Phosphoproteomic Analysis of Two Contrasting Maize Inbred Lines Provides Insights into the Mechanism of Salt-Stress Tolerance


Abstract
:1. Introduction
2. Results
2.1. Physiological Assays of Chang7-2 and Zheng58 in Response to Salt Stress
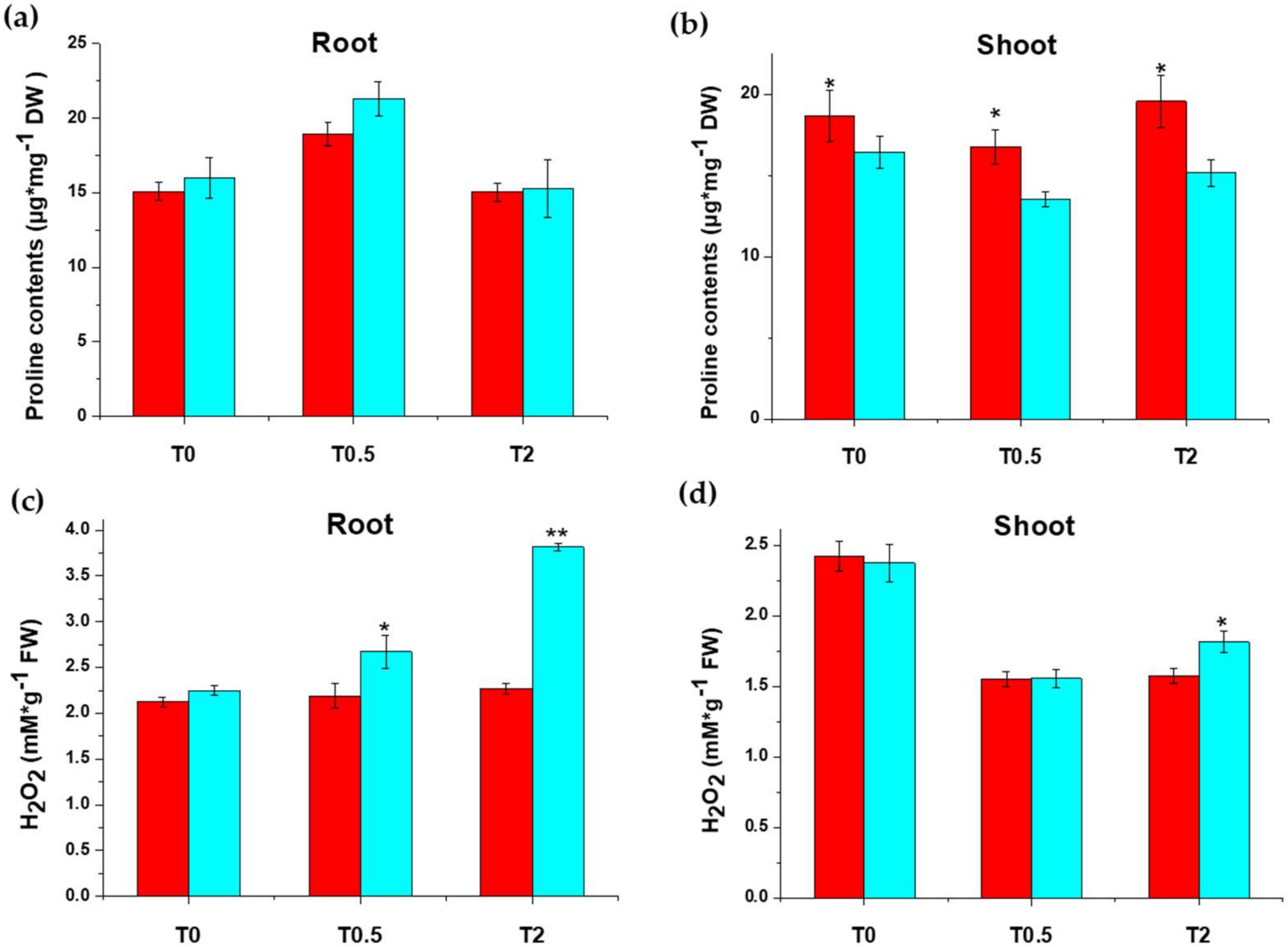
2.1.1. Proline and H2O2 Contents under Salt Treatment
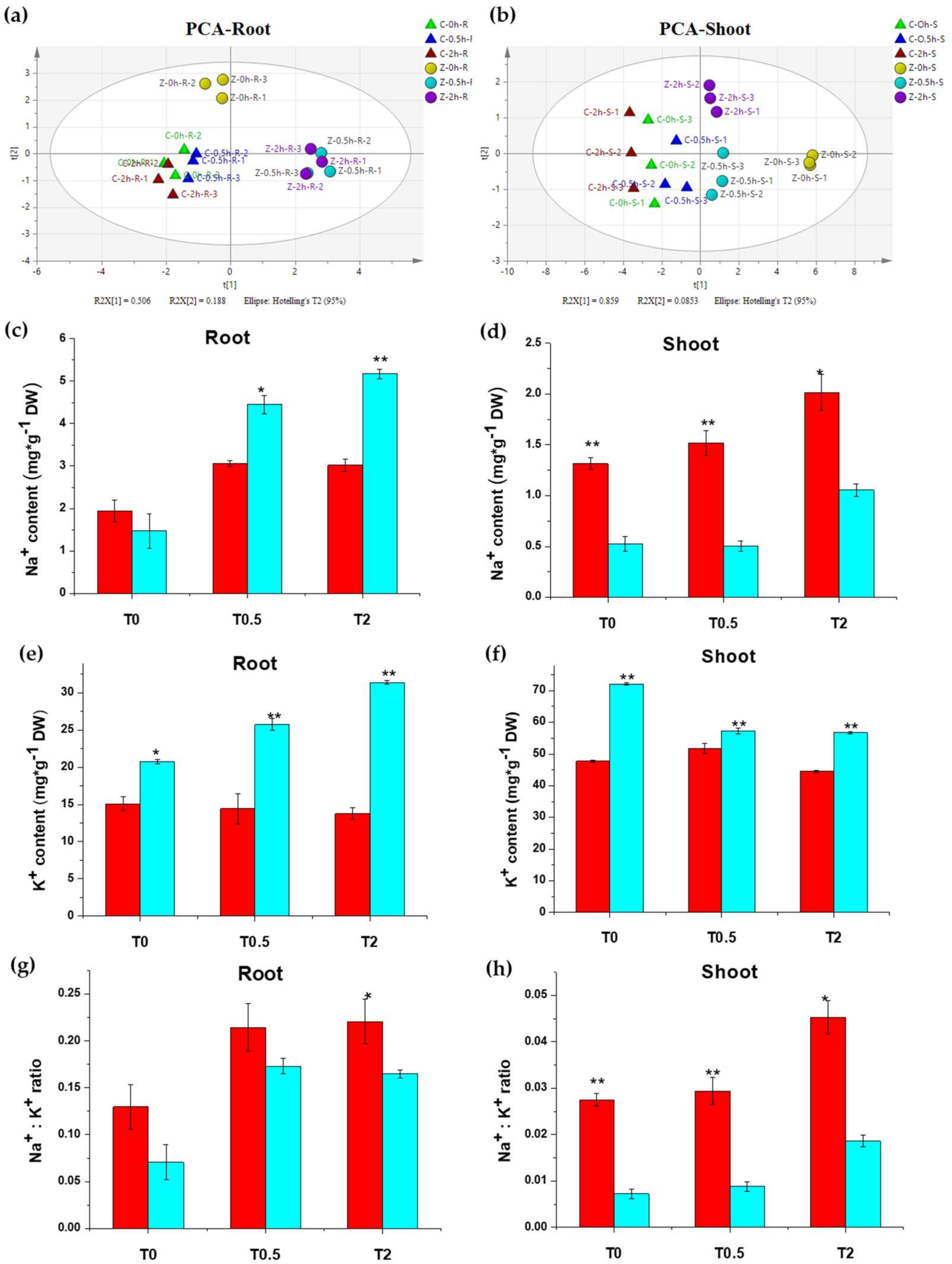
2.1.2. Metal Ions Response to Salt Stress in Chang7-2 and Zheng58
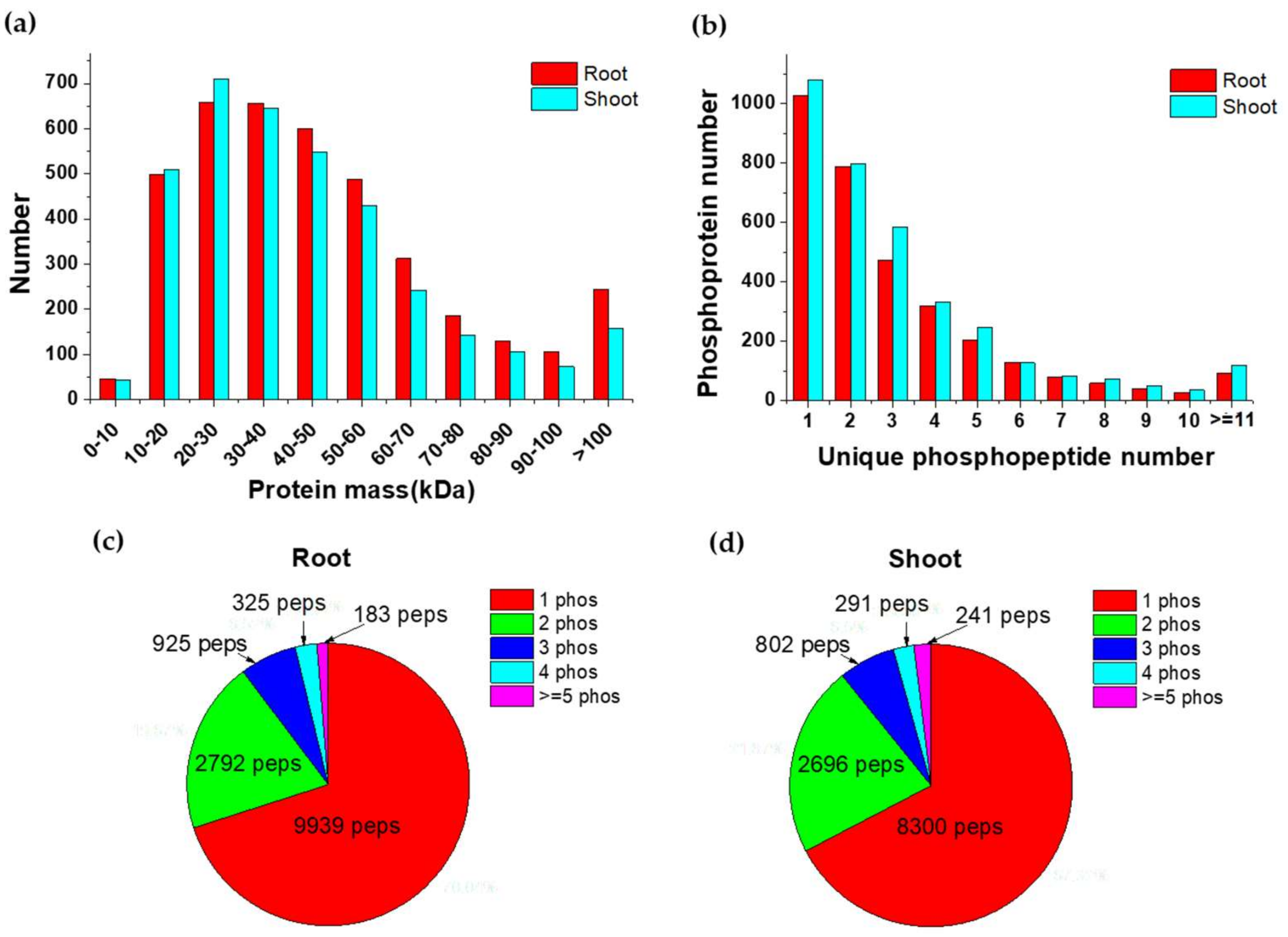
2.2. Overview of Phosphoproteins Identified in Maize Seedlings
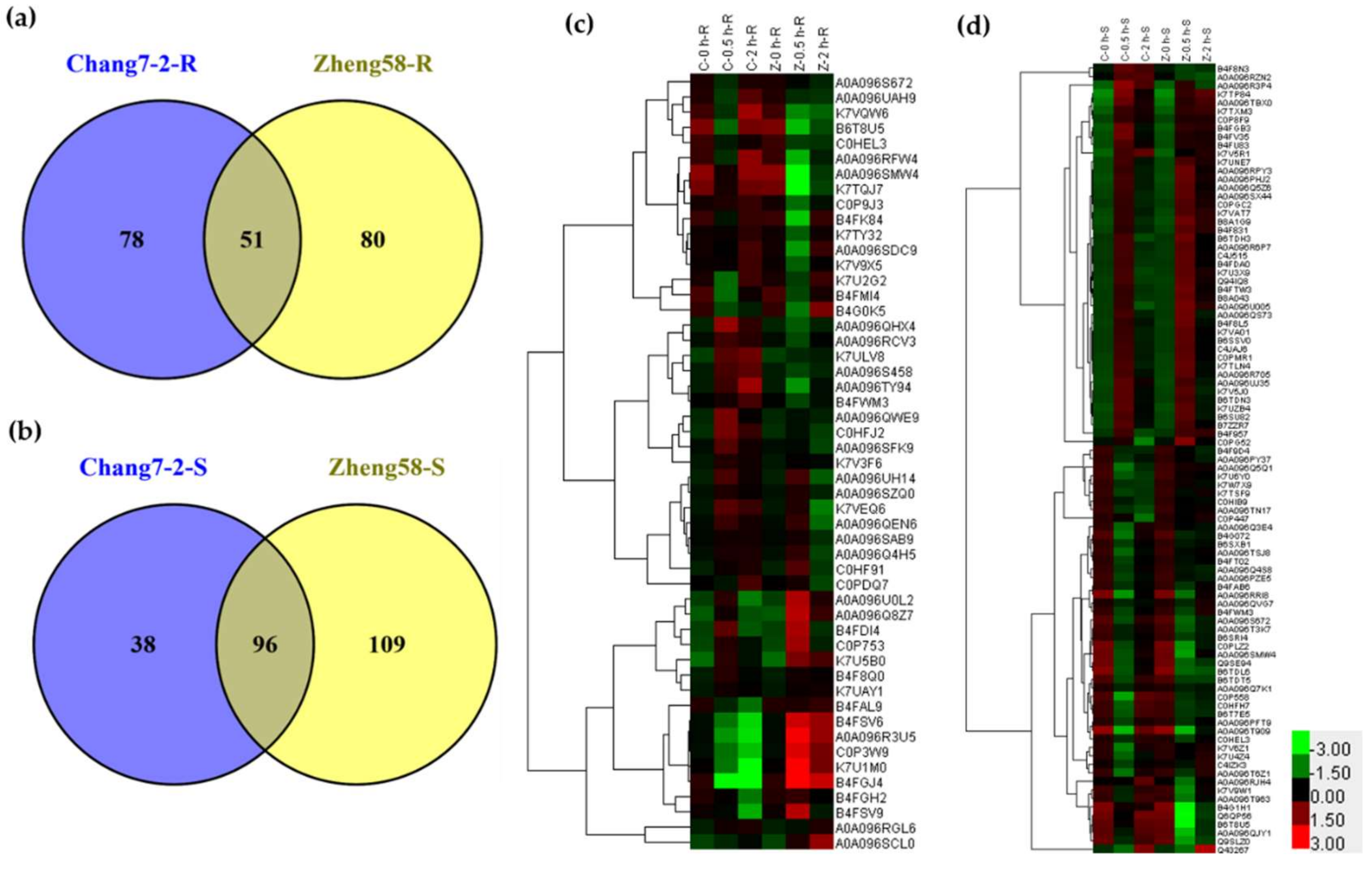
2.3. Differentially Regulated Phosphoproteins in Chang7-2 and Zheng58 in Response to Salt Treatment
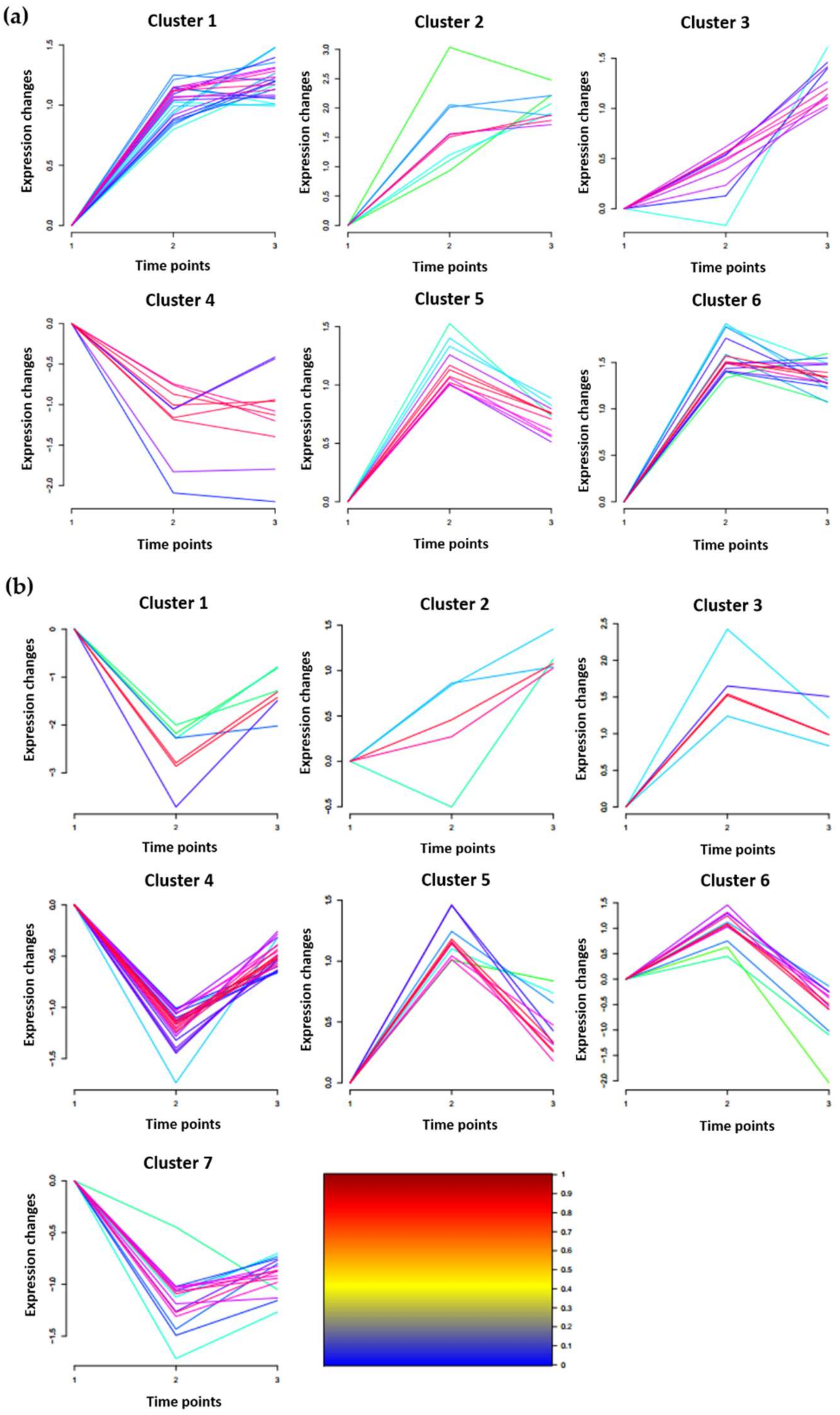
2.4. Clustering Analysis of DRPPs Unique to Chang7-2 or Zheng58 in Maize Roots
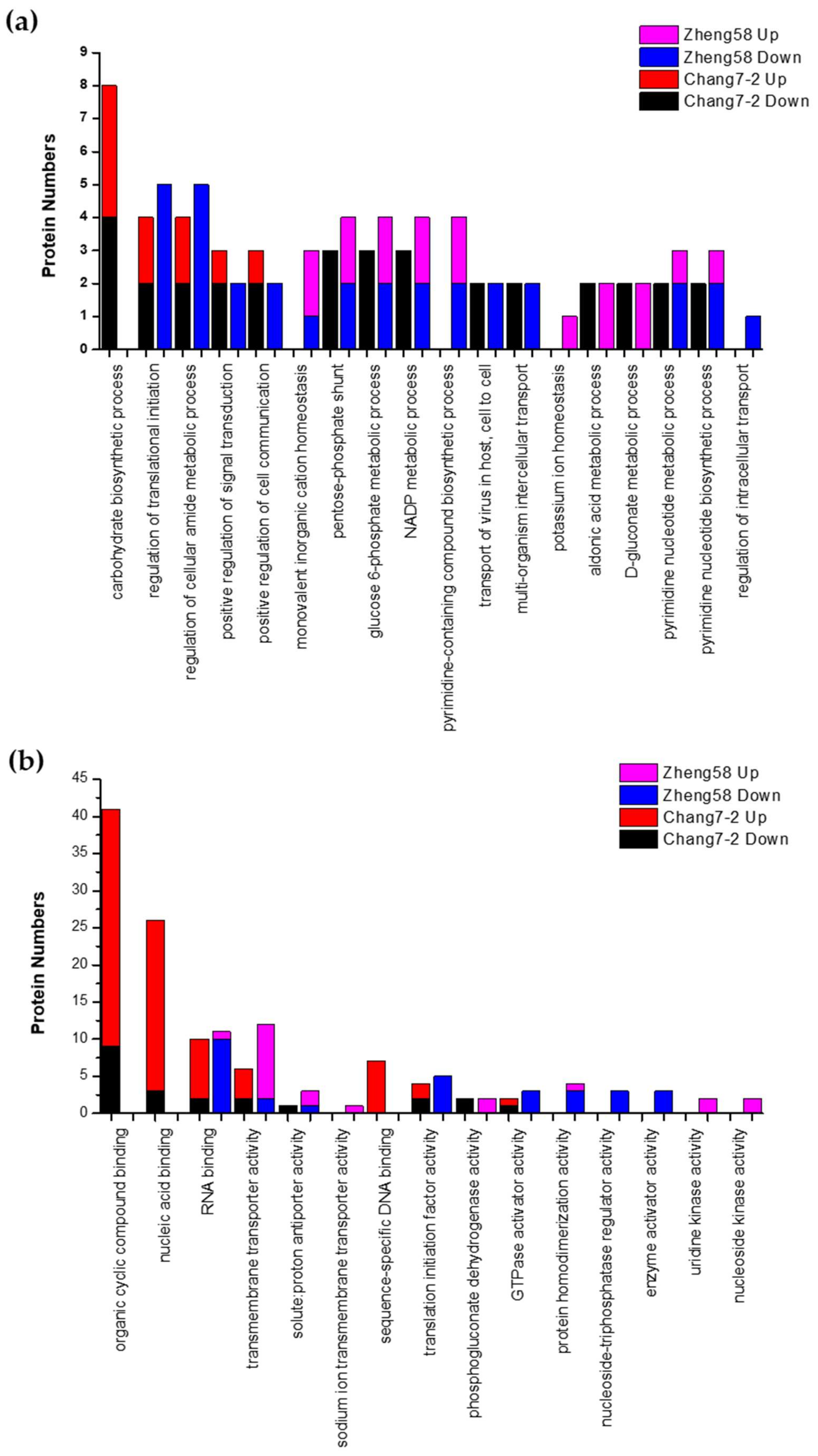
2.5. Gene Ontology Analysis of Salt-Responsive DRPPs in the Two Inbred Lines
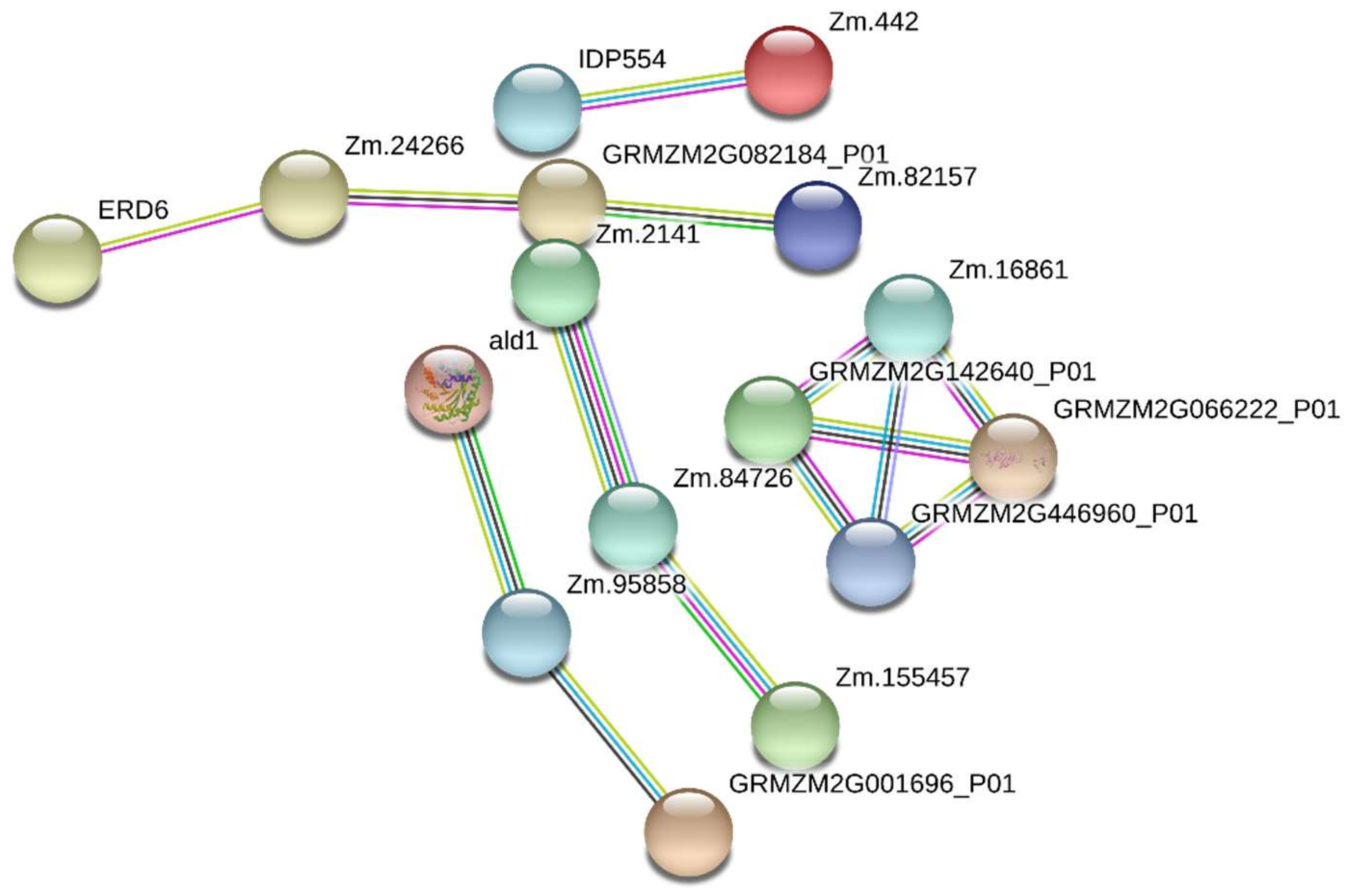
2.6. Protein–Protein Interaction Analysis
3. Discussion
3.1. Physiological Difference between Zheng58 and Chang7-2 under Salt Stress
3.2. Functional Analysis of DRPPs in Roots of Zheng58 and Chang7-2 under Salt Stress
3.2.1. Carbon Metabolism
3.2.2. Glutathione and Ascorbic Acid Metabolism
3.2.3. Glutamate Metabolism
3.2.4. Transport-Related Proteins
3.3. Functional Analysis of DRPPs in Shoots of Zheng58 and Chang7-2 under Salt Stress
3.3.1. Photosynthesis-Related Proteins
3.3.2. Proteins Involved in Carbon Metabolism
3.3.3. Glutathione Metabolism
3.3.4. Plant Hormone Signal Transduction
4. Materials and Methods
4.1. Plant Materials and NaCl Treatment
4.2. Physiological Parameter Measurements
4.3. Protein Extraction and Digestion
4.4. Phosphopeptide Enrichment
4.5. Mass Spectrometry Analysis
4.6. Protein Identification and Quantification
4.7. Bioinformatics Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| bZIP | Basic region-leucine zipper |
| DRPPs | Differential regulated phosphoproteins |
| FDR | false discovery rate |
| GABA | γ-aminobutyric acid |
| GO | Gene ontology |
| ICP-OES | Inductively Coupled Plasma Optical Emission Spectrometry |
| PCA | Principal component analysis |
| PEP1 | Phosphoenolpyruvate carboxylase 1 |
| PIP | plasma membrane intrinsic proteins |
| PSMs | Peptide spectrum matches |
| RILs | Recombinant inbred lines |
| ROS | Reactive oxygen species |
| SOS | Salt Overly Sensitive |
| STRING | Search Tool for the Retrieval of Interacting Genes |
| TCA | Tricarboxylic acid |
References
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.C.; Zhao, G.Y.; Jia, J.Z.; Liu, X.; Kong, X.Y. Molecular characterization of 60 isolated wheat MYB genes and analysis of their expression during abiotic stress. J. Exp. Bot. 2012, 63, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Blumwald, E.; Aharon, G.S.; Apse, M.P. Sodium transport in plant cells. Biochim. Biophys. Acta 2000, 1465, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Galvan-Ampudia, C.S.; Testerink, C. Salt stress signals shape the plant root. Curr. Opin. Plant Biol. 2011, 14, 296–302. [Google Scholar] [CrossRef]
