Acyl-CoA-Binding Domain-Containing 3 (ACBD3; PAP7; GCP60): A Multi-Functional Membrane Domain Organizer


Abstract
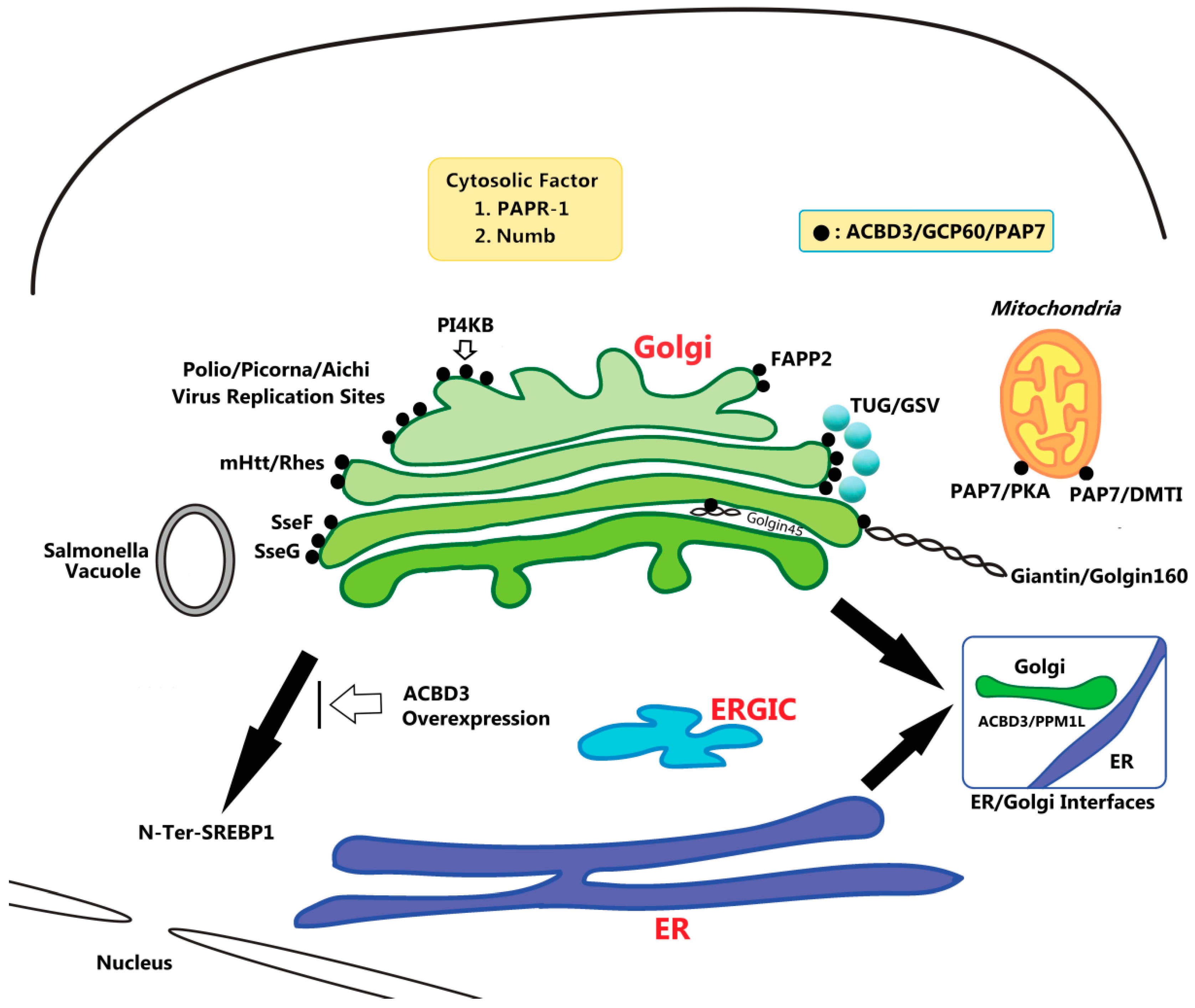
:1. Introduction
2. ACBD3 in Steroidogenesis
3. ACBD3 in Lipid Metabolism and Metabolic Homoeostasis
4. ACBD3 in Regulation of Ceramide and Glucosylceramide Transport
5. ACBD3 in Golgi Structure Maintenance
6. ACBD3 in Iron Uptake
7. ACBD3 Is Exploited by Viral and Bacterial Proteins to Promote Their Replication
8. ACBD3 in Intracellular Retention of GLUT4 Storage Vesicles (GSV)
9. ACBD3 in Huntington’s Disease (HD)
10. ACBD3 in Cancer
11. Conclusions
Funding
Conflicts of Interest
References
- Warren, G.; Malhotra, V. The organisation of the Golgi apparatus. Curr. Opin. Cell Biol. 1998, 10, 493–498. [Google Scholar] [CrossRef]
- Traub, L.M.; Kornfeld, S. The trans-Golgi network: A late secretory sorting station. Curr. Opin. Cell Biol. 1997, 9, 527–533. [Google Scholar] [CrossRef]
- Pulvirenti, T.; Giannotta, M.; Capestrano, M.; Capitani, M.; Pisanu, A.; Polishchuk, R.S.; San Pietro, E.; Beznoussenko, G.V.; Mironov, A.A.; Turacchio, G.; et al. A traffic-activated Golgi-based signalling circuit coordinates the secretory pathway. Nat. Cell Biol. 2008, 10, 912–922. [Google Scholar] [CrossRef]
- Machamer, C.E. The Golgi complex in stress and death. Front. Neurosci. 2015, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Preisinger, C.; Korner, R.; Wind, M.; Lehmann, W.D.; Kopajtich, R.; Barr, F.A. Plk1 docking to GRASP65 phosphorylated by Cdk1 suggests a mechanism for Golgi checkpoint signalling. EMBO J. 2005, 24, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Sohda, M.; Misumi, Y.; Yamamoto, A.; Yano, A.; Nakamura, N.; Ikehara, Y. Identification and characterization of a novel Golgi protein, GCP60, that interacts with the integral membrane protein giantin. J. Biol. Chem. 2001, 276, 45298–45306. [Google Scholar] [CrossRef] [PubMed]
- Sbodio, J.I.; Hicks, S.W.; Simon, D.; Machamer, C.E. GCP60 preferentially interacts with a caspase-generated golgin-160 fragment. J. Biol. Chem. 2006, 281, 27924–27931. [Google Scholar] [CrossRef]
- Sbodio, J.I.; Machamer, C.E. Identification of a redox-sensitive cysteine in GCP60 that regulates its interaction with golgin-160. J. Biol. Chem. 2007, 282, 29874–29881. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Guan, Y.; Chen, W.; Shi, C.; Yao, D.; Wang, F.; Lam, S.M.; Shui, G.; Cao, X. ACBD3 is required for FAPP2 transferring glucosylceramide through maintaining the Golgi integrity. J. Mol. Cell Biol. 2018. [Google Scholar] [CrossRef]
