Overexpression of Transglutaminase from Cucumber in Tobacco Increases Salt Tolerance through Regulation of Photosynthesis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
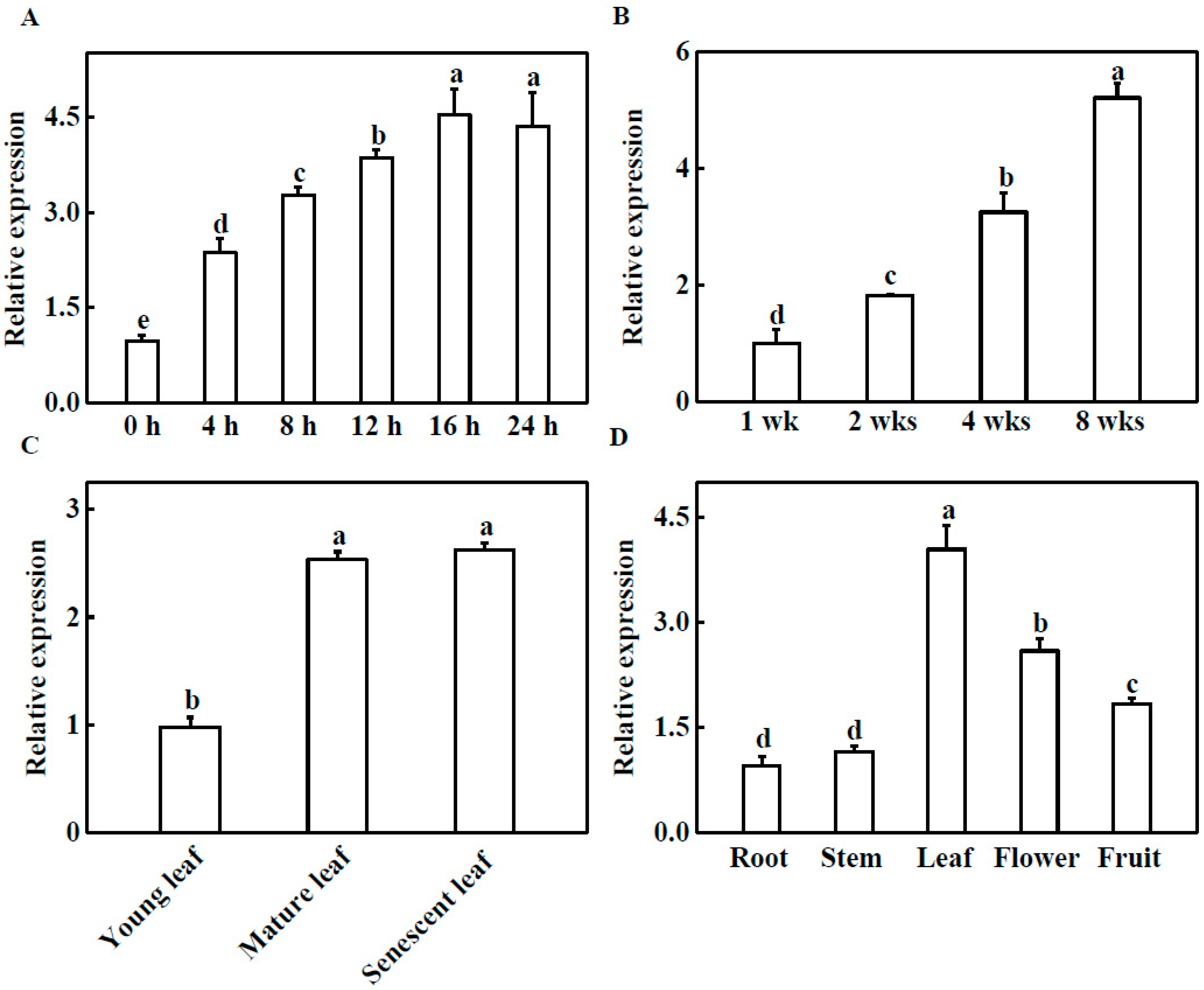
2.1. Expression Profile Analysis of TGase
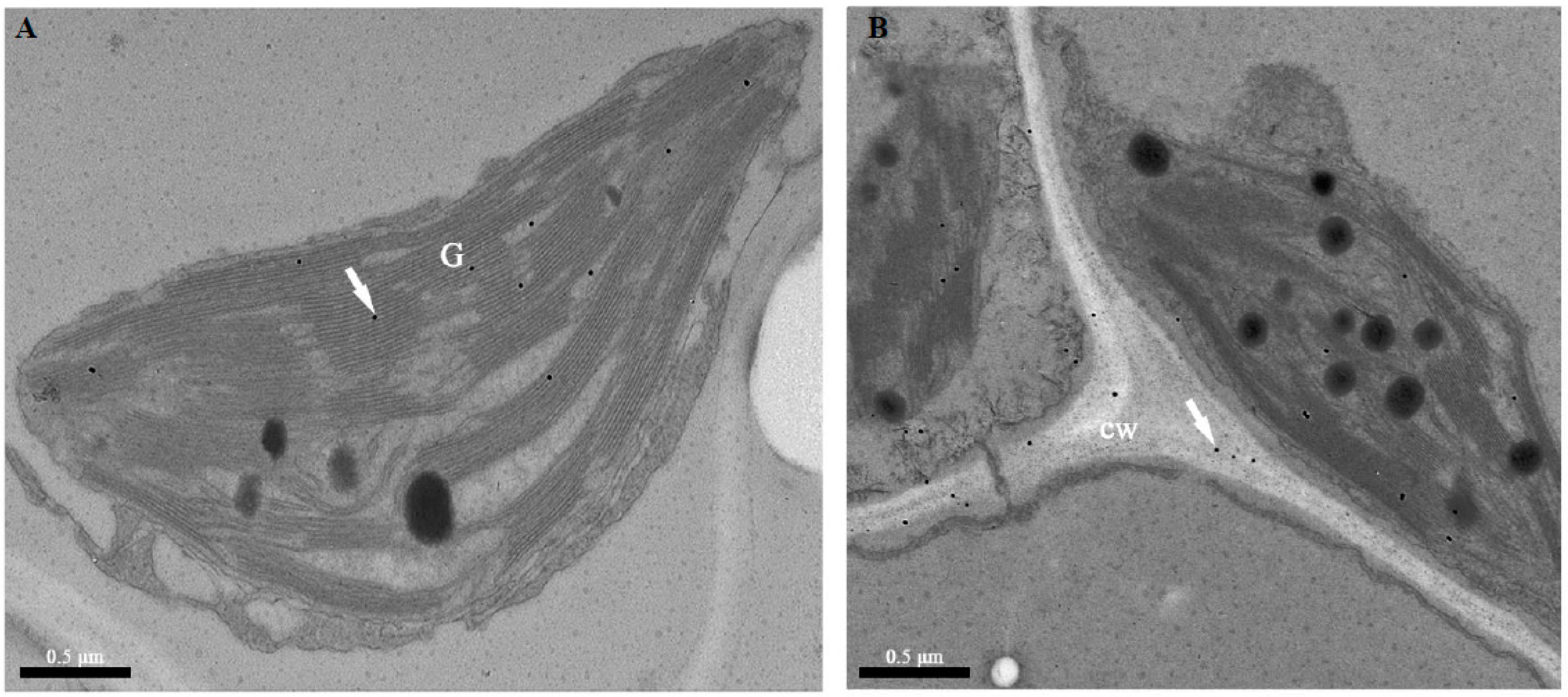
2.2. Immunolocalization of TGase Protein in Cucumber Leaves
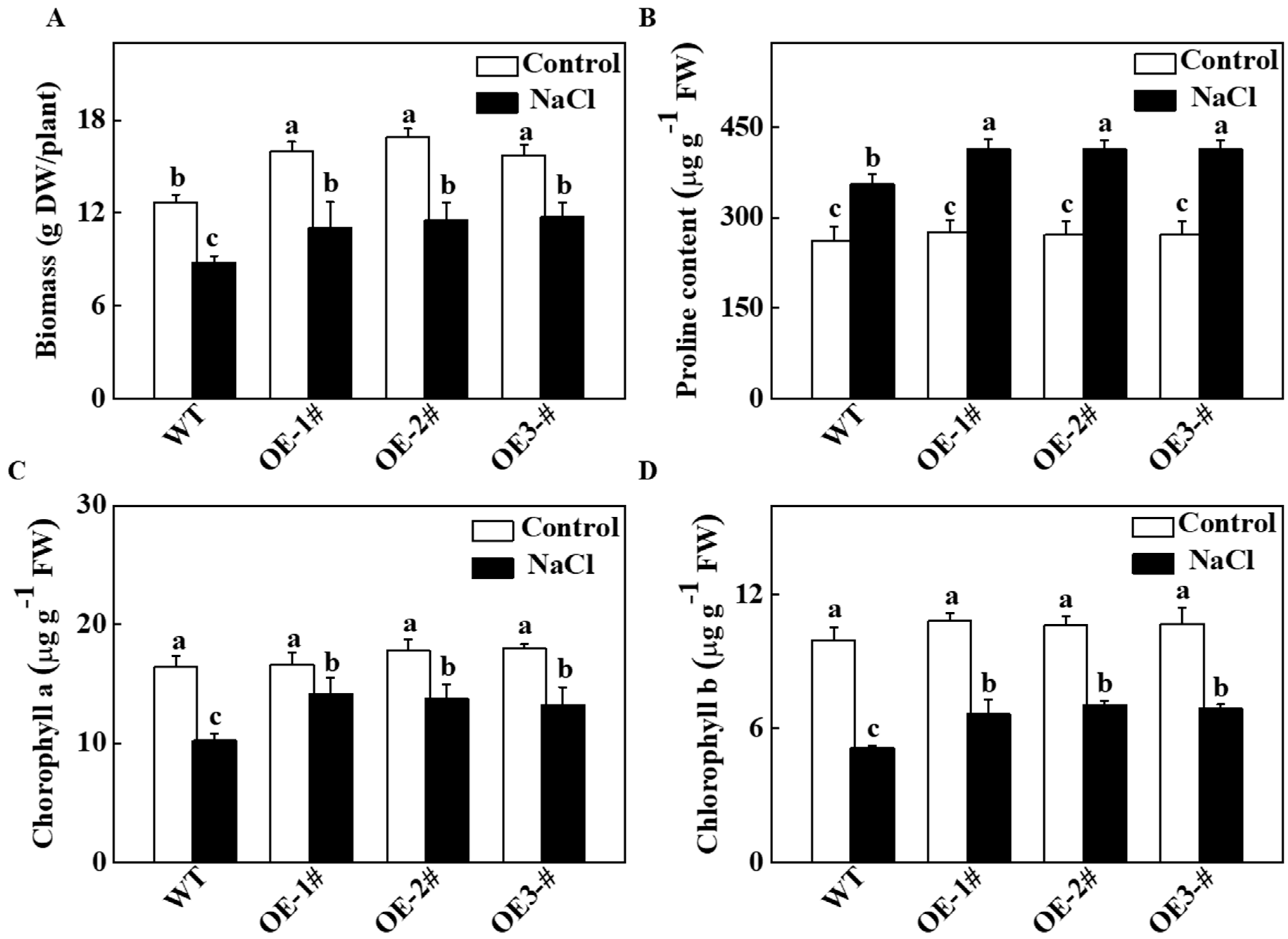
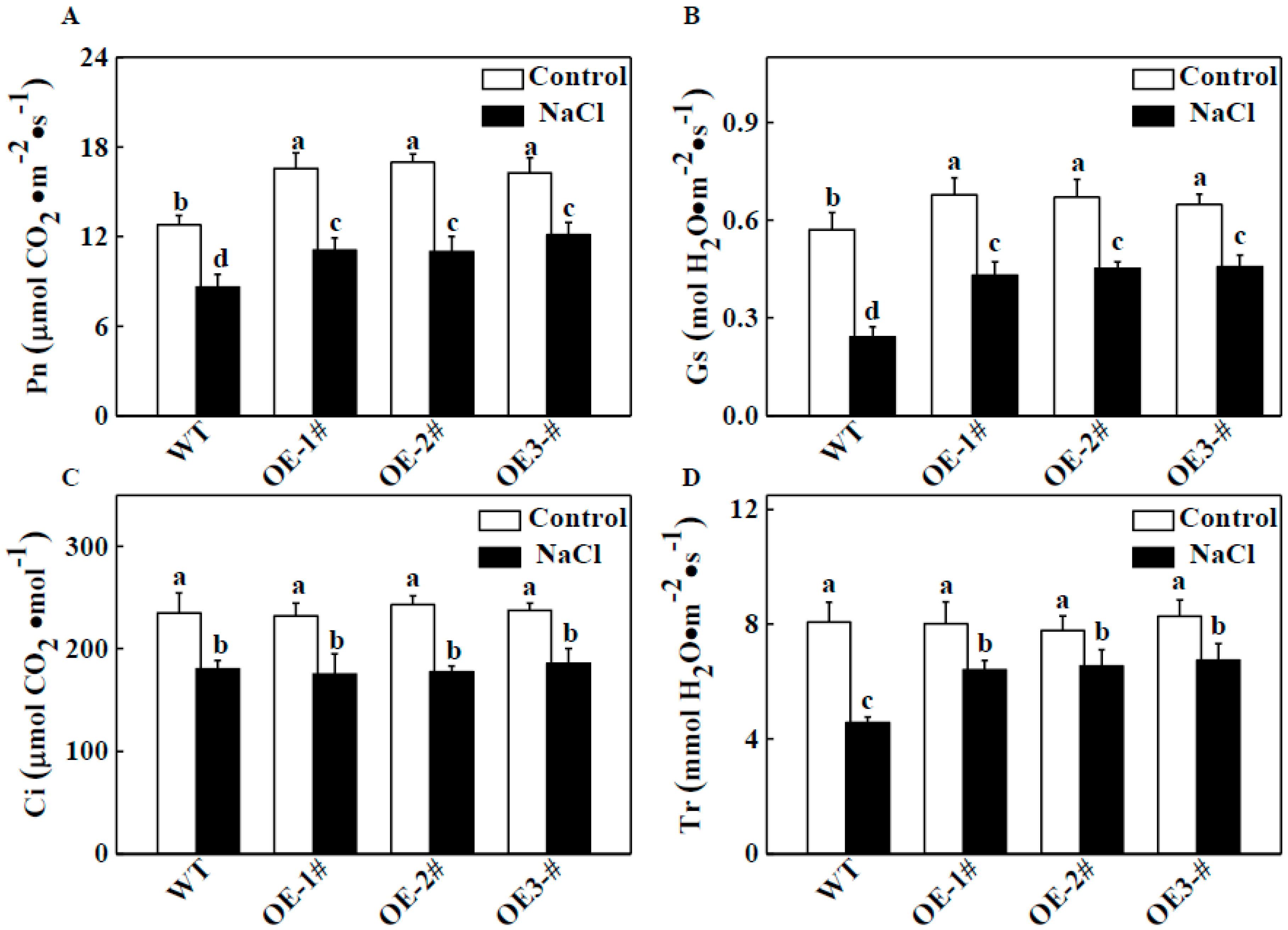
2.3. Effects of TGase on the Biomass and Photosynthetic Characteristics of Transgenic Tobacco Lines
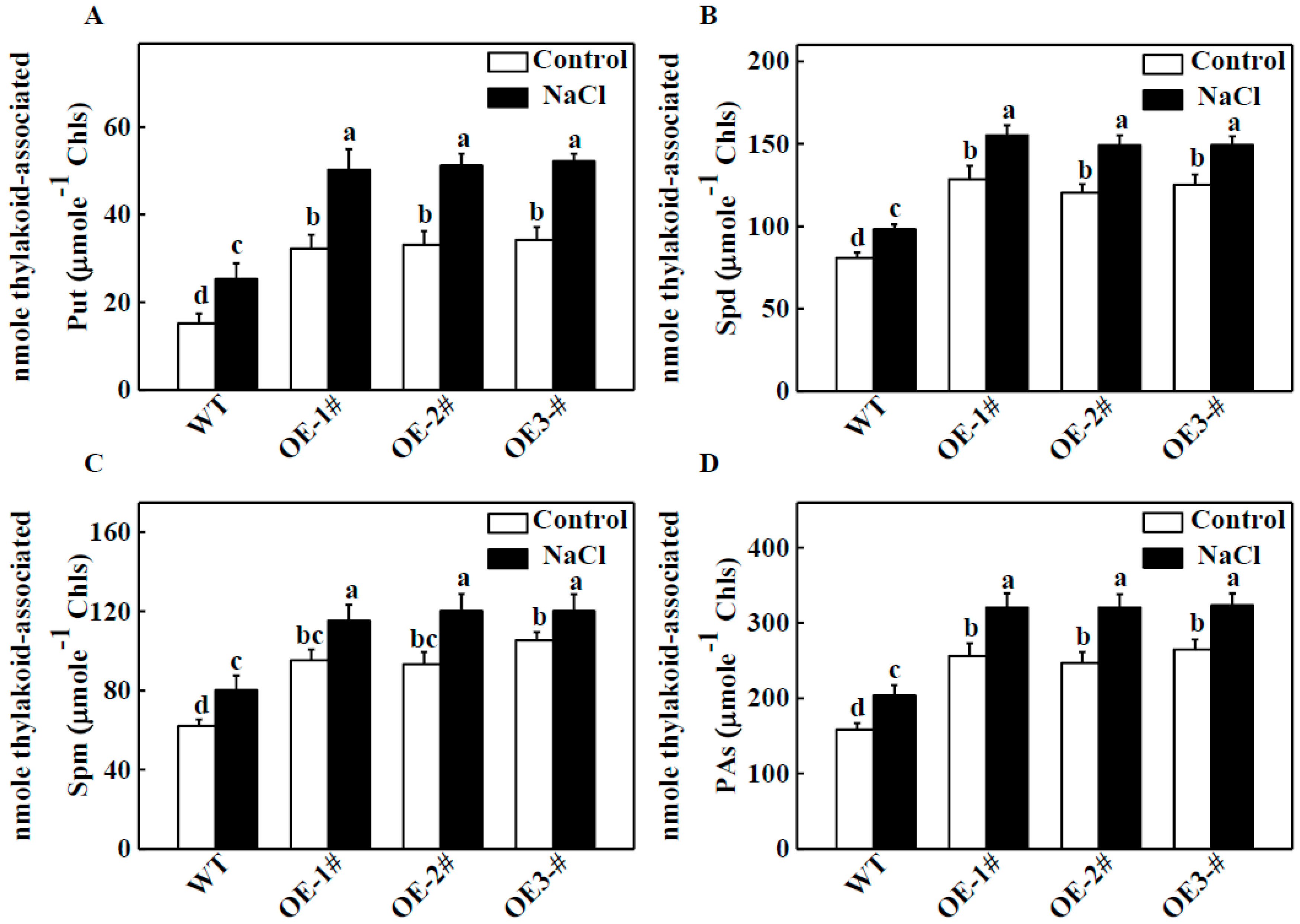
2.4. Effects of TGase on Endogenous PA Content in Thylakoid Membranes
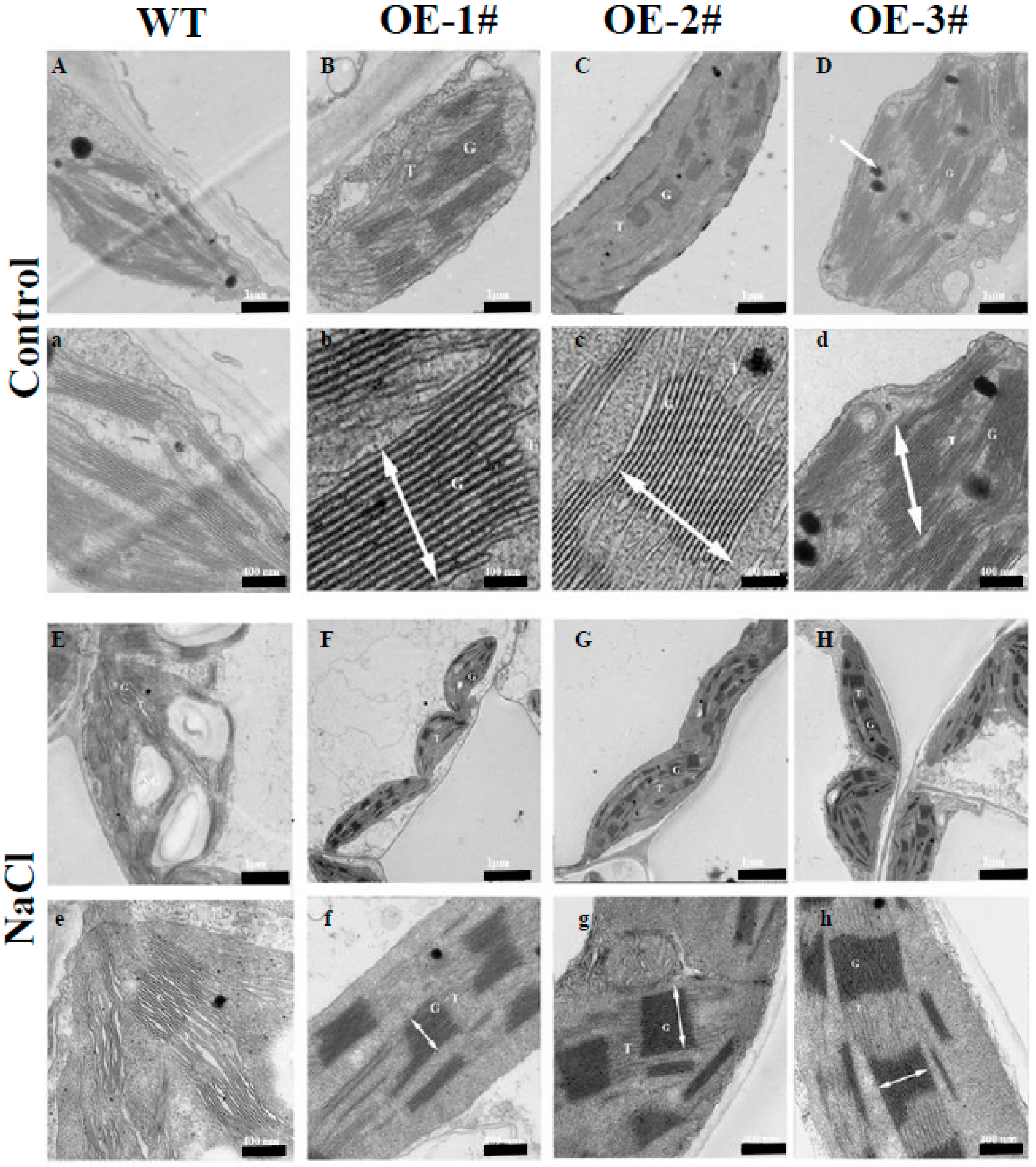
2.5. Effects of TGase on the Ultrastructure of Thylakoids
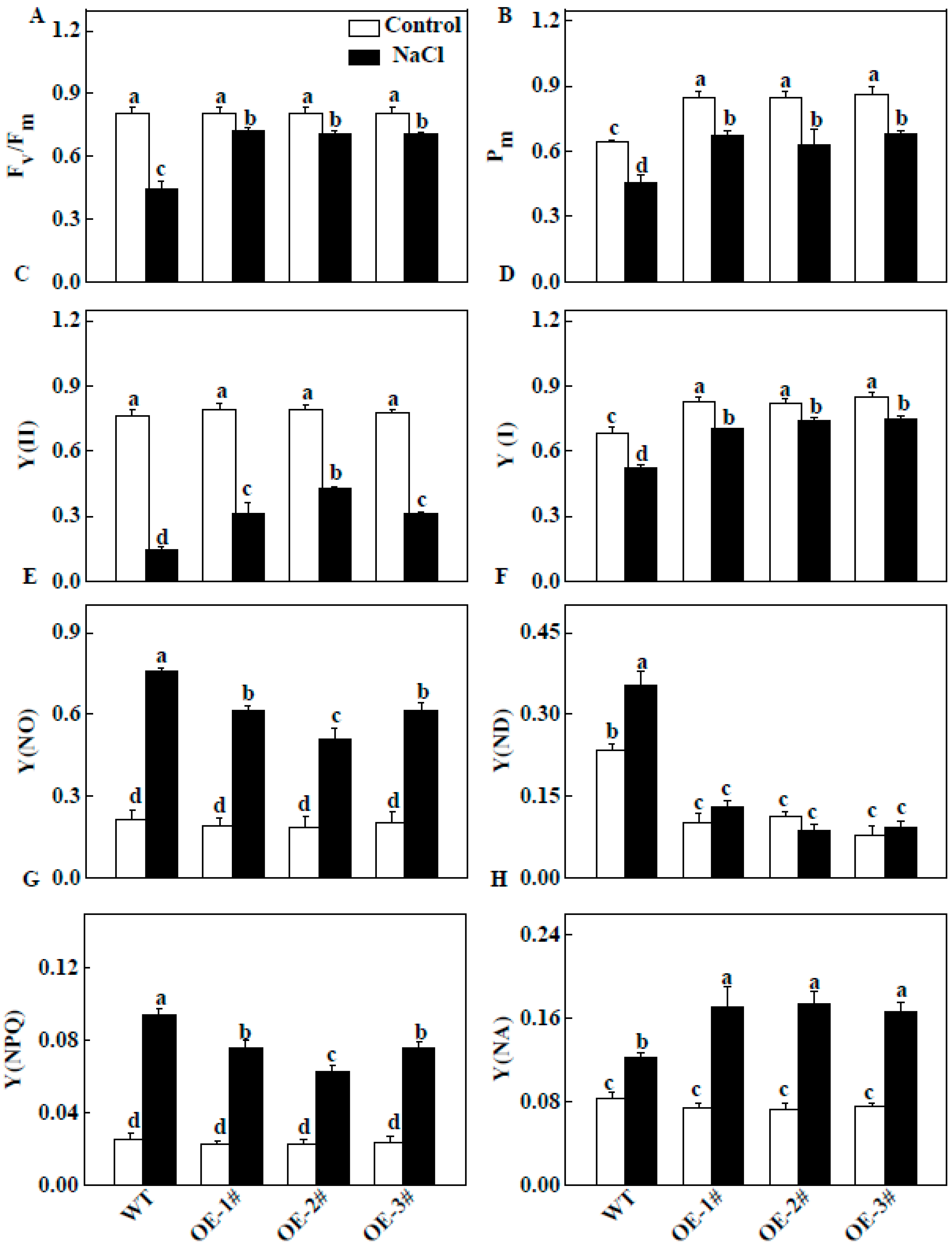
2.6. Effect of TGase on Chl a Fluorescence Transients (OJIP)
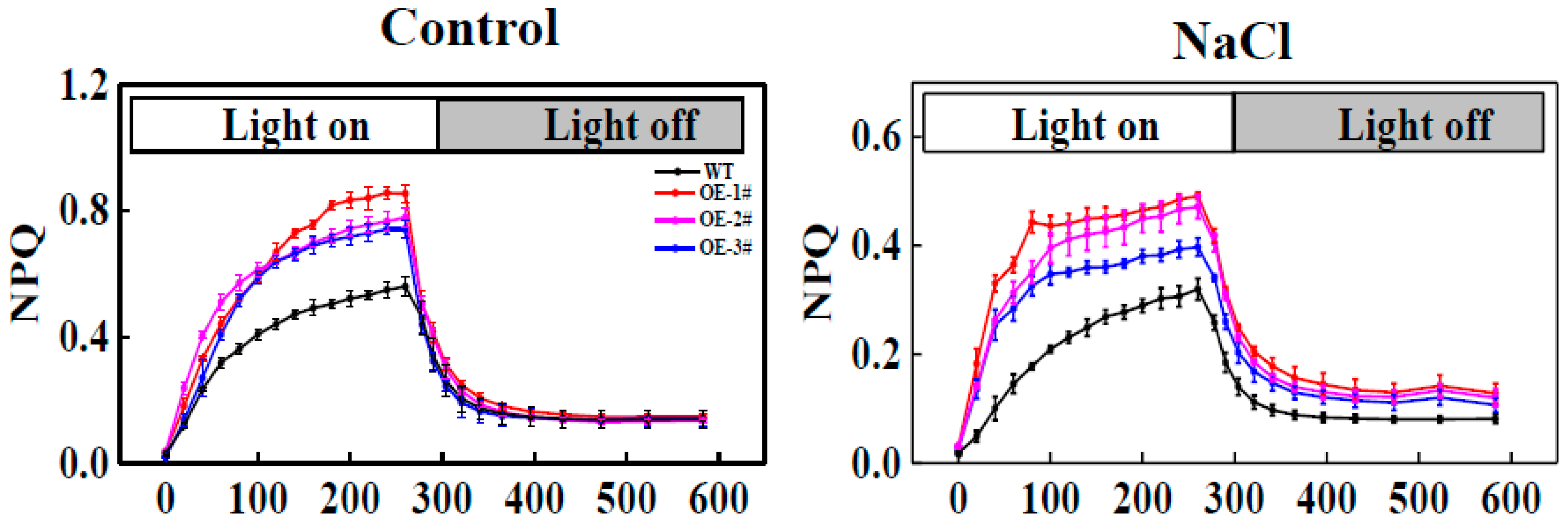
2.7. Effect of TGase on Nonphotochemical Quenching (NPQ) Induction
2.8. Effect of TGase on Quantum Yield of Energy Conversion in PSII and PSI
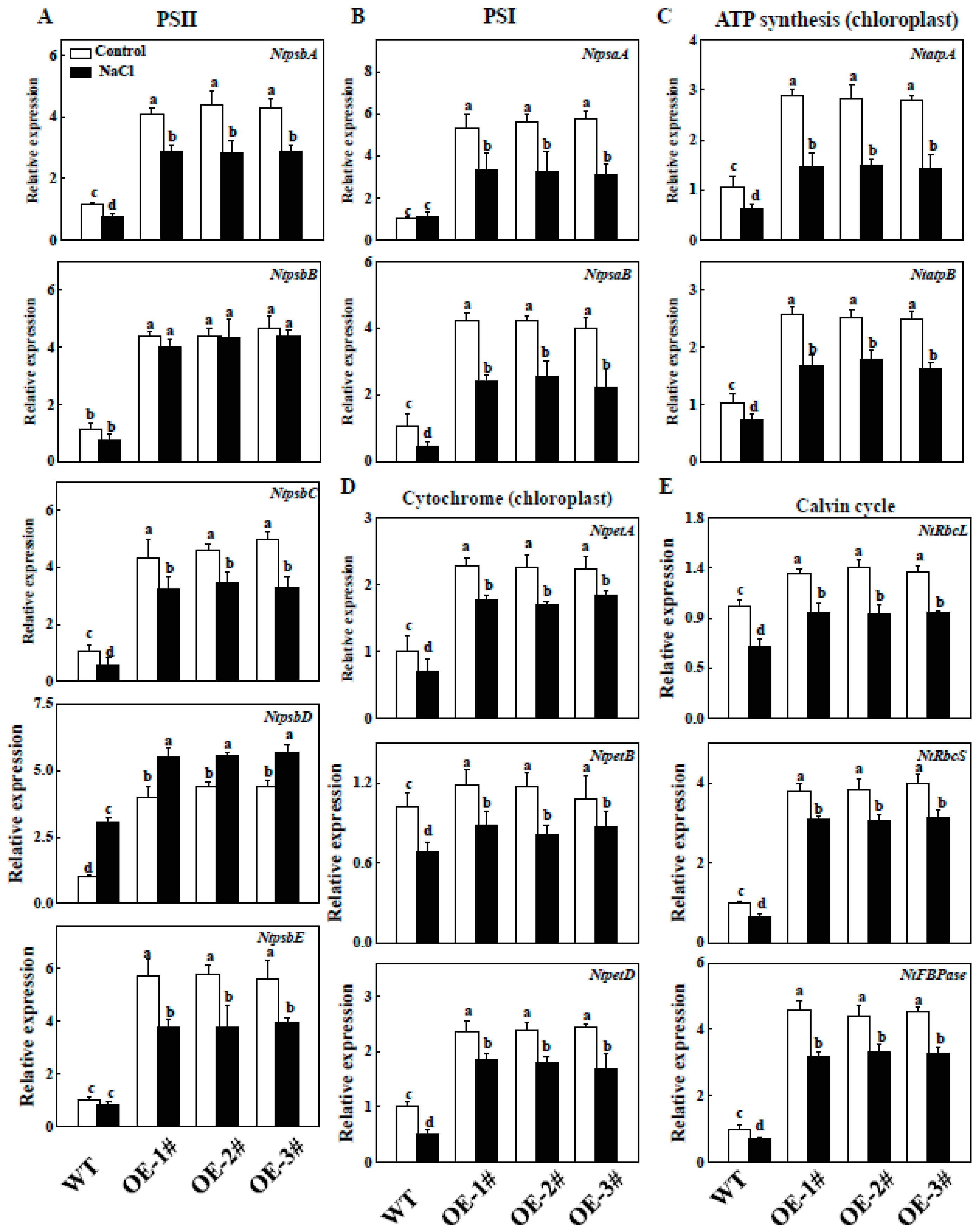
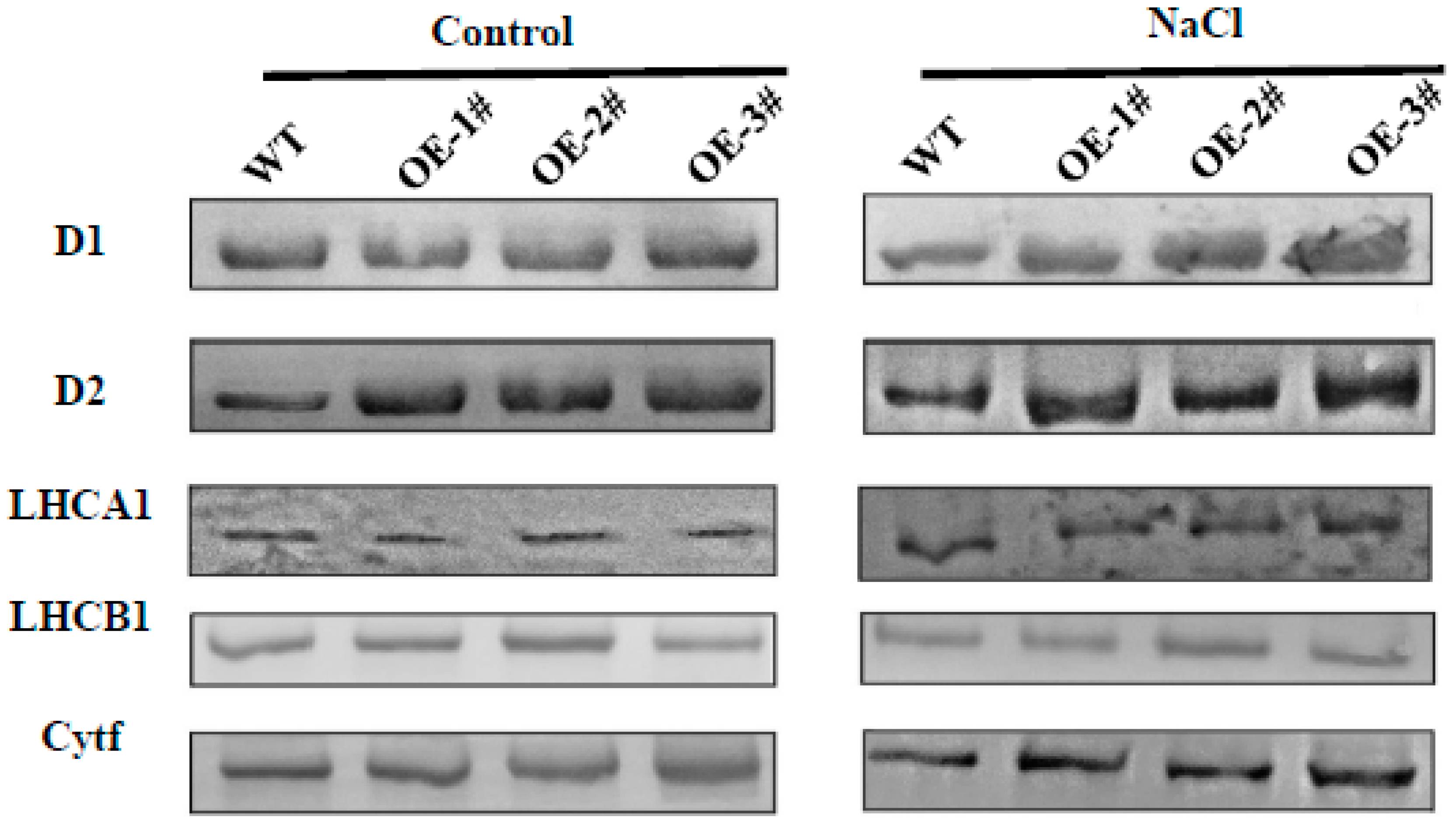
2.9. Effects of TGase on the Regulation of Photosynthesis Related Gene Expression
3. Discussion
4. Materials and Methods
4.1. Cucumber Plant Materials and Treatments for Expression Analysis
4.2. TEM Observations: Immunogold Transmission Electron Microscopy
4.3. Generation and Selection of Transgenic Plants
4.4. Salt Tolerance Analysis of the Transgenic Plants
4.5. Thylakoid Isolation
4.6. Analysis of Endogenous Polyamines in Thylakoid Membranes