- Kant, S.; Kant, P.; Raveh, E.; Barak, S. Evidence that differential gene expression between the halophyte, Thellungiella halophila, and Arabidopsis thaliana is responsible for higher levels of the compatible osmolyte proline and tight control of Na+ uptake in T. halophila. Plant Cell Environ. 2010, 29, 1220–1234. [Google Scholar] [CrossRef]
- Storey, R.; Jones, R.G.W. Quaternary ammonium compounds in plants in relation to salt resistance. Phytochemistry 1977, 16, 447–453. [Google Scholar] [CrossRef]
- Jouve, L.; Hoffmann, L.; Hausman, J.F. Polyamine, carbohydrate, and proline content changes during salt stress exposure of Aspen (Populus tremula L.): Involvement of oxidation and ssmoregulation metabolism. Plant Biol 2010, 6, 74–80. [Google Scholar]
- Erickson, B.K.; Jedrychowski, M.P.; McAlister, G.C.; Everley, R.A.; Kunz, R.; Gygi, S.P. Evaluating multiplexed quantitative phosphopeptide analysis on a hybrid quadrupole mass filter/linear ion trap/orbitrap mass spectrometer. Anal. Chem. 2015, 87, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Sugiyama, N.; Takahashi, F.; Anderson, J.C.; Ishihama, Y.; Peck, S.C.; Shinozaki, K. Genetics and phosphoproteomics reveal a protein phosphorylation network in the abscisic acid signaling pathway in Arabidopsis thaliana. Sci. Signal 2013, 6, rs8. [Google Scholar] [CrossRef]
- Kline, K.G.; Barrett-Wilt, G.A.; Sussman, M.R. In planta changes in protein phosphorylation induced by the plant hormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 15986–15991. [Google Scholar] [CrossRef] [Green Version]
- Minkoff, B.B.; Stecker, K.E.; Sussman, M.R. Rapid phosphoproteomic effects of abscisic acid (ABA) on wild-type and ABA receptor-deficient A. thaliana mutants. Mol. Cell Proteomics 2015, 14, 1169–1182. [Google Scholar] [CrossRef]
- Wang, X.; Bian, Y.Y.; Cheng, K.; Gu, L.F.; Ye, M.L.; Zou, H.F.; Sun, S.S.M.; He, J.X. A large-scale protein phosphorylation analysis reveals novel phosphorylation motifs and phosphoregulatory networks in Arabidopsis. J. Proteomics 2013, 78, 486–498. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.S.; Wang, X.Y.; Jiang, D.; Dong, S.T. Physiological and proteome studies of maize (Zea mays L.) in response to leaf removal under high plant density. BMC Plant Biol. 2018, 18, 378. [Google Scholar] [CrossRef]
- Chao, Q.; Gao, Z.F.; Wang, Y.F.; Li, Z.; Huang, X.H.; Wang, Y.C.; Mei, Y.C.; Zhao, B.G.; Li, L.; Jiang, Y.B. The proteome and phosphoproteome of maize pollen uncovers fertility candidate proteins. Plant Mol. Biol. 2016, 91, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.L.; Wang, Y.K.; Yang, H.; Wang, W.; Wu, J.Y.; Hu, X.L. Quantitative proteomic analyses identify ABA-related proteins and signal pathways in maize leaves under drought conditions. Front. Plant Sci. 2016, 7, 1827. [Google Scholar] [CrossRef]
- Wang, X.Y.; Shan, X.H.; Wu, Y.; Su, S.Z.; Li, S.P.; Liu, H.K.; Han, J.Y.; Xue, C.M.; Yuan, Y.P. iTRAQ-based quantitative proteomic analysis reveals new metabolic pathways responding to chilling stress in maize seedlings. J. Proteomics 2016, 146, 14–24. [Google Scholar] [CrossRef]
