AMPA Receptor Auxiliary Proteins of the CKAMP Family


Abstract
:1. Introduction
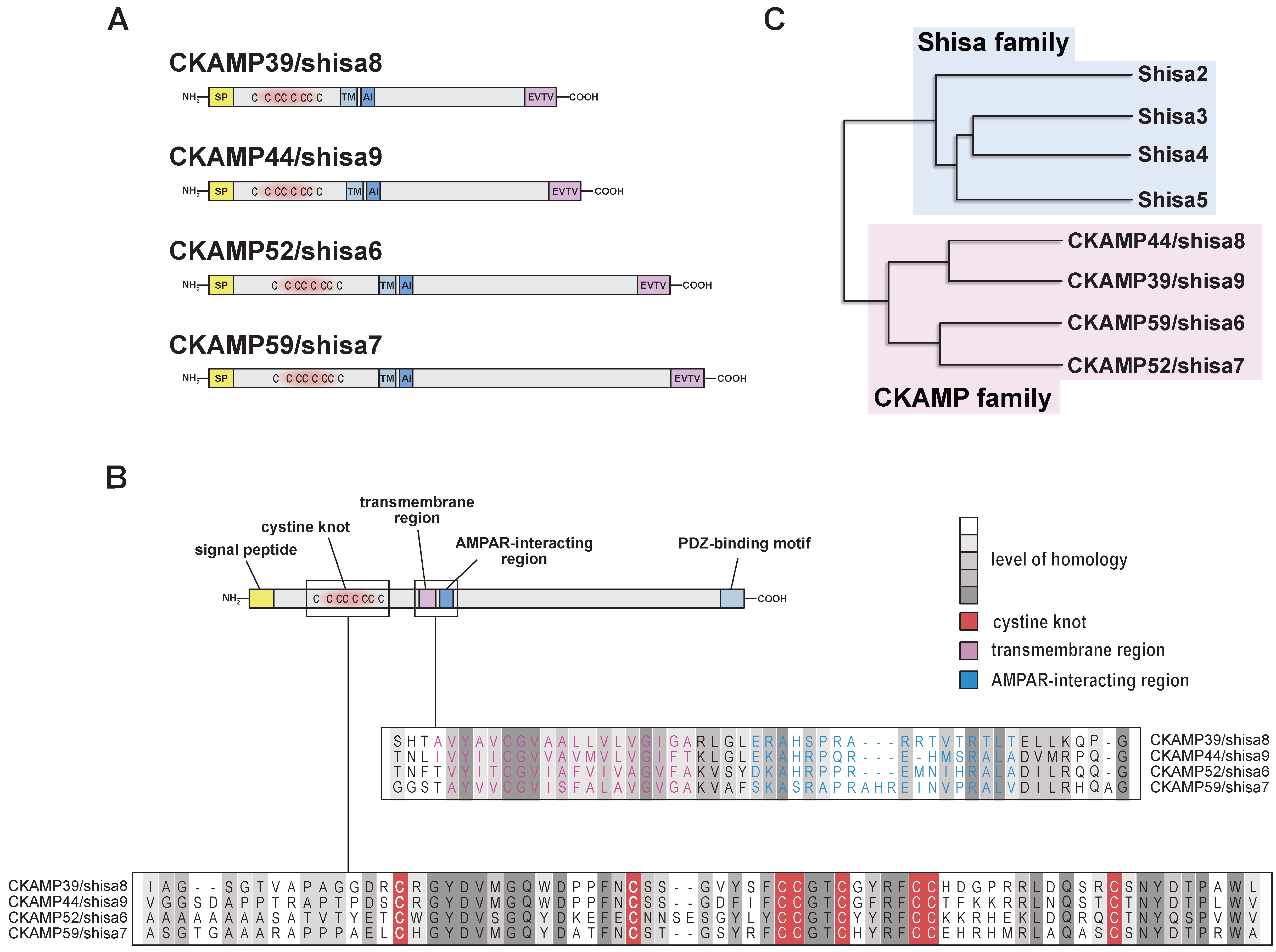
2. Structure of CKAMP Family Members
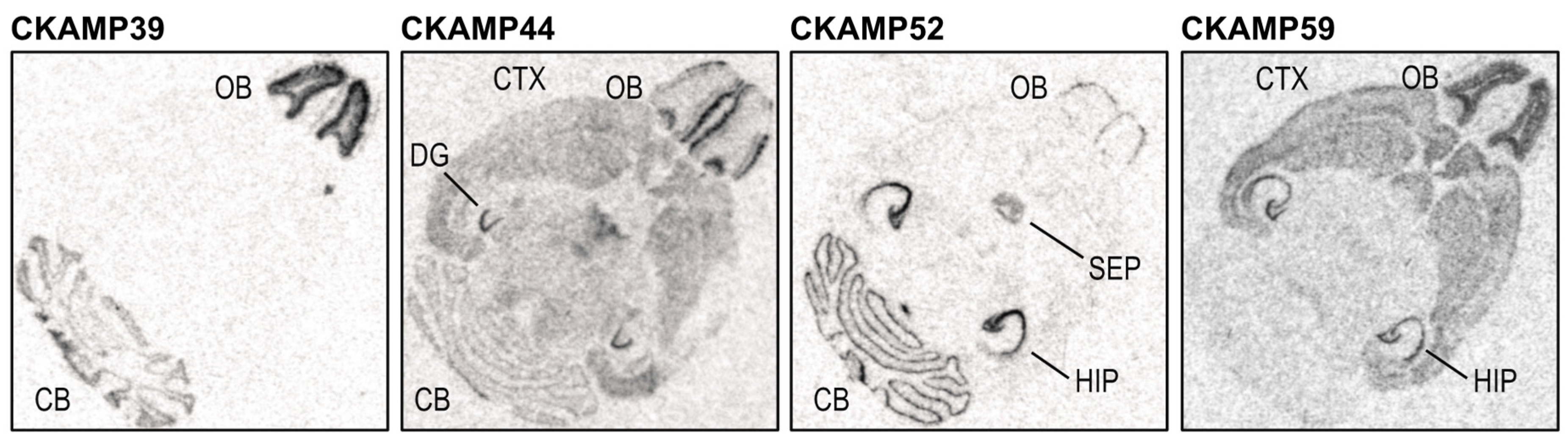
3. Expression of CKAMP Family Members
4. Function of CKAMP Family Members
4.1. Assembly and Surface Trafficking of AMPA Receptors
4.2. Subcellular Localization of AMPA Receptors
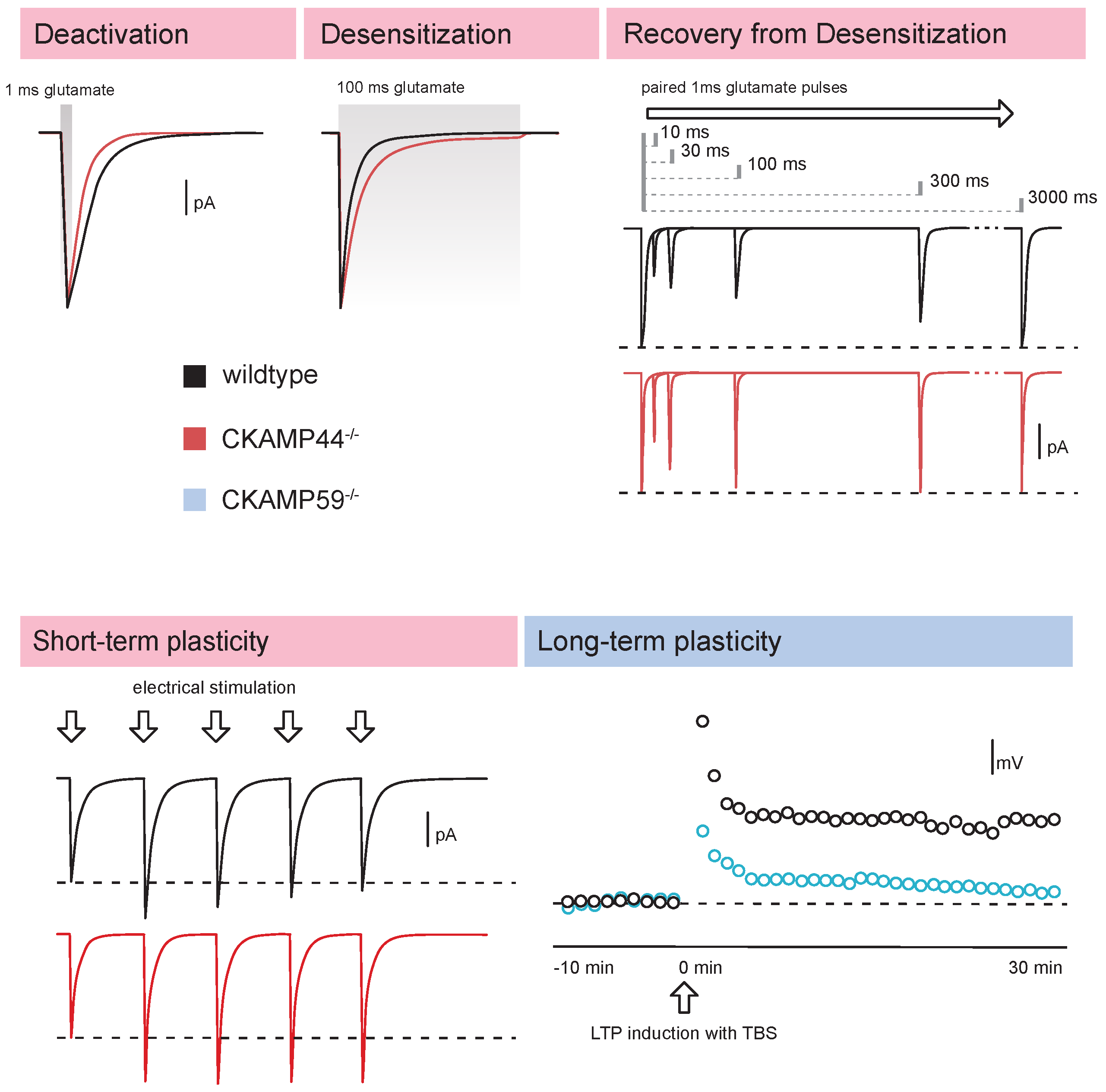
4.3. Gating of AMPA Receptors
4.4. Synaptic AMPA Receptor-Mediated Currents
4.5. Synaptic Short-Term Plasticity
4.6. Long-Term Plasticity
4.7. Behavior
4.8. Other Functions
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| CKAMP | Cysteine-knot AMPA receptor modulating protein |
| CNIH | Cornichon |
| DLG | Discs large homolog |
| ER | endoplasmic reticulum |
| FGF | Fibroblast Growth Factor |
| GRIP1 | Glutamate Receptor Interacting Protein 1 |
| GSG1L | Germ Cell-Specific Gene 1-Like Protein |
| Hz | Hertz |
| LTP | Long-term potentiation |
| MAGI1 | Membrane-Associated Guanylate Kinase Inverted 1 |
| mEPSC | Miniature excitatory postsynaptic current |
| PDZ | Post synaptic density protein (PSD95), Drosophila disc large tumor suppressor (Dlg1), and zonula occludens-1 protein (zo-1) |
| PICK1 | Protein Interacting with C Kinase-1 |
| PSD | Postsynaptic density |
| SAP | Synapse-associated protein |
| TARP | Transmembrane AMPA receptor regulatory proteins |
| Wnt | Wingless/Integrated |
References
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.J.; Myers, S.J.; Dingledine, R. Glutamate Receptor Ion Channels: Structure, Regulation, and Function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwenk, J.; Harmel, N.; Brechet, A.; Zolles, G.; Berkefeld, H.; Muller, C.S.; Bildl, W.; Baehrens, D.; Huber, B.; Kulik, A.; et al. High-Resolution Proteomics Unravel Architecture and Molecular Diversity of Native AMPA Receptor Complexes. Neuron 2012, 74, 621–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwenk, J.; Harmel, N.; Zolles, G.; Bildl, W.; Kulik, A.; Heimrich, B.; Chisaka, O.; Jonas, P.; Schulte, U.; Fakler, B.; et al. Functional proteomics identify cornichon proteins as auxiliary subunits of AMPA receptors. Science 2009, 323, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Shanks, N.F.; Savas, J.N.; Maruo, T.; Cais, O.; Hirao, A.; Oe, S.; Ghosh, A.; Noda, Y.; Greger, I.H.; Yates, J.R., 3rd; et al. Differences in AMPA and kainate receptor interactomes facilitate identification of AMPA receptor auxiliary subunit GSG1L. Cell Rep. 2012, 1, 590–598. [Google Scholar] [CrossRef] [PubMed]