4.7. Observation of the Ultrastructure of the Chloroplast
4.8. Chl a fluorescence Measurement and OJIP Transient Analyses
4.9. Quantitative Real-Time PCR
4.10. Protein Extraction and Western Blotting
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ioannidis, N.E.; Kotzabasis, K. Polyamines in chemiosmosis in vivo: A cunning mechanism for the regulation of ATP synthesis during growth and stress. Front. Plant Sci. 2014, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Guo, S.; An, Y.; Shan, X.; Wang, Y.; Shu, S.; Sun, J. Exogenous spermidine delays chlorophyll metabolism in cucumber leaves (Cucumis sativus L.) under high temperature stress. Acta Physiol. Plant. 2016, 38, 224. [Google Scholar] [CrossRef]
- Shu, S.; Yuan, Y.; Chen, J.; Sun, J.; Zhang, W.; Tang, Y.; Zhong, M.; Guo, S. The role of putrescine in the regulation of proteins and fatty acids of thylakoid membranes under salt stress. Sci. Rep. 2015, 5, 14390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagor, G.H.M.; Zhang, S.; Kojima, S.; Simm, S.; Berberich, T.; Kusano, T. Reducing cytoplasmic polyamine oxidase activity in Arabidopsis increases salt and drought tolerance by reducing reactive oxygen species production and increasing defense gene expression. Front. Plant Sci. 2016, 7, 214. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, C.; Cordeiro, A.; Alcázar, R.; Borrell, A.; Culiañez-Macià, F.A.; Tiburcio, A.F.; Altabella, T. Localization of arginine decarboxylase in tobacco plants. Physiol. Plant. 2004, 120, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Del Duca, S.; Serafini-Fracassini, D.; Cai, G. Senescence and programmed cell death in plants: Polyamine action mediated by transglutaminase. Front. Plant Sci. 2014, 5, 120. [Google Scholar] [CrossRef] [PubMed]