- Hou, Y.X.; Qiu, J.H.; Wang, Y.F.; Li, Z.Y.; Zhao, J.; Tong, X.H.; Lin, H.Y.; Zhang, J. A quantitative proteomic analysis of brassinosteroid-induced protein phosphorylation in rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 514. [Google Scholar] [CrossRef]
- Qiu, J.H.; Hou, Y.X.; Wang, Y.F.; Li, Z.Y.; Zhao, J.; Tong, X.H.; Lin, H.Y.; Wei, X.J.; Ao, H.J.; Zhang, J. A comprehensive proteomic survey of ABA-induced protein phosphorylation in rice (Oryza sativa L.). Int. J. Mol. Sci. 2017, 18, 60. [Google Scholar] [CrossRef]
- Zhong, M.; Li, S.F.; Huang, F.L.; Qiu, J.H.; Zhang, J.; Sheng, Z.H.; Tang, S.Q.; Wei, X.J.; Hu, P.S. The phosphoproteomic response of rice seedlings to cadmium stress. Int. J. Mol. Sci. 2017, 18, 2055. [Google Scholar] [CrossRef]
- Kamal, A.H.M.; Rashid, H.; Sakata, K.; Komatsu, S. Gel-free quantitative proteomic approach to identify cotyledon proteins in soybean under flooding stress. J. Proteomics 2015, 112, 1–13. [Google Scholar] [CrossRef]
- Oh, M.W.; Nanjo, Y.; Komatsu, S. Identification of nuclear proteins in soybean under flooding stress using proteomic technique. Protein Pept. Lett. 2014, 21, 458–467. [Google Scholar] [CrossRef]
- Yin, Y.Q.; Qi, F.; Gao, L.; Rao, S.Q.; Yang, Z.Q.; Fang, W.M. iTRAQ-based quantitative proteomic analysis of dark-germinated soybeans in response to salt stress. RSC Adv. 2018, 8, 17905–17913. [Google Scholar] [CrossRef] [Green Version]
- Ngara, R.; Ndimba, R.; Borch-Jensen, J.; Jensen, O.N.; Ndimba, B. Identification and profiling of salinity stress-responsive proteins in Sorghum bicolor seedlings. J. Proteomics 2012, 75, 4139–4150. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.K.; Cho, S.W.; Kwon, S.J.; Kamal, A.H.M.; Lee, D.G.; Sarker, K.; Lee, M.S.; Xin, Z.G.; Woo, S.H. Proteome characterization of copper stress responses in the roots of sorghum. Biometals 2017, 30, 765–785. [Google Scholar] [CrossRef]
- Pi, E.; Qu, L.Q.; Hu, J.W.; Huang, Y.Y.; Qiu, L.J.; Lu, H.F.; Jiang, B.; Liu, C.; Peng, T.T.; Zhao, Y.; et al. Mechanisms of soybean roots’ tolerances to salinity revealed by proteomic and phosphoproteomic comparisons between two cultivars. Mol. Cell Proteomics 2016, 15, 266–288. [Google Scholar] [CrossRef]
- Gao, L.; Yan, X.; Li, X.; Guo, G.; Hu, Y.; Ma, W.; Yan, Y. Proteome analysis of wheat leaf under salt stress by two-dimensional difference gel electrophoresis (2D-DIGE). Phytochemistry 2011, 72, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.Z.; Wu, D.D.; Liu, J.; Li, D.T.; Xu, C.Y.; Li, S.; Li, P.; Zhang, H.; Liu, X.; Jiang, C.; et al. Proteomic analysis of seedling roots of two maize inbred lines that differ significantly in the salt stress response. PLoS ONE 2015, 10, e0116697. [Google Scholar] [CrossRef] [PubMed]
- Macek, B.; Mann, M.; Olsen, J.V. Global and site-specific quantitative phosphoproteomics: Principles and applications. Annu. Rev. Pharmacool. Toxicol. 2009, 49, 199–221. [Google Scholar] [CrossRef]
