Transcriptome Profile of the Variegated Ficus microcarpa c.v. Milky Stripe Fig Leaf


Abstract
:1. Introduction
2. Results and Discussion
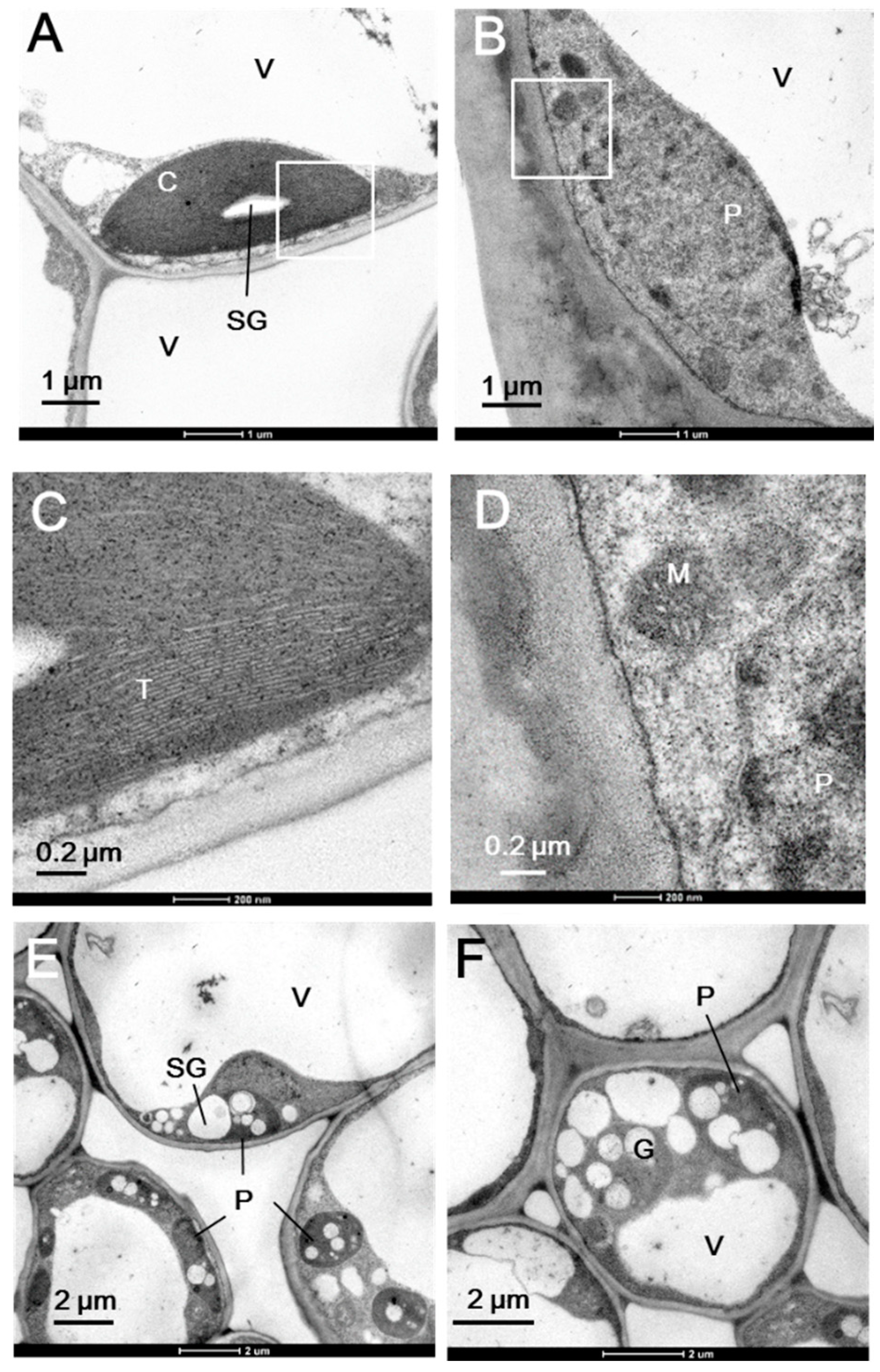
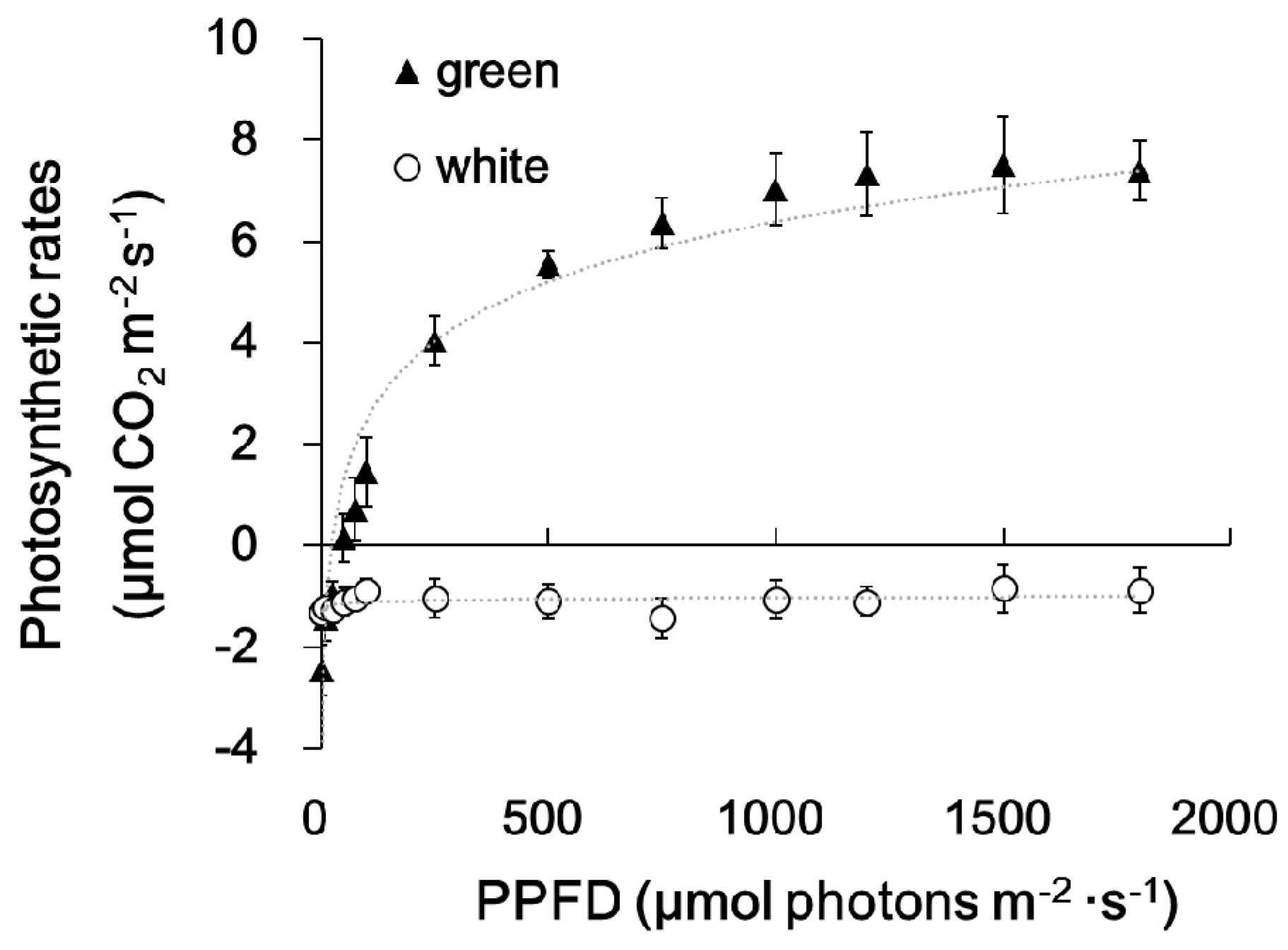
2.1. Plastid Morphology and Photosynthetic Activity
2.2. De novo Assembly and Bioinformatic Analysis
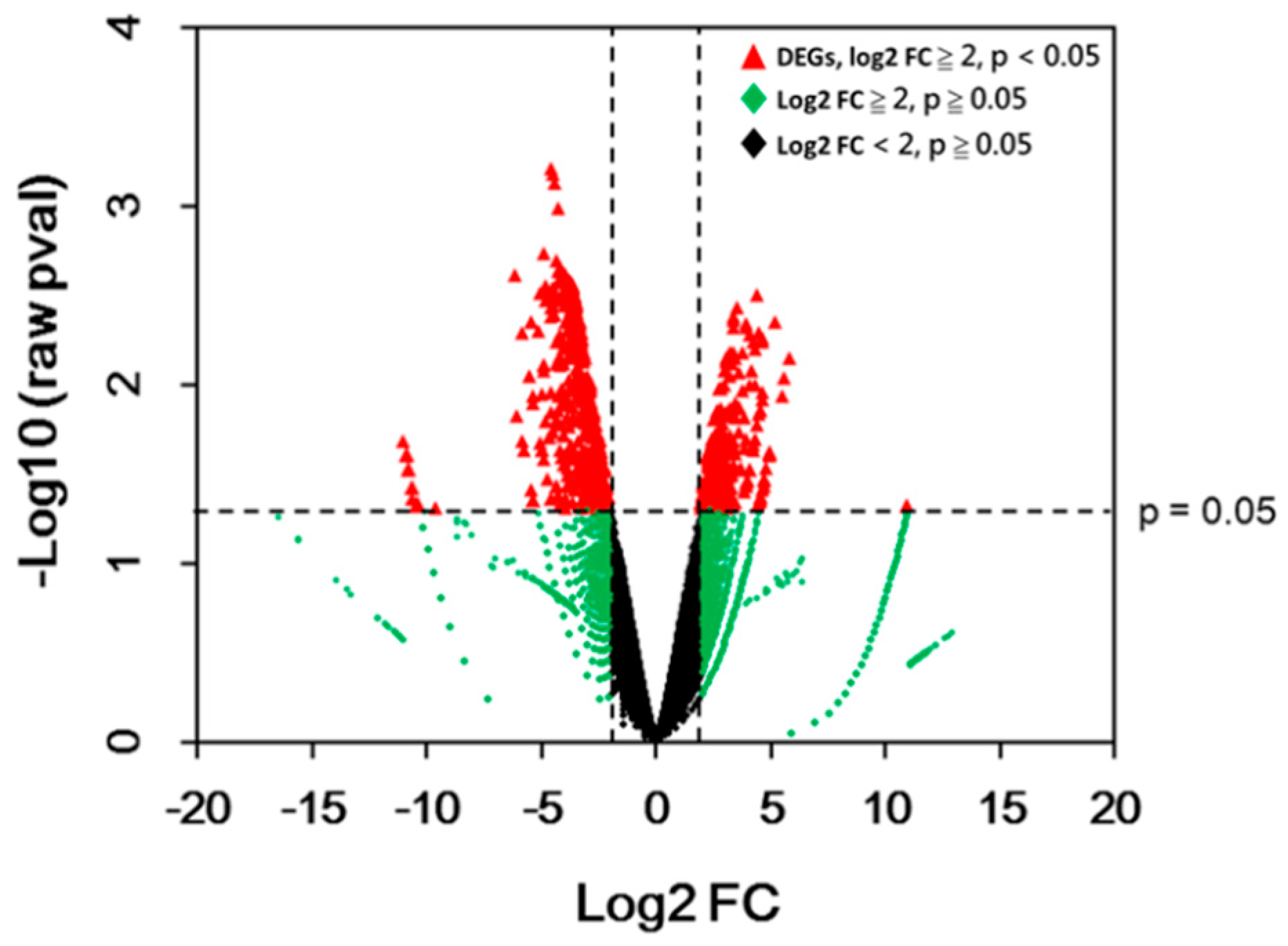
2.3. Differential Expressed Genes (DEGs)
2.3.1. Photosynthesis and Chlorophyll Metabolism
2.3.2. Chloroplast Organization
2.3.3. Nitrate Metabolism
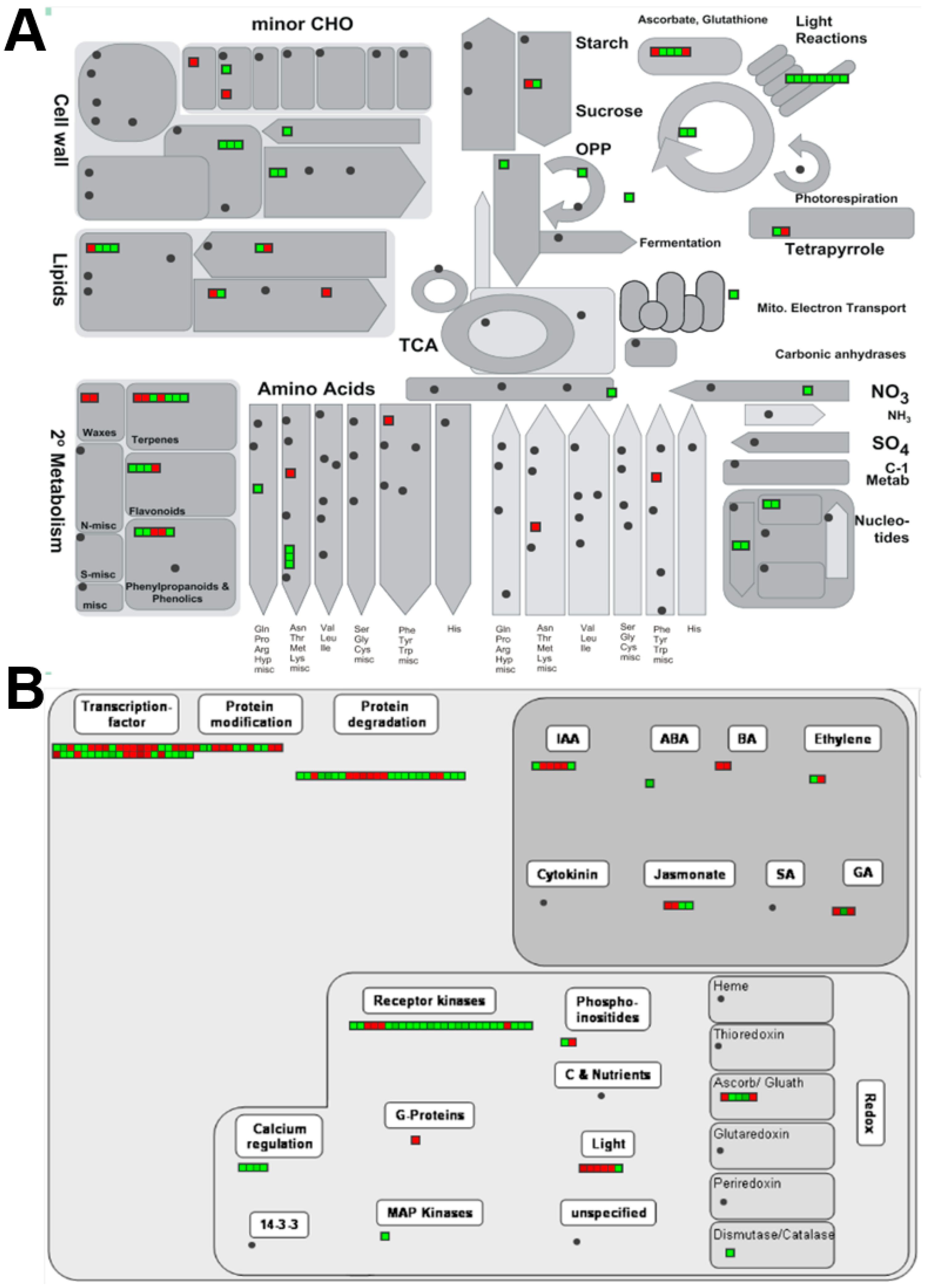
2.4. Over-Represented Analysis (ORA)
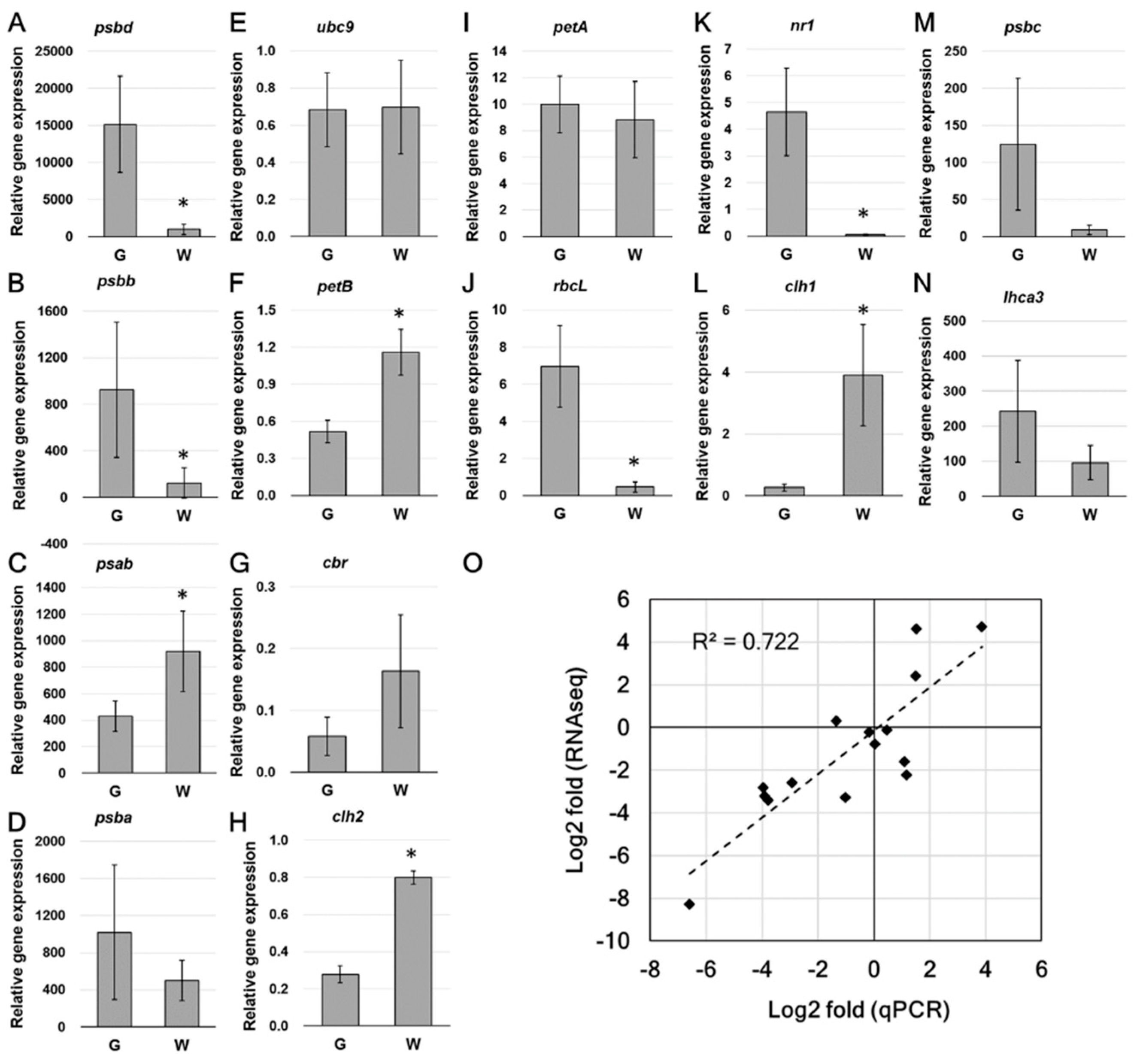
2.5. Real Time Quantitative PCR
3. Material and Methods
3.1. Plant Material
3.2. Light Response Curve
3.3. Transmission Electron Microscopy (TEM)
3.4. RNA Purification, Complementary cDNA Library Construction, and Transcriptome Sequencing
3.5. De novo Assembly, BLAST, and RNA-Sequencing (RNA-Seq)
3.6. Quantitative Reverse-Transcription Polymerase Chain Reaction (RT-qPCR)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, A.P. Ecology of a leaf color polymorphism in a tropical forest species: Habitat segregation and herbivory. Oecologia 1986, 69, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Soltau, U.; Dötterl, S.; Liede-Schumann, S. Leaf variegation in Caladium steudneriifolium (Araceae): A case of mimicry? Evol. Ecol. 2009, 23, 503–512. [Google Scholar] [CrossRef]
- Aluru, M.R.; Bae, H.; Wu, D.; Rodermel, S.R. The Arabidopsis immutans mutation affects plastid differentiation and the morphogenesis of white and green sectors in variegated plants. Plant Physiol. 