- von Engelhardt, J.; Mack, V.; Sprengel, R.; Kavenstock, N.; Li, K.W.; Stern-Bach, Y.; Smit, A.B.; Seeburg, P.H.; Monyer, H. CKAMP44: A brain-specific protein attenuating short-term synaptic plasticity in the dentate gyrus. Science 2010, 327, 1518–1522. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, E.; von Engelhardt, J. AMPA receptor complex constituents: Control of receptor assembly, membrane trafficking and subcellular localization. Mol. Cell. Neurosci. 2018, 91, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Jacobi, E.; von Engelhardt, J. Diversity in AMPA receptor complexes in the brain. Curr. Opin. Neurobiol. 2017, 45, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Tomita, S. Defined criteria for auxiliary subunits of glutamate receptors. J. Physiol. 2012, 590 Pt 1, 21–31. [Google Scholar] [CrossRef]
- Chen, L.; Bao, S.; Qiao, X.; Thompson, R.F. Impaired cerebellar synapse maturation in waggler, a mutant mouse with a disrupted neuronal calcium channel gamma subunit. Proc. Natl. Acad. Sci. USA 1999, 96, 12132–12137. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Fukaya, M.; Qiao, X.; Sakimura, K.; Watanabe, M.; Kano, M. Impairment of AMPA receptor function in cerebellar granule cells of ataxic mutant mouse stargazer. J. Neurosci. 1999, 19, 6027–6036. [Google Scholar] [CrossRef]
- Tomita, S.; Chen, L.; Kawasaki, Y.; Petralia, R.S.; Wenthold, R.J.; Nicoll, R.A.; Bredt, D.S. Functional studies and distribution define a family of transmembrane AMPA receptor regulatory proteins. J. Cell Biol. 2003, 161, 805–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrow, P.; Khodosevich, K.; Sapir, Y.; Schulmann, A.; Aslam, M.; Stern-Bach, Y.; Monyer, H.; von Engelhardt, J. Auxiliary subunits of the CKAMP family differentially modulate AMPA receptor properties. Elife 2015, 4, e09693. [Google Scholar] [CrossRef] [PubMed]