- Morandell, S.; Stasyk, T.; Grosstessner-Hain, K.; Roitinger, E.; Mechtler, K.; Bonn, G.K.; Huber, L.A. Phosphoproteomics strategies for the functional analysis of signal transduction. Proteomics 2006, 6, 4047–4056. [Google Scholar] [CrossRef]
- Fujii, H.; Chinnusamy, V.; Rodrigues, A.; Rubio, S.; Antoni, R.; Park, S.Y.; Cutler, S.R.; Sheen, J.; Rodriguez, P.L.; Zhu, J.K. In vitro reconstitution of an abscisic acid signalling pathway. Nature 2009, 462, 660–664. [Google Scholar] [CrossRef]
- Fujii, H.; Zhu, J.K. Osmotic stress signaling via protein kinases. Cell. Mol. Life Sci. 2012, 69, 3165–3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.T.; Ding, Y.L.; Yang, S.H. Molecular regulation of CBF signaling in cold acclimation. Trends Plant Sci. 2018, 23, 623–637. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Zolnierowicz, S. Type 2A protein phosphatase, the complex regulator of numerous signaling pathways. Biochem. Pharmacol. 2000, 60, 1225–1235. [Google Scholar] [CrossRef]
- Liu, J.P.; Ishitani, M.; Halfter, U.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000, 97, 3730–3734. [Google Scholar] [CrossRef]
- Halfter, U.; Ishitani, M.; Zhu, J.K. The Arabidopsis SOS2 protein kinase physically interacts with and is activated by the calcium-binding protein SOS3. Proc. Natl. Acad. Sci. USA 2000, 97, 3735–3740. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.P.; Zhu, J.K. A calcium sensor homolog required for plant salt tolerance. Science 1998, 280, 1943–1945. [Google Scholar] [CrossRef]
- Ishitani, M.; Liu, J.P.; Halfter, U.; Kim, C.S.; Shi, W.M.; Zhu, J.K. SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding. Plant Cell 2000, 12, 1667–1677. [Google Scholar] [CrossRef]
- Knight, H.; Trewavas, A.J.; Knight, M.R. Calcium signalling in Arabidopsis thaliana responding to drought and salinity. Plant J. 2010, 12, 1067–1078. [Google Scholar] [CrossRef]
- MartiNez-Atienza, J.; Jiang, X.Y.; Garciadeblas, B.; Mendoza, I.; Zhu, J.K.; Pardo, J.M.; Quintero, F.J. Conservation of the salt overly sensitive pathway in rice. Plant Physiol. 2007, 143, 1001–1012. [Google Scholar] [CrossRef]
- Wang, M.Y.; Gu, D.; Liu, T.S.; Wang, Z.Q.; Guo, X.Y.; Hou, W.; Bai, Y.F.; Chen, X.P.; Wang, G.Y. Overexpression of a putative maize calcineurin B-like protein in Arabidopsis confers salt tolerance. Plant Mol. Biol. 2007, 65, 733–746. [Google Scholar] [CrossRef]
- Xu, Q.; Fu, H.H.; Gupta, R.; Luan, S. Molecular characterization of a tyrosine-specific protein phosphatase encoded by a stress-responsive gene in Arabidopsis. Plant Cell 1998, 10, 849–857. [Google Scholar] [CrossRef]
- Geilfus, C.M.; Zoerb, C.; Mühling, K.H. Salt stress differentially affects growth-mediating β-expansins in resistant and sensitive maize (Zea mays L.). Plant Physiol. Biochem. 2010, 48, 993–998. [Google Scholar] [CrossRef]
- Muhammad, F.; Mubshar, H.; Abdul, W.; Kadambot, H.M.S. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar]