2001, 127, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Beardsell, D.; Norden, U. Ficus rubiginosa ‘variegata’, a chlorophyll-deficient chimera with mosaic patterns created by cell divisions from the outer meristematic layer. Ann. Bot. 2004, 94, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kouso, T.; Sakamoto, W. Variegated tobacco leaves generated by chloroplast FtsH suppression: Implication of FtsH function in the maintenance of thylakoid membranes. Plant Cell Physiol. 2012, 53, 391–404. [Google Scholar] [CrossRef]
- Lysenko, V. Fluorescence kinetic parameters and cyclic electron transport in guard cell chloroplasts of chlorophyll-deficient leaf tissues from variegated weeping fig (Ficus benjamina L.). Planta 2012, 235, 1023–1033. [Google Scholar] [CrossRef]
- Aluru, M.R.; Yu, F.; Fu, A.; Rodermel, S. Arabidopsis variegation mutants: New insights into chloroplast biogenesis. J. Exp. Bot. 2006, 57, 1871–1881. [Google Scholar] [CrossRef]
- Joet, T.; Genty, B.; Josse, E.M.; Kuntz, M.; Cournac, L.; Peltier, G. Involvement of a plastid terminal oxidase in plastoquinone oxidation as evidenced by expression of the Arabidopsis thaliana enzyme in tobacco. J. Biol. Chem. 2002, 277, 31623–31630. [Google Scholar] [CrossRef]
- Rosso, D.; Ivanov, A.G.; Fu, A.; Geisler-Lee, J.; Hendrickson, L.; Geisler, M.; Stewart, G.; Krol, M.; Hurry, V.; Rodermel, S.R.; et al. IMMUTANS does not act as a stress-induced safety valve in the protection of the photosynthetic apparatus of Arabidopsis during steady-state photosynthesis. Plant Physiol. 2006, 142, 574–585. [Google Scholar] [CrossRef]
- Wetzel, C.M.; Jiang, C.Z.; Meehan, L.J.; Voytas, D.F.; Rodermel, S.R. Nuclear-organelle interactions: The immutans variegation mutant of Arabidopsis is plastid autonomous and impaired in carotenoid biosynthesis. Plant J. 1994, 6, 161–175. [Google Scholar] [CrossRef]