- Karataeva, A.R.; Klaassen, R.V.; Stroder, J.; Ruiperez-Alonso, M.; Hjorth, J.J.J.; van Nierop, P.; Spijker, S.; Mansvelder, H.D.; Smit, A.B. C-Terminal Interactors of the AMPA Receptor Auxiliary Subunit Shisa9. PLoS ONE 2014, 9, e99280. [Google Scholar] [CrossRef]
- Klaassen, R.V.; Stroeder, J.; Coussen, F.; Hafner, A.S.; Petersen, J.D.; Renancio, C.; Schmitz, L.J.M.; Normand, E.; Lodder, J.C.; Rotaru, D.C.; et al. Shisa6 traps AMPA receptors at postsynaptic sites and prevents their desensitization during synaptic activity. Nat. Commun. 2016, 7, 10682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, L.J.M.; Klaassen, R.V.; Ruiperez-Alonso, M.; Zamri, A.E.; Stroeder, J.; Rao-Ruiz, P.; Lodder, J.C.; van der Loo, R.J.; Mansvelder, H.D.; Smit, A.B.; et al. The AMPA receptor-associated protein Shisa7 regulates hippocampal synaptic function and contextual memory. Elife 2017, 6, e24192. [Google Scholar] [CrossRef] [PubMed]
- Khodosevich, K.; Jacobi, E.; Farrow, P.; Schulmann, A.; Rusu, A.; Zhang, L.; Sprengel, R.; Monyer, H.; von Engelhardt, J. Coexpressed auxiliary subunits exhibit distinct modulatory profiles on AMPA receptor function. Neuron 2014, 83, 601–615. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Grishin, N.V. Unexpected diversity in Shisa-like proteins suggests the importance of their roles as transmembrane adaptors. Cell Signal. 2012, 24, 758–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunde, S.A.; Rademacher, N.; Zieger, H.; Shoichet, S.A. Protein kinase C regulates AMPA receptor auxiliary protein Shisa9/CKAMP44 through interactions with neuronal scaffold PICK1. FEBS Open Bio 2017, 7, 1234–1245. [Google Scholar] [CrossRef] [Green Version]
- Nagano, T.; Takehara, S.; Takahashi, M.; Aizawa, S.; Yamamoto, A. Shisa2 promotes the maturation of somitic precursors and transition to the segmental fate in Xenopus embryos. Development 2006, 133, 4643–4654. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, A.; Nagano, T.; Takehara, S.; Hibi, M.; Aizawa, S. Shisa promotes head formation through the inhibition of receptor protein maturation for the caudalizing factors, Wnt and FGF. Cell 2005, 120, 223–235. [Google Scholar] [CrossRef]