- Katerji, N.; van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M. Salinity effect on crop development and yield, analysis of salt tolerance according to several classification methods. Agric. Water Manag. 2003, 62, 37–66. [Google Scholar] [CrossRef]
- Nakagami, H.; Sugiyama, N.; Mochida, K.; Daudi, A.; Yoshida, Y.; Toyoda, T.; Tomita, M.; Ishihama, Y.; Shirasu, K. Large-scale comparative phosphoproteomics identifies conserved phosphorylation sites in plants. Plant Physiol. 2010, 153, 1161–1174. [Google Scholar] [CrossRef]
- Bi, Y.D.; Wang, H.X.; Lu, T.C.; Li, X.H.; Shen, Z.O.; Chen, Y.B.; Wang, B.C. Large-scale analysis of phosphorylated proteins in maize leaf. Planta 2011, 233, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.L.C.; Geilfus, C.M.; Carpentier, S.C. Genotype-specific growth and proteomic responses of maize toward salt stress. Front. Plant Sci. 2018, 9, 661. [Google Scholar] [CrossRef]
- Luo, M.J.; Zhao, Y.X.; Wang, Y.D.; Shi, Z.; Zhang, P.P.; Zhang, Y.X.; Song, W.; Zhao, J.R. Comparative proteomics of contrasting maize genotypes provides insights into salt-stress tolerance mechanisms. J. Proteome Res. 2018, 17, 141–153. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.M.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14 (Suppl 1), S165–S183. [Google Scholar] [CrossRef] [PubMed]
- Girousse, C.; Bournoville, R.; Bonnemain, J.L. Water deficit-induced changes in concentrations in proline and some other amino acids in the Phloem Sap of Alfalfa. Plant Physiol. 1996, 111, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Ueda, A.; Shi, W.M.; Sanmiya, K.; Shono, M.; Takabe, T. Functional analysis of salt-inducible proline transporter of Barley roots. Plant Cell Physiol. 2001, 42, 1282–1289. [Google Scholar] [CrossRef]
- Matsushita, N.; Matoh, T. Function of the shoot base of salt-tolerant reed (Phragmites communis Trinius) plants for Na+ exclusion from the shoots. Soil Sci. Plant Nutr. 1992, 38, 565–571. [Google Scholar] [CrossRef]
- Zhang, M.; Cao, Y.B.; Wang, Z.P.; Wang, Z.Q.; Shi, J.P.; Liang, X.Y.; Song, W.B.; Chen, Q.J.; Lai, J.S.; Jiang, C.F. A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na+ exclusion and salt tolerance in maize. New Phytol. 2017, 217, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Chen, X.L.; Du, D.; Zheng, W.; Hu, L.Q.; Yang, H.; Cheng, J.Q.; Gong, M. MetaboGroupS: A group entropy-based web platform for evaluating normalization methods in blood metabolomics data from maintenance hemodialysis patients. Anal. Chem. 2018, 90, 11124–11130. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Schnarrenberger, C.; Oeser, A.; Tolbert, N.E. Two isoenzymes each of glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase in spinach leaves. Arch. Biochem. Biophys. 1973, 154, 438–448. [Google Scholar] [CrossRef]
- Diaz, V.P.; Wolff, T.; Markovic, J.; Pallardó, F.V.; Foyer, C.H. A nuclear glutathione cycle within the cell cycle. Biochem. J. 2010, 431, 169–178. [Google Scholar] [Green Version]
- Lea, P.J.; Leegood, R.C. Plant biochemistry and molecular biology. Plant Mol. Biol. 1993, 19, 169–191. [Google Scholar]