- Serafini-Fracassini, D.; Del Duca, S.; D’Orazi, D. First evidence for polyamine conjugation mediated by an enzymic activity in plants. Plant Physiol. 1988, 87, 757–761. [Google Scholar] [CrossRef]
- Tiburcio, A.F.; Altabella, T.; Bitrián, M.; Alcázar, R. The roles of polyamines during the lifespan of plants: From development to stress. Planta 2014, 240, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sfichi, L.; Loannidis, N.; Kotzabasis, K. Thylakoid-associated polyamines adjust the UV-B sensitivity of the photosynthetic apparatus by means of light-harvesting complex II changes. Photochem. Photobiol. 2004, 80, 499–506. [Google Scholar] [CrossRef]
- Demetriou, G.; Neonaki, C.; Navakoudis, E.; Kotzabasis, K. Salt stress impact on the molecular structure and function of the photosynthetic apparatus—The protective role of polyamines. Biochim. Biophys. Acta 2007, 1767, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Dondini, L.; Del Duca, S.; Dall’Agata, L.; Bassi, R.; Gastaldelli, M.; Della Mea, M.; Di Sandro, A.; Claparols, I.; Serafini-Fracassini, D. Suborganellar localisation and effect of light on Helianthus tuberosus chloroplast transglutaminases and their substrates. Planta 2003, 217, 84–95. [Google Scholar] [PubMed]
