Gonzalez, R. Engineered reversal of the β-oxidation cycle for the synthesis of fuels and chemicals. Nature 2011, 476, 355–3559. [Google Scholar] [CrossRef]
- Goh, E.B.; Baidoo, E.E.K.; Keasling, J.D.; Beller, H.R. Engineering of bacterial methyl ketone synthesis for biofuels. Appl. Environ. Microbiol. 2012, 78, 70–80. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Qin, J.; Siewers, V.; Nielsen, J. Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat. Commun. 2016, 7, 11709. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Clomburg, J.M.; Gonzalez, R. Synthesis of medium-chain length (C6–C10) fuels and chemicals via β-oxidation reversal in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2015, 42, 465–475. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhou, Y.J.; Krivoruchko, A.; Grininger, M.; Zhao, Z.K.; Nielsen, J. Expanding the product portfolio of fungal type I fatty acid synthases. Nat. Chem. Biol. 2017, 13, 360–362. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Villa, T.G. Oily yeasts as oleaginous cell factories. Appl. Microbiol. Biotechnol. 2011, 90, 1219–1227. [Google Scholar] [CrossRef]
- Beopoulos, A.; Chardo, T.; Nicaud, J.M. Yarrowia lipolytica: A model and a tool to understand the mechanisms implicated in lipid accumulation. Biochimie 2009, 91, 692–696. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part II: Technology and potential applications. Euro. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Huan, L.; Zhao, L.; Zan, X.; Song, Y.; Ratledge, C. Boosting fatty acid synthesis in Rhodococcus opacus PD630 by overexpression of autologous thioesterases. Biotechnol. Lett. 2016, 38, 999–1008. [Google Scholar] [CrossRef]
- Rigouin, C.; Croux, C.; Borsenberger, V.; Khaled, M.B.; Chardot, T.; Marty, A.; Bordes, F. Increasing medium chain fatty acids production in Yarrowia lipolytica by metabolic engineering. Microb. Cell Fact. 2018, 17, 142. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, J.; Chen, W.N. Engineering the Saccharomyces cerevisiae β-Oxidation Pathway to Increase Medium Chain Fatty Acid Production as Potential Biofuel. PLoS ONE 2014, 9, e84853. [Google Scholar] [CrossRef] [PubMed]
- Harwood, J.L. Fatty acid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 101–138. [Google Scholar] [CrossRef]
- Cronan, J.E.; Rock, C.O. Biosynthesis of Membrane Lipids Escherichia coli and Salmonella. In Cellular and Molecular Biology; ASM Press: Washington, DC, USA, 1996; pp. 612–636. [Google Scholar]
- Liu, X.; Hicks, W.M.; Silver, P.A.; Way, J.C. Engineering acyl carrier protein to enhance production of shortened fatty acids. Biotechnol. Biofuels 2016, 9, 24. [Google Scholar] [PubMed]
- Gajewski, J.; Pavlovic, R.; Fischer, M.; Boles, E.; Grininger, M. Engineering fungal de novo fatty acid synthesis for short chain fatty acid production. Nat. Commun. 2017, 8, 14650. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, T.; Xian, M.; Cao, Y.; Fang, F.; Zou, H. Fatty acid from the renewable sources: A promising feedstock for the production of biofuels and biobased chemicals. Biotechnol. Adv. 2014, 32, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Lynd, L.R.; Zyl, V.W.H.; McBride, J.E.; Laser, M. Consolidated bioprocessing of cellulosic biomass: An update. Curr. Opin. Biotechnol. 2005, 16, 577–583. [Google Scholar] [CrossRef]
- Runguphan, W.; Keasling, J.D. Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid-derived biofuels and chemicals. Metab. Eng. 2014, 21, 103–113. [Google Scholar] [CrossRef]