- Edwards, R.; Dixon, D.P.; Walbot, V. Plant glutathione S -transferases: Enzymes with multiple functions in sickness and in health. Trends Plant Sci. 2000, 5, 193–198. [Google Scholar] [CrossRef]
- Lara, H.; Vogeli, P.; Stoll, P.; Kramer, S.S.; Stranzinger, G.; Neuenschwander, S. Intragenic deletion in the gene encoding l-gulonolactone oxidase causes vitamin C deficiency in pigs. Mamm. Genome 2004, 15, 323–333. [Google Scholar]
- Hyun, T.K.; Eom, S.H.; Jeun, Y.C.; Han, S.H.; Kim, J.S. Identification of glutamate decarboxylases as a γ-aminobutyric acid (GABA) biosynthetic enzyme in soybean. Ind. Crops Prod. 2013, 49, 864–870. [Google Scholar] [CrossRef]
- Hare, P.D.; Cress, W.A.; Staden, J.V. Proline synthesis and degradation: A model system for elucidating stress-related signal transduction. J. Exp. Bot. 1999, 50, 413–434. [Google Scholar] [CrossRef]
- Demiral, T.; Türkan, I. Exogenous glycinebetaine affects growth and proline accumulation and retards senescence in two rice cultivars under NaCl stress. Environ. Exp. Bot. 2006, 56, 72–79. [Google Scholar] [CrossRef]
- Hmida-Sayari, A.; Gargouri-Bouzid, R.; Bidani, A.; Jaoua, L.; Savouré, A.; Jaoua, S. Overexpression of Δ1-pyrroline-5-carboxylate synthetase increases proline production and confers salt tolerance in transgenic potato plants. Plant Sci. 2005, 169, 746–752. [Google Scholar] [CrossRef]
- Khatun, S.; Flowers, T.J. Effects of salinity on seed set in rice. Plant Cell Environ. 2010, 18, 61–67. [Google Scholar] [CrossRef]
- Li, J.; Long, Y.; Qi, G.N.; Li, J.; Xu, Z.J.; Wu, W.H.; Wang, Y. The Os-AKT1 channel is critical for K+ uptake in rice roots and is modulated by the rice CBL1-CIPK23 complex. Plant Cell 2014, 26, 3387–3402. [Google Scholar] [CrossRef]
- Wu, G.Q.; Wang, J.L.; Feng, R.J.; Li, S.J.; Wang, C.M. iTRAQ-based comparative proteomic analysis provides insights into molecular mechanisms of salt tolerance in Sugar Beet (Beta vulgaris L.). Int. J. Mol. Sci. 2018, 19, 3866. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Pandey, G.K.; Tuteja, N. Calcium- and salt-stress signaling in plants: Shedding light on SOS pathway. Arch. Biochem. Biophys. 2008, 471, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Barrena, M.J.; Martínez-Ripoll, M.; Zhu, J.K.; Albert, A. The structure of the Arabidopsis thaliana SOS3: Molecular mechanism of sensing calcium for salt stress response. J. Mol. Biol. 2005, 345, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Plants use calcium to resolve salt stress. Trends Plant Sci. 1998, 3, 411–412. [Google Scholar] [CrossRef]
- Shigaki, T.; Rees, I.; Nakhleh, L.; Hirschi, K.D. Identification of three distinct phylogenetic groups of CAX cation/proton antiporters. J. Mol. Evol. 2006, 63, 815–825. [Google Scholar] [CrossRef]
- Melzer, E.; O’Leary, M.H. Anapleurotic CO2 fixation by phosphoenolpyruvate carboxylase in C3 plants. Plant Physiol. 1987, 84, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Leegood, R.C.; Ap, R.T. Phosphoenolpyruvate carboxykinase and gluconeogenesis in cotyledons of Cucurbita pepo. Biochim. Biophys. Acta 1978, 524, 207–218. [Google Scholar] [CrossRef]