- Shu, S.; Yuan, L.-Y.; Guo, S.-R.; Sun, J.; Yuan, Y.-H. Effects of exogenous spermine on chlorophyll fluorescence, antioxidant system and ultrastructure of chloroplasts in Cucumis sativus L. under salt stress. Plant Physiol. Biochem. 2013, 63, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Hamdani, S.; Yaakoubi, H.; Carpentier, R. Polyamines interaction with thylakoid proteins during stress. J. Photochem. Photobiol. 2011, 104, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, N.E.; Cruz, J.A.; Kotzabasis, K.; Kramer, D.M. Evidence that putrescine modulates the higher plant photosynthetic proton circuit. PLoS ONE 2012, 7, e29864. [Google Scholar] [CrossRef]
- Ioannidis, N.E.; Ortigosa, S.M.; Veramendi, J.; Pintó-Marijuan, M.; Fleck, I.; Carvajal, P.; Kotzabasis, K.; Santos, M.; Torné, J.M. Remodeling of tobacco thylakoids by over-expression of maize plastidial transglutaminase. Biochim. Biophys. Acta 2009, 1787, 1215–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serafini-Fracassini, D.; Del Duca, S. Transglutaminases: Widespread cross-linking enzymes in plants. Ann. Bot. 2008, 102, 145–152. [Google Scholar] [CrossRef]
- Lilley, G.R.; Skill, J.; Griffin, M.; Bonner, P.L. Detection of Ca2+-dependent transglutaminase activity in root and leaf tissue of monocotyledonous and dicotyledonous plants. Plant Physiol. 1998, 117, 1115–1123. [Google Scholar] [CrossRef]
- Della Mea, M.; Caparrós-Ruiz, D.; Claparols, I.; Serafini-Fracassini, D.; Rigau, J. AtPng1p. The first plant transglutaminase. Plant Physiol. 2004, 135, 2046–2054. [Google Scholar] [CrossRef]