- Molders, A.; Koch, A.; Menke, R.; Klocker, N. Heterogeneity of the astrocytic AMPA-receptor transcriptome. Glia 2018, 66, 2604–2616. [Google Scholar] [CrossRef] [PubMed]
- Schwenk, J.; Baehrens, D.; Haupt, A.; Bildl, W.; Boudkkazi, S.; Roeper, J.; Fakler, B.; Schulte, U. Regional diversity and developmental dynamics of the AMPA-receptor proteome in the mammalian brain. Neuron 2014, 84, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Pandya, N.J.; Koopmans, F.; Castelo-Szekelv, V.; van der Schors, R.C.; Smit, A.B.; Li, K.W. Interaction proteomics reveals brain region-specific AMPA receptor complexes. J. Proteome Res. 2014, 13, 5695–5706. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Aslam, M.; Gollisch, T.; Allen, K.; von Engelhardt, J. CKAMP44 modulates integration of visual inputs in the lateral geniculate nucleus. Nat. Commun. 2018, 9, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellgaard, L.; Helenius, A. Quality control in the endoplasmic reticulum. Nat. Rev. Mol. Cell. Biol. 2003, 4, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Bedoukian, M.A.; Weeks, A.M.; Partin, K.M. Different domains of the AMPA receptor direct stargazin-mediated trafficking and stargazin-mediated modulation of kinetics. J. Biol. Chem. 2006, 281, 23908–23921. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.H.; St-Gelais, F.; Zhang, W.; Tomita, S.; Howe, J.R. Two families of TARP isoforms that have distinct effects on the kinetic properties of AMPA receptors and synaptic currents. Neuron 2007, 55, 890–904. [Google Scholar] [CrossRef] [PubMed]
- Erlenhardt, N.; Yu, H.; Abiraman, K.; Yamasaki, T.; Wadiche, J.I.; Tomita, S.; Bredt, D.S. Porcupine Controls Hippocampal AMPAR Levels, Composition, and Synaptic Transmission. Cell Rep. 2016, 14, 782–794. [Google Scholar] [CrossRef]
- Gill, M.B.; Kato, A.S.; Roberts, M.F.; Yu, H.; Wang, H.; Tomita, S.; Bredt, D.S. Cornichon-2 modulates AMPA receptor-transmembrane AMPA receptor regulatory protein assembly to dictate gating and pharmacology. J. Neurosci. 2011, 31, 6928–6938. [Google Scholar] [CrossRef]
- Gu, X.; Mao, X.; Lussier, M.P.; Hutchison, M.A.; Zhou, L.; Hamra, F.K.; Roche, K.W.; Lu, W. GSG1L suppresses AMPA receptor-mediated synaptic transmission and uniquely modulates AMPA receptor kinetics in hippocampal neurons. Nat. Commun. 2016, 7, 10873. [Google Scholar] [CrossRef] [Green Version]