- Roman, E. Studies on the Role of UDP-Glucose Dehydrogenase in Polysaccharide Biosynthesis. Ph.D. Thesis, Acta Universitatis Upsaliensis, Uppsala, Sweden, 2004. [Google Scholar]
- Arora, M.; Kaushik, A.; Rani, N.; Kaushik, C.P. Effect of cyanobacterial exopolysaccharides on salt stress alleviation and seed germination. J. Environ. Biol. 2010, 31, 701–704. [Google Scholar]
- Ghanem, M.E.; Han, R.M.; Classen, B.; Quetin-Leclerq, J.L.; Mahy, G.; Ruan, C.J.; Qin, P.; Pérez-Alfocea, F.; Lutts, S. Mucilage and polysaccharides in the halophyte plant species Kosteletzkya virginica: Localization and composition in relation to salt stress. J. Plant Physiol. 2010, 167, 382–392. [Google Scholar] [CrossRef]
- Brocard, I.M.; Lynch, T.J.; Finkelstein, R.R. Regulation and role of the Arabidopsis abscisic acid-insensitive 5 gene in abscisic acid, sugar, and stress response. Plant Physiol. 2002, 129, 1533–1543. [Google Scholar] [CrossRef]
- Setter, T.L.; Yan, J.B.; Warburton, M.; Ribaut, J.M.; Xu, Y.B.; Sawkins, M.; Buckler, E.S.; Zhang, Z.W.; Gore, M.A. Genetic association mapping identifies single nucleotide polymorphisms in genes that affect abscisic acid levels in maize floral tissues during drought. J. Exp. Bot. 2011, 62, 701–716. [Google Scholar] [CrossRef]
- Singh, K.; Singla-Pareek, S.L.; Pareek, A. Dissecting out the crosstalk between salinity and hormones in roots of Arabidopsis. OMICS 2011, 15, 913–924. [Google Scholar] [CrossRef]
- Knight, H.; Zarka, D.G.; Okamoto, H.; Thomashow, M.F.; Knight, M.R. Abscisic acid induces CBF gene transcription and subsequent induction of cold-regulated genes via the CRT promoter element. Plant Physiol. 2004, 135, 1710–1717. [Google Scholar] [CrossRef]
- Zhao, B.Y.; Hu, Y.F.; Li, J.J.; Yao, X.; Liu, K.D. BnaABF2, a bZIP transcription factor from rapeseed (Brassica napus L.), enhances drought and salt tolerance in transgenic Arabidopsis. Bot. Stud. 2016, 57, 12. [Google Scholar] [CrossRef]
- Xu, J.; Li, H.D.; Chen, L.Q.; Wang, Y.; Liu, L.L.; He, L.; Wu, W.H. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef]
- Yin, B.J.; Li, T.T.; Zhang, S.R.; Li, Z.; He, P.L. Sensitive analysis of 33 free amino acids in serum, milk, and muscle by ultra-high performance liquid chromatography-quadrupole-orbitrap high resolution mass spectrometry. Food Anal. Methods 2016, 9, 2814–2823. [Google Scholar] [CrossRef]
- Carpentier, S.C.; Witters, E.; Laukens, K.; Deckers, P.; Swennen, R.; Panis, B. Preparation of protein extracts from recalcitrant plant tissues: An evaluation of different methods for two-dimensional gel electrophoresis analysis. Proteomics 2005, 5, 2497–2507. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Annotation | p-Value | C-0.5h | C-2h | Z-0.5h | Z-2h |
|---|---|---|---|---|---|---|
| fc 1 | fc | fc 2 | fc | |||
| Carbon Metabolism | ||||||
| B4FSV6 | 6-phosphogluconate dehydrogenase 2 | 0.0095 | 0.37 | 0.31 | 1.88 | 2.05 |
| B4FGJ4 | Pyruvate dehydrogenase | 1.1 × 10−6 | 0.08 | 0.06 | 3.60 | 2.02 |