- Fan, J.; Liu, J.; Culty, M.; Papadopoulos, V. Acyl-coenzyme A binding domain containing 3 (ACBD3; PAP7; GCP60): An emerging signaling molecule. Prog. Lipid Res. 2010, 49, 218–234. [Google Scholar] [CrossRef] [PubMed]
- Soupene, E.; Kuypers, F.A. Ligand binding to the ACBD6 protein regulates the acyl-CoA transferase reactions in membranes. J. Lipid Res. 2015, 56, 1961–1971. [Google Scholar] [CrossRef]
- Ishikawa-Sasaki, K.; Sasaki, J.; Taniguchi, K. A complex comprising phosphatidylinositol 4-kinase IIIbeta, ACBD3, and Aichi virus proteins enhances phosphatidylinositol 4-phosphate synthesis and is critical for formation of the viral replication complex. J. Virol. 2014, 88, 6586–6598. [Google Scholar] [CrossRef]
- Yue, X.; Bao, M.; Christiano, R.; Li, S.; Mei, J.; Zhu, L.; Mao, F.; Yue, Q.; Zhang, P.; Jing, S.; et al. ACBD3 functions as a scaffold to organize the Golgi stacking proteins and a Rab33b-GAP. FEBS Lett. 2017, 591, 2793–2802. [Google Scholar] [CrossRef]
- Belman, J.P.; Bian, R.R.; Habtemichael, E.N.; Li, D.T.; Jurczak, M.J.; Alcazar-Roman, A.; McNally, L.J.; Shulman, G.I.; Bogan, J.S. Acetylation of TUG protein promotes the accumulation of GLUT4 glucose transporters in an insulin-responsive intracellular compartment. J. Biol. Chem. 2015, 290, 4447–4463. [Google Scholar] [CrossRef]
- Sbodio, J.I.; Paul, B.D.; Machamer, C.E.; Snyder, S.H. Golgi protein ACBD3 mediates neurotoxicity associated with Huntington’s disease. Cell Rep. 2013, 4, 890–897. [Google Scholar] [CrossRef]
- Greninger, A.L.; Knudsen, G.M.; Betegon, M.; Burlingame, A.L.; DeRisi, J.L. ACBD3 interaction with TBC1 domain 22 protein is differentially affected by enteroviral and kobuviral 3A protein binding. MBio 2013, 4, e00098-13. [Google Scholar] [CrossRef]
- Shinoda, Y.; Fujita, K.; Saito, S.; Matsui, H.; Kanto, Y.; Nagaura, Y.; Fukunaga, K.; Tamura, S.; Kobayashi, T. Acyl-CoA binding domain containing 3 (ACBD3) recruits the protein phosphatase PPM1L to ER-Golgi membrane contact sites. FEBS Lett. 2012, 586, 3024–3029. [Google Scholar] [CrossRef]
- Okazaki, Y.; Ma, Y.; Yeh, M.; Yin, H.; Li, Z.; Yeh, K.Y.; Glass, J. DMT1 (IRE) expression in intestinal and erythroid cells is regulated by peripheral benzodiazepine receptor-associated protein 7. Am. J. Physiol. 2012, 302, G1180–G1190. [Google Scholar] [CrossRef]
- Greninger, A.L.; Knudsen, G.M.; Betegon, M.; Burlingame, A.L.; Derisi, J.L. The 3A protein from multiple picornaviruses utilizes the golgi adaptor protein ACBD3 to recruit PI4KIIIbeta. J. Virol. 2012, 86, 3605–3616. [Google Scholar] [CrossRef]