- Harmel, N.; Cokic, B.; Zolles, G.; Berkefeld, H.; Mauric, V.; Fakler, B.; Stein, V.; Klocker, N. AMPA Receptors Commandeer an Ancient Cargo Exporter for Use as an Auxiliary Subunit for Signaling. PLoS ONE 2012, 7, e30681. [Google Scholar] [CrossRef]
- Milstein, A.D.; Zhou, W.; Karimzadegan, S.; Bredt, D.S.; Nicoll, R.A. TARP subtypes differentially and dose-dependently control synaptic AMPA receptor gating. Neuron 2007, 55, 905–918. [Google Scholar] [CrossRef]
- Penn, A.C.; Williams, S.R.; Greger, I.H. Gating motions underlie AMPA receptor secretion from the endoplasmic reticulum. EMBO J. 2008, 27, 3056–3068. [Google Scholar] [CrossRef] [Green Version]
- von Engelhardt, J. University Mainz, Mainz, Germany. Unpublished Work. 2010. [Google Scholar]
- Lu, W.; Shi, Y.; Jackson, A.C.; Bjorgan, K.; During, M.J.; Sprengel, R.; Seeburg, P.H.; Nicoll, R.A. Subunit composition of synaptic AMPA receptors revealed by a single-cell genetic approach. Neuron 2009, 62, 254–268. [Google Scholar] [CrossRef] [Green Version]
- Heine, M.; Groc, L.; Frischknecht, R.; Beique, J.C.; Lounis, B.; Rumbaugh, G.; Huganir, R.L.; Cognet, L.; Choquet, D. Surface mobility of postsynaptic AMPARs tunes synaptic transmission. Science 2008, 320, 201–205. [Google Scholar] [CrossRef]
- Bats, C.; Groc, L.; Choquet, D. The interaction between Stargazin and PSD-95 regulates AMPA receptor surface trafficking. Neuron 2007, 53, 719–734. [Google Scholar] [CrossRef]
- Constals, A.; Penn, A.C.; Compans, B.; Toulme, E.; Phillipat, A.; Marais, S.; Retailleau, N.; Hafner, A.S.; Coussen, F.; Hosy, E.; et al. Glutamate-Induced AMPA Receptor Desensitization Increases Their Mobility and Modulates Short-Term Plasticity through Unbinding from Stargazin. Neuron 2015, 85, 787–803. [Google Scholar] [CrossRef] [Green Version]
- Herring, B.E.; Shi, Y.; Suh, Y.H.; Zheng, C.Y.; Blankenship, S.M.; Roche, K.W.; Nicoll, R.A. Cornichon proteins determine the subunit composition of synaptic AMPA receptors. Neuron 2013, 77, 1083–1096. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Chavez, A.E.; Mineur, Y.S.; Morimoto-Tomita, M.; Lutzu, S.; Kim, K.S.; Picciotto, M.R.; Castillo, P.E.; Tomita, S. CaMKII Phosphorylation of TARPgamma-8 Is a Mediator of LTP and Learning and Memory. Neuron 2016, 92, 75–83. [Google Scholar] [CrossRef]