- Del Duca, S.; Faleri, C.; Iorio, R.A.; Cresti, M.; Serafini-Fracassini, D.; Cai, G. Distribution of transglutaminase in pear pollen tubes in relation to cytoskeleton and membrane dynamics. Plant Physiol. 2013, 161, 1706–1721. [Google Scholar] [CrossRef]
- Del Duca, S.; Tidu, V.; Bassi, R.; Esposito, C.; Serafmi-Fracassini, D. Identification of chlorophyll-a/b proteins as substrates of transglutaminase activity in isolated chloroplasts of Helianthus tuberosus L. Planta 1994, 193, 283–289. [Google Scholar] [CrossRef]
- Campos, N.; Castañón, S.; Urreta, I.; Santos, M.; Torné, J. Rice transglutaminase gene: Identification, protein expression, functionality, light dependence and specific cell location. Plant Sci. 2013, 205, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, E.; Santos, M.; Talavera, D.; Rodrıguez-Falcón, M.; Torné, J. Molecular cloning and characterization of a maize transglutaminase complementary DNA. Gene 2004, 336, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Li, J.; Guo, S.; Kang, Y. Exogenous spermidine affects polyamine metabolism in salinity-stressed Cucumis sativus roots and enhances short-term salinity tolerance. J. Plant Physiol. 2008, 165, 1620–1635. [Google Scholar] [CrossRef] [PubMed]
- Sang, Q.; Shan, X.; An, Y.; Shu, S.; Sun, J.; Guo, S. Proteomic analysis reveals the positive effect of exogenous spermidine in tomato seedlings’ response to high-temperature stress. Front. Plant Sci. 2017, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-Y.; Yuan, Y.-H.; Shu, S.; Guo, S.-R. Regulatory mechanism of NaCl stress on photosynthesis and antioxidant capacity mediated by transglutaminase in cucumber (Cucumis sativus L.) seedlings. Sci. Hortic. 2018, 235, 294–306. [Google Scholar] [CrossRef]