- Zhou, Y.; Atkins, J.B.; Rompani, S.B.; Bancescu, D.L.; Petersen, P.H.; Tang, H.; Zou, K.; Stewart, S.B.; Zhong, W. The mammalian Golgi regulates numb signaling in asymmetric cell division by releasing ACBD3 during mitosis. Cell 2007, 129, 163–178. [Google Scholar] [CrossRef]
- Chen, Y.; Bang, S.; Park, S.; Shi, H.; Kim, S.F. Acyl-CoA-binding domain containing 3 modulates NAD+ metabolism through activating poly(ADP-ribose) polymerase 1. Biochem. J. 2015, 469, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Klima, M.; Toth, D.J.; Hexnerova, R.; Baumlova, A.; Chalupska, D.; Tykvart, J.; Rezabkova, L.; Sengupta, N.; Man, P.; Dubankova, A.; et al. Structural insights and in vitro reconstitution of membrane targeting and activation of human PI4KB by the ACBD3 protein. Sci. Rep. 2016, 6, 23641. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.J.; Liu, M.; Holden, D.W. Salmonella Effectors SseF and SseG Interact with Mammalian Protein ACBD3 (GCP60) To Anchor Salmonella-Containing Vacuoles at the Golgi Network. MBio 2016, 7, e00474-16. [Google Scholar] [CrossRef] [PubMed]
- Greninger, A.L. Picornavirus--host interactions to construct viral secretory membranes. Prog. Mol. Biol. Transl. Sci. 2015, 129, 189–212. [Google Scholar]
- Teoule, F.; Brisac, C.; Pelletier, I.; Vidalain, P.O.; Jegouic, S.; Mirabelli, C.; Bessaud, M.; Combelas, N.; Autret, A.; Tangy, F.; et al. The Golgi protein ACBD3, an interactor for poliovirus protein 3A, modulates poliovirus replication. J. Virol. 2013, 87, 11031–11046. [Google Scholar]
- Sasaki, J.; Ishikawa, K.; Arita, M.; Taniguchi, K. ACBD3-mediated recruitment of PI4KB to picornavirus RNA replication sites. Embo J. 2012, 31, 754–766. [Google Scholar] [CrossRef]
- Liu, J.; Matyakhina, L.; Han, Z.; Sandrini, F.; Bei, T.; Stratakis, C.A.; Papadopoulos, V. Molecular cloning, chromosomal localization of human peripheral-type benzodiazepine receptor and PKA regulatory subunit type 1A (PRKAR1A)-associated protein PAP7, and studies in PRKAR1A mutant cells and tissues. FASEB J. 2003, 17, 1189–1191. [Google Scholar] [CrossRef]
- Liu, J.; Rone, M.B.; Papadopoulos, V. Protein-protein interactions mediate mitochondrial cholesterol transport and steroid biosynthesis. J. Biol. Chem. 2006, 281, 38879–38893. [Google Scholar] [CrossRef]