- Rouach, N.; Byrd, K.; Petralia, R.S.; Elias, G.M.; Adesnik, H.; Tomita, S.; Karimzadegan, S.; Kealey, C.; Bredt, D.S.; Nicoll, R.A. TARP gamma-8 controls hippocampal AMPA receptor number, distribution and synaptic plasticity. Nat. Neurosci. 2005, 8, 1525–1533. [Google Scholar] [CrossRef]
- Opazo, P.; Labrecque, S.; Tigaret, C.M.; Frouin, A.; Wiseman, P.W.; De Koninck, P.; Choquet, D. CaMKII triggers the diffusional trapping of surface AMPARs through phosphorylation of stargazin. Neuron 2010, 67, 239–252. [Google Scholar] [CrossRef]
- Opazo, P.; Sainlos, M.; Choquet, D. Regulation of AMPA receptor surface diffusion by PSD-95 slots. Curr. Opin. Neurobiol. 2012, 22, 453–460. [Google Scholar] [CrossRef]
- Tokue, M.; Ikami, K.; Mizuno, S.; Takagi, C.; Miyagi, A.; Takada, R.; Noda, C.; Kitadate, Y.; Hara, K.; Mizuguchi, H.; et al. SHISA6 Confers Resistance to Differentiation-Promoting Wnt/beta-Catenin Signaling in Mouse Spermatogenic Stem Cells. Stem Cell Rep. 2017, 8, 561–575. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CKAMP39 | CKAMP44 | CKAMP52 | CKAMP59 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | A2 | A1/2 | A1 | A2 | A1/2 | A1 | A2 | A1/2 | A1 | A2 | A1/2 | |
| Surface expression | ↓ [12] | ↓ [12] | N/A | ↑ [16] | N/A | N/A | - [12] | ↓ [12] | N/A | ↓ [12] | ↓ [12] | N/A |
| Current amplitude | ↓ [12] | ↓ [12] | N/A | ↓ [34] | N/A | N/A | - [12] | ↓ [12] | N/A | - [12] | ↓ [12] | N/A |
| Rise time | - [34] | - [34] | N/A | - [34] | N/A | N/A | - [14] | - [14] | - [14] | - [15] | N/A | N/A |
| Deactivation rate | - [12] | -/↓ [12] | N/A | - [34] | N/A | N/A | -/↓ [12,14] | ↓ [12,14] | ↓ [14] | - [12,15] | - [12] | N/A |
| Desensitization rate | - [12] | ↑ [12] | N/A | ↑ [16] | N/A | N/A | -/↓ [12,14] | -/↑ [12] | ↓ [14] | -/↑ [12,15] | - [12] | N/A |
| Steady-state current amplitude | - [12] | ↓ [12] | N/A | ↓ [16] | N/A | N/A | ↑ [12,14] | ↑ [12] | ↑ [14] | -/↓ [12] | - [12] | N/A |
| Recovery from Desensitization rate | ↓ [12] | ↓ [12] | N/A | ↓ [34] | ↓ [16] | N/A | -/↓ [12] | -/↑ [12] | ↓ [14] | -/↓ [12,15] | - [12] | N/A |