- Sobieszczuk-Nowicka, E.; Zmienko, A.; Samelak-Czajka, A.; Łuczak, M.; Pietrowska-Borek, M.; Iorio, R.; Del Duca, S.; Figlerowicz, M.; Legocka, J. Dark-induced senescence of barley leaves involves activation of plastid transglutaminases. Amino Acids 2015, 47, 825–838. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zhong, M.; Shu, S.; Du, N.; Sun, J.; Guo, S. Proteomic and physiological analyses reveal putrescine responses in roots of cucumber stressed by NaCl. Front. Plant Sci. 2016, 7, 1035. [Google Scholar] [CrossRef]
- Zhuo, C.; Liang, L.; Zhao, Y.; Guo, Z.; Lu, S. A cold responsive ethylene responsive factor from Medicago falcata confers cold tolerance by up-regulation of polyamine turnover, antioxidant protection, and proline accumulation. Plant Cell Environ. 2018, 41, 2021–2032. [Google Scholar] [CrossRef]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Ioannidis, N.E.; Malliarakis, D.; Torné, J.M.; Santos, M.; Kotzabasis, K. The over-expression of the plastidial transglutaminase from maize in Arabidopsis increases the activation threshold of photoprotection. Front. Plant Sci. 2016, 7, 635. [Google Scholar] [CrossRef] [PubMed]
- Del Duca, S.; Tidu, V.; Bassi, R.; Serafini-Fracassini, D.; Esposito, C. Identification of transglutaminase activity and its substrates in isolated chloroplast of Helianthus tuberosus. Planta 1994, 193, 283–289. [Google Scholar] [CrossRef]