- McMahon, M.D.; Prather, K.L. Functional screening and in vitro analysis reveal thioesterases with enhanced substrate specificity profiles that improve short-chain fatty acid production in Escherichia coli. Appl. Environ. Microbiol. 2013, 80, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Voelker, T.A.; Jones, A.; Cranmer, A.M.; Davies, H.M.; Knutzon, D.S. Broad-range and binary-range acyl-acyl-carrier-protein thioesterases suggest an alternative mechanism for medium-chain production in seeds. Plant Physiol. 1997, 114, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Pollard, M.R.; Anderson, L.; Fan, C.; Hawkins, D.J.; Davies, H.M. A specific acyl-ACP thioesterase implicated in medium-chain fatty acid production in immature cotyledons of Umbellularia californica. Arch. Biochem. Biophys. 1991, 284, 306–312. [Google Scholar] [CrossRef]
- Rodríguez-Frómeta, R.A.; Gutiérrez, A.; Torres-Martínez, S.; Garre, V. Malic enzyme activity is not the only bottleneck for lipid accumulation in the oleaginous fungus Mucor circinelloides. Appl. Microbiol. Biotechnol. 2013, 97, 3063–3072. [Google Scholar] [CrossRef]
- Tang, X.; Chen, H.; Gu, Z.; Zhang, H.; Chen, Y.Q.; Song, Y.; Chen, W. Comparative Proteome Analysis between High Lipid-Producing Strain Mucor circinelloides WJ11 and Low Lipid-Producing Strain CBS 277.49. J. Agric. Food Chem. 2017, 65, 5074–5082. [Google Scholar] [CrossRef]
- Zhang, Y.; Luan, X.; Zhang, H.; Garre, V.; Song, Y.; Ratledge, C. Improved γ-linolenic acid production in Mucor circinelloides by homologous overexpressing of delta-12 and delta-6 desaturases. Microb. Cell Fact. 2017, 116, 113. [Google Scholar] [CrossRef]
- Hameed, A.; Hussain, S.A.; Yang, J.; Ijaz, M.U.; Liu, Q.; Suleria, H.A.R.; Song, Y. Antioxidants Potential of the Filamentous Fungi (Mucor circinelloides). Nutrients 2017, 9, 1101. [Google Scholar] [CrossRef]
- Kelder, B.; Mukeji, P.; Kirchner, S.; Hovanec, G.; Leonard, A.E.; Chuang, L.T.; Kopchick, J.J.; Huang, Y.S. Expression of fungal desaturase genes in cultured mammalian cells. Mol. Cell Biochem. 2001, 219, 7–11. [Google Scholar] [CrossRef]
- Michinaka, Y.; Aki, T.; Shimauchi, T.; Nakajima, T.; Kawamoto, S.; Shigeta, S.; Suzuki, O.; Ono, K. Differential response to low temperature of two Delta6 fatty acid desaturases from Mucor circinelloides. Appl. Microbiol. Biotechnol. 2003, 62, 362–368. [Google Scholar] [CrossRef]
- Carvalho, A.K.F.; Rivaldi, J.D.; Barbosa, J.C.; Castro, H.F.D. Biosynthesis, characterization and enzymatic transesterification of single cell oil of Mucor circinelloides-a sustainable pathway for biofuel production. Bioresour. Technol. 2015, 181, 47–53. [Google Scholar] [CrossRef]
- Carvalho, A.K.F.; Conceicao, L.R.V.D.; Silva, J.P.V.; Perez, V.H.; Castro, H.F.D. Biodiesel production from Mucor circinelloides using ethanol and heteropolyacid in one and two-step transesterification. Fuel 2017, 202, 503–511. [Google Scholar] [CrossRef]
- Wynn, J.P. Biochemical events leading to diversion of carbon into storage lipids in oleaginous fungi Mucor circinelloides and Mortierella alpine. Microbiology 2001, 147, 2857–2864. [Google Scholar] [CrossRef] [PubMed]
- Salunke, D.; Mangalekar, R.; Kuvalekar, A.; Harsulkar, A. Bioconversion of alpha-linolenic acid into long chain polyunsaturated fatty acids by oleaginous fungi. Int. J. Phar. Biol. Sci. 2014, 5, 27–35. [Google Scholar]
- Sherkhanov, S.; Korman, T.P.; Bowie, J.U. Improving the tolerance of Escherichia coli to medium-chain fatty acid production. Metab. Eng. 2014, 25, 1–7. [Google Scholar] [CrossRef]
- Ruter, C.D. Engineering Yarrowia lipolytica for production of medium-chain fatty acids. Appl. Microbiol. Biotechnol. 2015, 99, 7359–7368. [Google Scholar] [CrossRef] [PubMed]
- Stefan, A.; Hochkoeppler, A.; Ugolini, L.; Lazzeri, L.; Conte, E. The Expression of the Cuphea Palustris Thioesterase CpFatB2 in Yarrowia Lipolytica Triggers Oleic Acid Accumulation. Biotechnol. Prog. 2016, 32, 26–35. [Google Scholar] [CrossRef]