- Chen, Y.; Patel, V.; Bang, S.; Cohen, N.; Millar, J.; Kim, S.F. Maturation and activity of sterol regulatory element binding protein 1 is inhibited by acyl-CoA binding domain containing 3. PLoS ONE 2012, 7, e49906. [Google Scholar] [CrossRef]
- Bai, P. Biology of Poly(ADP-Ribose) Polymerases: The Factotums of Cell Maintenance. Mol. Cell 2015, 58, 947–958. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- D’Angelo, G.; Polishchuk, E.; Di Tullio, G.; Santoro, M.; Di Campli, A.; Godi, A.; West, G.; Bielawski, J.; Chuang, C.C.; van der Spoel, A.C.; et al. Glycosphingolipid synthesis requires FAPP2 transfer of glucosylceramide. Nature 2007, 449, 62–67. [Google Scholar] [CrossRef]
- D’Angelo, G.; Uemura, T.; Chuang, C.C.; Polishchuk, E.; Santoro, M.; Ohvo-Rekila, H.; Sato, T.; Di Tullio, G.; Varriale, A.; D’Auria, S.; et al. Vesicular and non-vesicular transport feed distinct glycosylation pathways in the Golgi. Nature 2013, 501, 116–120. [Google Scholar] [CrossRef]
- Saito, S.; Matsui, H.; Kawano, M.; Kumagai, K.; Tomishige, N.; Hanada, K.; Echigo, S.; Tamura, S.; Kobayashi, T. Protein phosphatase 2Cepsilon is an endoplasmic reticulum integral membrane protein that dephosphorylates the ceramide transport protein CERT to enhance its association with organelle membranes. J. Biol. Chem. 2008, 283, 6584–6593. [Google Scholar] [CrossRef]
- Pusapati, G.V.; Luchetti, G.; Pfeffer, S.R. Ric1-Rgp1 complex is a guanine nucleotide exchange factor for the late Golgi Rab6A GTPase and an effector of the medial Golgi Rab33B GTPase. J. Biol. Chem. 2012, 287, 42129–42137. [Google Scholar] [CrossRef]
- Pfeffer, S.R. Rab GTPase localization and Rab cascades in Golgi transport. Biochem. Soc. Trans. 2012, 40, 1373–1377. [Google Scholar] [CrossRef]
- Chen, Y.; Mathias, L.; Falero-Perez, J.M.; Kim, S.F. PKA-mediated phosphorylation of Dexras1 suppresses iron trafficking by inhibiting S-nitrosylation. FEBS Lett. 2015, 589, 3212–3219. [Google Scholar] [CrossRef]
- Wilkinson, G.; Dennis, D.; Schuurmans, C. Proneural genes in neocortical development. Neuroscience 2013, 253, 256–273. [Google Scholar] [CrossRef]
- Delang, L.; Paeshuyse, J.; Neyts, J. The role of phosphatidylinositol 4-kinases and phosphatidylinositol 4-phosphate during viral replication. Biochem. Pharmacol. 2012, 84, 1400–1408. [Google Scholar] [CrossRef]
- Arita, M.; Wakita, T.; Shimizu, H. Valosin-containing protein (VCP/p97) is required for poliovirus replication and is involved in cellular protein secretion pathway in poliovirus infection. J. Virol. 2012, 86, 5541–5553. [Google Scholar] [CrossRef]