| Rectification | N/A | N/A | N/A | N/A | N/A | N/A | - [14] | N/A | - [14] | - [15] | N/A | N/A |
| Apparent glut affinity | ↑ [12] | ↑ [12] | N/A | ↑ [5,12] | ↑ [12] | N/A | ↑ [12] | ↑ [12] | N/A | N/A | N/A | N/A |
| Apparent CTZ affinity | N/A | N/A | N/A | ↓ [5] | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
| IKA/IGlut | N/A | N/A | N/A | N/A | N/A | ↓ [21] * | N/A | N/A | N/A | N/A | N/A | N/A |
| CK39 | CK44 | CK52 | CK59 | |
|---|---|---|---|---|
| Extrasynaptic (somatic) AMPA receptor-mediated current amplitudes | N/A | ↑ [16,24] | NA | N/A |
| Rise time | N/A | - [16,24] | - [14] | N/A |
| Deactivation rate | N/A | ↓ * [5,16] | - [14] | N/A |
| Desensitization rate | N/A | ↑ * [5,16] | NA/ | N/A |
| Steady-state current amplitude | N/A | ↓ [5,16,24] | N/A | N/A |
| Recovery from desensitization rate | N/A | ↓ [5,16,24] | N/A | N/A |
| Rectification | N/A | ↑ [16] | N/A | N/A |
| Conductance | N/A | ↑ [16] | N/A | N/A |
| Apparent glutamate affinity | N/A | ↑ [5,16] | N/A | N/A |
| CK39 | CK44 | CK52 | CK59 | |
|---|---|---|---|---|
| mEPSC amplitudes | N/A | ↑ [16] | - [14] | - [15] |
| mEPSC frequency | N/A | ↑ [16] | - [14] | - [15] |
| mEPSC rise time | N/A | - [16] | ↑ [14] | - [15] |
| mEPSC decay time | N/A | - [16] | ↑ [14] | ↑ [15] |
| AMPA/NMDA ratio | N/A | ↑ [16,24] | N/A | - [15] |
| Short-term plasticity (↑ = more facilitation) | N/A | ↓ [5,16,24] | ↑ [14] | - [15] |
| Long-term potentiation | N/A | - [16] | N/A | ↑ [15] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
von Engelhardt, J. AMPA Receptor Auxiliary Proteins of the CKAMP Family. Int. J. Mol. Sci. 2019, 20, 1460. https://doi.org/10.3390/ijms20061460
von Engelhardt J. AMPA Receptor Auxiliary Proteins of the CKAMP Family. International Journal of Molecular Sciences. 2019; 20(6):1460. https://doi.org/10.3390/ijms20061460
Chicago/Turabian Stylevon Engelhardt, Jakob. 2019. "AMPA Receptor Auxiliary Proteins of the CKAMP Family" International Journal of Molecular Sciences 20, no. 6: 1460. https://doi.org/10.3390/ijms20061460
APA Stylevon Engelhardt, J. (2019). AMPA Receptor Auxiliary Proteins of the CKAMP Family. International Journal of Molecular Sciences, 20(6), 1460. https://doi.org/10.3390/ijms20061460