- Kato, Y.; Miura, E.; Matsushima, R.; Sakamoto, W. White leaf sectors in yellow variegated2 are formed by viable cells with undifferentiated plastids. Plant Physiol. 2007, 144, 952–960. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, X.; Xu, B.; Li, Y.; Ma, Y.; Wang, G. Phenotype and transcriptome analysis reveals chloroplast development and pigment biosynthesis together influenced the leaf color formation in mutants of Anthurium andraeanum ‘Sonate’. Front. Plant Sci. 2015, 6, 139. [Google Scholar] [CrossRef]
- Miura, E.; Kato, Y.; Matsushima, R.; Albrecht, V.; Laalami, S.; Sakamoto, W. The balance between protein synthesis and degradation in chloroplasts determines leaf variegation in Arabidopsis yellow variegated mutants. Plant Cell 2007, 19, 1313–1328. [Google Scholar] [CrossRef]
- Bae, C.H.; Abe, T.; Matsuyama, T.; Fukunishi, N.; Nagata, N.; Nakano, T.; Kaneko, Y.; Miyoshi, K.; Matsushima, H.; Yoshida, S. Regulation of chloroplast gene expression is affected in ali, a novel tttobacco albino mutant. Ann. Bot. 2001, 88, 545–553. [Google Scholar] [CrossRef]
- Sheue, C.R.; Pao, S.H.; Chien, L.F.; Chesson, P.; Peng, C.I. Natural foliar variegation without costs? The case of Begonia. Ann. Bot. 2012, 109, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Miura, E.; Kato, Y.; Sakamoto, W. Comparative transcriptome analysis of green/white variegated sectors in Arabidopsis yellow variegated2: Responses to oxidative and other stresses in white sectors. J. Exp. Bot. 2010, 61, 2433–2445. [Google Scholar] [CrossRef]
- Aluru, M.R.; Zola, J.; Foudree, A.; Rodermel, S.R. Chloroplast photooxidation-induced transcriptome reprogramming in Arabidopsis immutans white leaf sectors. Plant Physiol. 2009, 150, 904–923. [Google Scholar] [CrossRef]