- Sobieszczuk-Nowicka, E.; Krzesłowska, M.; Legocka, J. Transglutaminases and their substrates in kinetin-stimulated etioplast-to-chloroplast transformation in cucumber cotyledons. Protoplasma 2008, 233, 187. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yang, S.-J.; Zhang, S.-B.; Zhang, J.-L.; Cao, K.-F. Cyclic electron flow plays an important role in photoprotection for the resurrection plant Paraboearufescens under drought stress. Planta 2012, 235, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Carvajal-Vallejos, P.; Villalobos, E.; Franco, C.; Almeida, A.; Coelho, A.; Torné, J.; Santos, M. Characterisation of Zea mays L. plastidial transglutaminase: Interactions with thylakoid membrane proteins. Plant Biol. 2010, 12, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Campos, N.; Villalobos, E.; Fontanet, P.; Torné, J.M.; Santos, M. A peptide of 17 aminoacids from the N-terminal region of maize plastidial transglutaminase is essential for chloroplast targeting. Am. J. Mol. Biol. 2012, 2, 245–257. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A simple and general-method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, Z.-Z.; Zhou, X.-F.; Yin, H.-B.; Li, X.; Xin, X.-F.; Hong, X.-H.; Zhu, J.-K.; Gong, Z. Overexpression of SOS (Salt Overly Sensitive) genes increases salt tolerance in transgenic Arabidopsis. Mol. Plant 2009, 2, 22–31. [Google Scholar] [CrossRef]
- Zhang, R.H.; Li, J.; Guo, S.R.; Tezuka, T. Effects of exogenous putrescine on gas-exchange characteristics and chlorophyll fluorescence of NaCl-stressed cucumber seedlings. Photosynth. Res. 2009, 100, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Force, L.; Critchley, C.; van Rensen, J.J. New fluorescence parameters for monitoring photosynthesis in plants. Photosynth. Res. 2003, 78, 17. [Google Scholar] [CrossRef] [PubMed]