- Lomakin, I.B.; Xiong, Y.; Steitz, T.A. The crystal structure of yeast fatty acid synthase, a cellular machine with eight active sites working together. Cell 2007, 129, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Leibundgut, M.; Maier, T.; Jenni, S.; Ban, N. The multienzyme architecture of eukaryotic fatty acid synthases. Curr. Opin. Struct. Biol. 2008, 18, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Cronan, J. Inhibition of fatty acid synthesis in Escherichia coli in the absence of phospholipid synthesis and release of inhibition by thioesterase action. J. Bacteriol. 1994, 176, 2814–2821. [Google Scholar] [CrossRef] [PubMed]
- Ohlrogge, J.B.; Jaworski, J.G. Regulation of fatty acid synthesis. Annu. Rev. Plant Biol. 1997, 48, 109–136. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–563. [Google Scholar] [CrossRef]
- Nicolás, F.E.; de Haro, J.P.; Torres-Martínez, S.; Ruiz-Vázquez, R.M. Mutants defective in a Mucor circinelloides dicer-like gene are not compromised in siRNA silencing but display developmental defects. Fung. Genet. Biol. 2007, 44, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Bartnicki-García, S.; Nickerson, W.J. Nutrition, growth and morphogenesis of Mucor rouxii. J. Bacteriol. 1962, 84, 841–858. [Google Scholar] [PubMed]
- Torres-Martínez, S.; Ruiz-Vázquez, R.M.; Garre, V.; López-García, S.; Navarro, E.; Vila, A. Molecular tools for carotenogenesis analysis in the zygomycete Mucor circinelloides. Methods Mol. Biol. 2012, 898, 85–107. [Google Scholar]
- Vellanki, S.; Navarro-Mendoza, M.I.; Garcia, A.E.; Murcia, L.; Perez-Arques, C.; Garre, V.; Nicolas, F.E.; Lee, S.C. Mucor circinelloides: Growth, maintenance, and genetic manipulation. Curr. Protocol. Microbiol. 2018, 49, e53. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, A.; Ratledge, C. Desaturation of polyunsaturated fatty acids in Mucor circinelloides and the involvement of a novel membrane-bound malic enzyme. Eur. J. Biochem. 1992, 209, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar]
- Carroll, K.K. Separation of lipid classes by chromatography on Florisil. J. Lipid Res. 1961, 2, 135–141. [Google Scholar]
- Freeman, C.; West, D. Complete separation of lipid classes on a single thin-layer plate. J. Lipid Res. 1966, 7, 324–327. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M65 (Control) | M65-TE-01 | M65-TE-02 | M65-TE-03 | M65-TE-04 | |
|---|---|---|---|---|---|
| DCW (g/L) | 13.25 ± 0.11 | 11.34 ± 0.31 | 06.12 ± 1.12 | 11.54 ± 0.82 | 09.14 ± 0.45 |
| % TLC (DCW) | 38.31 ± 0.19 | 65.32 ± 0.43 | 64.80 ± 1.91 | 57.01 ± 1.23 | 65.23 ± 1.42 |
| TLC (g/L) | 05.07 ± 0.11 | 07.40 ± 0.09 | 03.96 ± 0.07 | 06.57 ± 0.04 | 05.96 ± 0.73 |
| % MCFA | 02.33 ± 0.91 | 30.52 ± 1.14 | 28.87 ± 1.34 | 39.37 ± 0.79 | 46.55 ± 0.45 |
| MCFA(g/L) | 0.11 ± 0.04 | 02.25 ± 0.05 | 01.14 ± 0.02 | 02.58 ± 0.04 | 02.77 ± 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, S.A.; Hameed, A.; Khan, M.A.K.; Zhang, Y.; Zhang, H.; Garre, V.; Song, Y. Engineering of Fatty Acid Synthases (FASs) to Boost the Production of Medium-Chain Fatty Acids (MCFAs) in Mucor circinelloides. Int. J. Mol. Sci. 2019, 20, 786. https://doi.org/10.3390/ijms20030786
Hussain SA, Hameed A, Khan MAK, Zhang Y, Zhang H, Garre V, Song Y. Engineering of Fatty Acid Synthases (FASs) to Boost the Production of Medium-Chain Fatty Acids (MCFAs) in Mucor circinelloides. International Journal of Molecular Sciences. 2019; 20(3):786. https://doi.org/10.3390/ijms20030786
Chicago/Turabian StyleHussain, Syed Ammar, Ahsan Hameed, Md. Ahsanul Kabir Khan, Yao Zhang, Huaiyuan Zhang, Victoriano Garre, and Yuanda Song. 2019. "Engineering of Fatty Acid Synthases (FASs) to Boost the Production of Medium-Chain Fatty Acids (MCFAs) in Mucor circinelloides" International Journal of Molecular Sciences 20, no. 3: 786. https://doi.org/10.3390/ijms20030786
APA StyleHussain, S. A., Hameed, A., Khan, M. A. K., Zhang, Y., Zhang, H., Garre, V., & Song, Y. (2019). Engineering of Fatty Acid Synthases (FASs) to Boost the Production of Medium-Chain Fatty Acids (MCFAs) in Mucor circinelloides. International Journal of Molecular Sciences, 20(3), 786. https://doi.org/10.3390/ijms20030786