- Satou, M.; Enoki, H.; Oikawa, A.; Ohta, D.; Saito, K.; Hachiya, T.; Sakakibara, H.; Kusano, M.; Fukushima, A.; Saito, K.; et al. Integrated analysis of transcriptome and metabolome of Arabidopsis albino or pale green mutants with disrupted nuclear-encoded chloroplast proteins. Plant Mol. Biol. 2014, 85, 411–428. [Google Scholar] [CrossRef]
- Chen, M.C.M.; Yang, J.H.; Liu, C.H.; Lin, K.H.; Yang, C.M. Molecular, structural, and phylogenetic characterization of two chlorophyllase isoforms in Pachira macrocarpa. Plant Syst. Evol. 2014, 300, 633–643. [Google Scholar] [CrossRef]
- Chen, M.C.M.; Chao, P.Y.; Huang, M.Y.; Yang, J.H.; Yang, Z.W.; Lin, K.H.; Yang, C.M. Chlorophyllase activity in green and non-green tissues of variegated plants. S. Afr. J. Bot. 2012, 81, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Hortensteiner, S.; Krautler, B. Chlorophyll breakdown in higher plants. Biochim. Biophys. Acta 2011, 1807, 977–988. [Google Scholar] [CrossRef] [Green Version]
- Shimoda, Y.; Ito, H.; Tanaka, A. Arabidopsis STAY-GREEN, Mendel’s green cotyledon gene, encodes magnesium-dechelatase. Plant Cell 2016, 28, 2147–2160. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Park, S.Y.; Paek, N.C. The divergent roles of STAYGREEN (SGR) homologs in chlorophyll degradation. Mol. Cells 2015, 38, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Shimoda, Y.; Matsuda, K.; Tanaka, A.; Ito, H. Mg-dechelation of chlorophyll a by Stay-Green activates chlorophyll b degradation through expressing Non-Yellow Coloring 1 in Arabidopsis thaliana. J. Plant Physiol. 2018, 222, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Mullet, J.E.; Klein, P.G.; Klein, R.R. Chlorophyll regulates accumulation of the plastid-encoded chlorophyll apoproteins CP43 and D1 by increasing apoprotein stability. Proc. Natl. Acad. Sci. USA 1990, 87, 4038–4042. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, W. Leaf-variegated mutations and their responsible genes in Arabidopsis thaliana. Genes Genet. Syst. 2003, 78, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kleine, T. Arabidopsis thaliana mTERF proteins: Evolution and functional classification. Front. Plant Sci. 2012, 3, 233. [Google Scholar] [CrossRef]
- Robles, P.; Micol, J.L.; Quesada, V. Unveiling plant mTERF functions. Mol. Plant 2012, 5, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Robles, P.; Micol, J.L.; Quesada, V. Arabidopsis MDA1, a nuclear-encoded protein, functions in chloroplast development and abiotic stress responses. PLoS ONE 2012, 7, e42924. [Google Scholar] [CrossRef]
- Robles, P.; Micol, J.L.; Quesada, V. Mutations in the plant-conserved MTERF9 alter chloroplast gene expression, development and tolerance to abiotic stress in Arabidopsis thaliana. Physiol. Plant. 2015, 154, 297–313. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Plumley, F.G.; Schmidt, G.W. Nitrogen-dependent regulation of photosynthetic gene expression. Proc. Natl. Acad. Sci. USA 1989, 86, 2678–2682. [Google Scholar] [CrossRef] [Green Version]
- Saux, C.; Lemoine, Y.; Marion-Poll, A.; Valadier, M.H.; Deng, M.; Morot-Gaudry, J.F. Consequence of absence of nitrate reductase activity onphotosynthesis in Nicotiana plumbaginifolia plants. Plant Physiol. 1987, 84, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.C.; Sherameti, I.; Oelmuller, R. Siroheme: An essential component for life on earth. Plant Signal. Behav. 2010, 5, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Garai, S.; Tripathy, B.C. Alleviation of nitrogen and sulfur deficiency and enhancement of photosynthesis in Arabidopsis thaliana by overexpression of Uroporphyrinogen III methyltransferase (UPM1). Front. Plant Sci. 2017, 8, 2265. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.M.; Wang, R.L.; Zhu, T.; Rothstein, S.J. Global transcription profiling reveals differential responses to chronic nitrogen stress and putative nitrogen regulatory components in Arabidopsis. BMC Genom. 2007, 8, 281. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Barker, A.V. Ethylene evolution and ammonium accumulation by tomato plants with various nitrogen forms and regimes of acidity. Part I. J. Plant Nutr. 1992, 15, 2457–2469. [Google Scholar] [CrossRef]
- McClellan, C.A.; Chang, C. The role of protein turnover in ethylene biosynthesis and response. Plant Sci. 2008, 175, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, N.; Sato, T.; Ogura, N.; Nakagawa, H. Control by glutamine of the synthesis of nitrate reductase in cultured spinach cells. Plant Cell Physiol. 1992, 33, 727–731. [Google Scholar]
- Vincentz, M.; Moureaux, T.; Leydecker, M.T.; Vaucheret, H.; Caboche, M. Regulation of nitrate and nitrite reductase expression in Nicotiana plumbaginifolia leaves by nitrogen and carbon metabolites. Plant J. 1993, 3, 315–324. [Google Scholar] [CrossRef]
- Kato, Y.; Miura, E.; Ido, K.; Ifuku, K.; Sakamoto, W. The variegated mutants lacking chloroplastic FtsHs are defective in D1 degradation and accumulate reactive oxygen species. Plant Physiol. 2009, 151, 1790–1801. [Google Scholar] [CrossRef]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Anders, S.; McCarthy, D.J.; Chen, Y.; Okoniewski, M.; Smyth, G.K.; Huber, W.; Robinson, M.D. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat. Protoc. 2013, 8, 1765–1786. [Google Scholar] [CrossRef] [Green Version]