| C0P3W9 | phosphoenolpyruvate carboxykinase | 0.0124 | 0.38 | 0.38 | 2.16 | 1.58 |
| Glutathione and Ascorbic Acid Metabolism | ||||||
| B4FK84 | Glutathione S-transferase 3 | 0.0427 | 0.47 | 1.59 | 0.04 | 0.53 |
| K7U1M0 | l-gulonolactone oxidase-like | 0.0054 | 0.32 | 0.19 | 2.57 | 1.37 |
| Glutamate Metabolism | ||||||
| B4F972 | Glutamate decarboxylase | 0.0203 | - | - | 0.30 | 0.80 |
| A0A096RAH5 | glutamate synthase | 0.0002 | 1.01 | 0.79 | 2.00 | 1.53 |
| Transport-Related Proteins | ||||||
| K7V3Z4 | Potassium channel AKT1 | 0.0092 | 1.28 | 0.99 | 2.06 | 1.39 |
| W5U5W2 | high-affinity potassium transporter | 0.0095 | 1.07 | 0.95 | 2.49 | 0.78 |
| A0A096Q7K1 | Sodium/hydrogen exchanger | 4.29 × 10−6 | 1.55 | 1.64 | 2.13 | 0.71 |
| B4F910 | Calcium/proton exchanger CAX1-like protein | 1.01 × 10−5 | 1.17 | 0.99 | 1.16 | 0.35 |
| Accession | Annotation | p-Value | C-0.5h | C-2h | Z-0.5h | Z-2h |
|---|---|---|---|---|---|---|
| fc | fc | fc | fc | |||
| Photosynthesis-Related Proteins | ||||||
| B6TH55 | Photosystem I reaction center subunit IV A | 2.45 × 10−8 | 1.56 | 1.21 | 2.13 | 1.60 |
| P24993 | Photosystem II reaction center protein H | 8.67 × 10−8 | 1.32 | 1.16 | 0.53 | 2.44 |
| Carbon Metabolism | ||||||
| Q43267 | phosphoenolpyruvate carboxylase 1 (PEP1) | 8.42 × 10−9 | 0.51 | 4.72 | 0.39 | 6.46 |
| Q9SLZ0 | Phosphoenolpyruvate carboxykinase | 1.65 × 10−11 | 0.26 | 0.90 | 0.08 | 0.27 |
| A0A096T909 | UDP-glucose 6-dehydrogenase | 8.58 × 10−12 | 0.05 | 0.77 | 0.04 | 0.26 |
| Glutamate Metabolism | ||||||
| B4FTF8 | glutathione S-transferase | 1.12 × 10−7 | 1.18 | 1.40 | 2.05 | 1.74 |
| B6T7H0 | glutathione S-transferase 6 | 9.47 × 10−7 | 1.10 | 0.75 | 2.07 | 1.25 |
| Plant Hormone Signal Transduction | ||||||
| B4F831 | Abscisic acid-insensitive 5-like protein | 9.29 × 10−9 | 2.05 | 1.52 | 2.36 | 2.50 |
| B6TNQ7 | Ninja-family protein 6 | 6.63 × 10−9 | 1.93 | 1.88 | 2.20 | 2.53 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Bai, X.; Jiang, C.; Li, Z. Phosphoproteomic Analysis of Two Contrasting Maize Inbred Lines Provides Insights into the Mechanism of Salt-Stress Tolerance. Int. J. Mol. Sci. 2019, 20, 1886. https://doi.org/10.3390/ijms20081886
Zhao X, Bai X, Jiang C, Li Z. Phosphoproteomic Analysis of Two Contrasting Maize Inbred Lines Provides Insights into the Mechanism of Salt-Stress Tolerance. International Journal of Molecular Sciences. 2019; 20(8):1886. https://doi.org/10.3390/ijms20081886
Chicago/Turabian StyleZhao, Xiaoyun, Xue Bai, Caifu Jiang, and Zhen Li. 2019. "Phosphoproteomic Analysis of Two Contrasting Maize Inbred Lines Provides Insights into the Mechanism of Salt-Stress Tolerance" International Journal of Molecular Sciences 20, no. 8: 1886. https://doi.org/10.3390/ijms20081886
APA StyleZhao, X., Bai, X., Jiang, C., & Li, Z. (2019). Phosphoproteomic Analysis of Two Contrasting Maize Inbred Lines Provides Insights into the Mechanism of Salt-Stress Tolerance. International Journal of Molecular Sciences, 20(8), 1886. https://doi.org/10.3390/ijms20081886