- Bilger, W.; Björkman, O. Relationships among violaxanthin deepoxidation, thylakoid membrane conformation, and nonphotochemical chlorophyll fluorescence quenching in leaves of cotton (Gossypium hirsutum L.). Planta 1994, 193, 238–246. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, M.; Wang, Y.; Zhang, Y.; Shu, S.; Sun, J.; Guo, S. Overexpression of Transglutaminase from Cucumber in Tobacco Increases Salt Tolerance through Regulation of Photosynthesis. Int. J. Mol. Sci. 2019, 20, 894. https://doi.org/10.3390/ijms20040894
Zhong M, Wang Y, Zhang Y, Shu S, Sun J, Guo S. Overexpression of Transglutaminase from Cucumber in Tobacco Increases Salt Tolerance through Regulation of Photosynthesis. International Journal of Molecular Sciences. 2019; 20(4):894. https://doi.org/10.3390/ijms20040894
Chicago/Turabian StyleZhong, Min, Yu Wang, Yuemei Zhang, Sheng Shu, Jin Sun, and Shirong Guo. 2019. "Overexpression of Transglutaminase from Cucumber in Tobacco Increases Salt Tolerance through Regulation of Photosynthesis" International Journal of Molecular Sciences 20, no. 4: 894. https://doi.org/10.3390/ijms20040894
APA StyleZhong, M., Wang, Y., Zhang, Y., Shu, S., Sun, J., & Guo, S. (2019). Overexpression of Transglutaminase from Cucumber in Tobacco Increases Salt Tolerance through Regulation of Photosynthesis. International Journal of Molecular Sciences, 20(4), 894. https://doi.org/10.3390/ijms20040894