- Usadel, B.; Nagel, A.; Steinhauser, D.; Gibon, Y.; Blasing, O.E.; Redestig, H.; Sreenivasulu, N.; Krall, L.; Hannah, M.A.; Poree, F.; et al. PageMan: An interactive ontology tool to generate, display, and annotate overview graphs for profiling experiments. BMC Bioinform. 2006, 7, 535. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, R.; Rodiger, A.; Agne, B.; Baerenfaller, K.; Baginsky, S. Common and specific protein accumulation patterns in different albino/pale-green mutants reveals regulon organization at the proteome level. Plant Physiol. 2012, 160, 2189–2201. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NGS and de novo Assembly | |
|---|---|
| Reads of G | 23,679,092 |
| Reads of W | 22,358,400 |
| De novo assembled contig | |
| G | 95,394 |
| W | 102,861 |
| RNASeq | |
| Annotated contigs | 61,363 |
| Genes after trim | 59,602 |
| DEGs | 467 |
| up-regulated DEGs | 183 |
| down-regulated DEGs | 284 |
| Symbol | Log 2 Fold Change | At Orthologue | Description |
|---|---|---|---|
| Photosystem | |||
| psba | −3.3 | ATCG00020 | PSII D1 protein |
| psbb | −2.6 | ATCG00680 | CP47 subunit of the PSII |
| psbc | −3.4 | ATCG00280 | CP43 subunit of the PSII |
| psbd | −3.2 | ATCG00270 | PSII D2 protein |
| psbh | −2.4 | ATCG00710 | PSII reaction center protein H |
| psaa | −2.3 | ATCG00350 | PsaA protein comprising the reaction center for PSI |
| rbcl | −2.8 | ATCG00490 | Large subunit of RUBISCO |
| petb | −2.2 | ATCG00720 | Cytochrome b(6) subunit of the cytochrome b6f complex |
| ndhb.2 | −3.0 | ATCG01250 | NADH dehydrogenase ND2 |
| Tetrapyrrole metabolism | |||
| upm1 | −3.2 | AT5G40850 | Urophorphyrin III methylase |
| nyc1 | 2.4 | AT4G13250 | Chlorophyll b reductase |
| clh1 | 4.7 | AT1G19670 | Chlorophyllase I |
| clh2 | 4.6 | AT5G43860 | Chlorophyllase II |
| nye1 | 3.1 | AT4G22920 | Non-yellowing 1, stay-green 1 |
| sgrl | 3.7 | AT1G44000 | Stay-green like protein |
| Chloroplast organization | |||
| hcf173 | 3.4 | AT1G16720 | High chlorophyll fluorescence phenotype 173 |
| sig5 | 2.8 | AT5G24120 | Sigma factor 5 |
| sco3 | −3.1 | AT3G19570 | Snowy cotyledon 3, QWRF domain containing 1 |
| mterf9 | −10.4 | AT5G55580 | Mitochondrial transcription termination factor (mTERF) 9 |
| emb2219 | −3.2 | AT2G21710 | Embryo defective 2219 (mTERF) |
| mterf* | −2.4 | AT1G78930 | Mitochondrial transcription termination factor (mTERF) |
| Nitrate metabolism | |||
| nir1 | −2.9 | AT2G15620 | Nitrite reductase 1 (NiR1) |
| nrt1 | −2.4 | AT1G69850 | Nitrate transporter 1 (NRT1) |
| xip1 | −4.1 | AT5G49660 | Xylem intermixed with phloem 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shih, T.-H.; Lin, S.-H.; Huang, M.-Y.; Huang, W.-D.; Yang, C.-M. Transcriptome Profile of the Variegated Ficus microcarpa c.v. Milky Stripe Fig Leaf. Int. J. Mol. Sci. 2019, 20, 1338. https://doi.org/10.3390/ijms20061338
Shih T-H, Lin S-H, Huang M-Y, Huang W-D, Yang C-M. Transcriptome Profile of the Variegated Ficus microcarpa c.v. Milky Stripe Fig Leaf. International Journal of Molecular Sciences. 2019; 20(6):1338. https://doi.org/10.3390/ijms20061338
Chicago/Turabian StyleShih, Tin-Han, Szu-Hsien Lin, Meng-Yuan Huang, Wen-Dar Huang, and Chi-Ming Yang. 2019. "Transcriptome Profile of the Variegated Ficus microcarpa c.v. Milky Stripe Fig Leaf" International Journal of Molecular Sciences 20, no. 6: 1338. https://doi.org/10.3390/ijms20061338
APA StyleShih, T. -H., Lin, S. -H., Huang, M. -Y., Huang, W. -D., & Yang, C. -M. (2019). Transcriptome Profile of the Variegated Ficus microcarpa c.v. Milky Stripe Fig Leaf. International Journal of Molecular Sciences, 20(6), 1338. https://doi.org/10.3390/ijms20061338