- Ishikawa-Sasaki, K.; Nagashima, S.; Taniguchi, K.; Sasaki, J. Model of OSBP-Mediated Cholesterol Supply to Aichi Virus RNA Replication Sites Involving Protein-Protein Interactions among Viral Proteins, ACBD3, OSBP, VAP-A/B, and SAC1. J. Virol. 2018, 92, e01952-17. [Google Scholar] [CrossRef]
- Xiao, X.; Lei, X.; Zhang, Z.; Ma, Y.; Qi, J.; Wu, C.; Xiao, Y.; Li, L.; He, B.; Wang, J. Enterovirus 3A facilitates viral replication by promoting PI4KB-ACBD3 interaction. J. Virol. 2017. [Google Scholar] [CrossRef]
- Roberts, M.F. Defining a Viral Membrane-Remodeling Complex on an Atomic Level. Structure 2017, 25, 3–4. [Google Scholar] [CrossRef]
- McPhail, J.A.; Ottosen, E.H.; Jenkins, M.L.; Burke, J.E. The Molecular Basis of Aichi Virus 3A Protein Activation of Phosphatidylinositol 4 Kinase IIIbeta, PI4KB, through ACBD3. Structure 2017, 25, 121–131. [Google Scholar] [CrossRef]
- Klima, M.; Chalupska, D.; Rozycki, B.; Humpolickova, J.; Rezabkova, L.; Silhan, J.; Baumlova, A.; Dubankova, A.; Boura, E. Kobuviral Non-structural 3A Proteins Act as Molecular Harnesses to Hijack the Host ACBD3 Protein. Structure 2017, 25, 219–230. [Google Scholar] [CrossRef]
- Dorobantu, C.M.; van der Schaar, H.M.; Ford, L.A.; Strating, J.R.; Ulferts, R.; Fang, Y.; Belov, G.; van Kuppeveld, F.J. Recruitment of PI4KIIIbeta to coxsackievirus B3 replication organelles is independent of ACBD3, GBF1, and Arf1. J. Virol. 2014, 88, 2725–2736. [Google Scholar] [CrossRef]
- MacDonald, M.E.; Ambrose, C.M.; Duyao, M.P.; Myers, R.H.; Lin, C.; Srinidhi, L.; Barnes, G.; Taylor, S.A.; James, M.; Groot, N.; et al. A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993, 72, 971–983. [Google Scholar] [CrossRef]
- Subramaniam, S.; Sixt, K.M.; Barrow, R.; Snyder, S.H. Rhes, a striatal specific protein, mediates mutant-huntingtin cytotoxicity. Science 2009, 324, 1327–1330. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, L.; Pei, Y.Y.; Wang, J.; Wu, H.; Yuan, J.; Wang, L. Overexpressed ACBD3 has prognostic value in human breast cancer and promotes the self-renewal potential of breast cancer cells by activating the Wnt/beta-catenin signaling pathway. Exp. Cell Res. 2018, 363, 39–47. [Google Scholar] [CrossRef]
- Lytle, N.K.; Barber, A.G.; Reya, T. Stem cell fate in cancer growth, progression and therapy resistance. Nat. Rev. Cancer 2018, 18, 669–680. [Google Scholar] [CrossRef]
- O’Brien, C.A.; Kreso, A.; Ryan, P.; Hermans, K.G.; Gibson, L.; Wang, Y.; Tsatsanis, A.; Gallinger, S.; Dick, J.E. ID1 and ID3 regulate the self-renewal capacity of human colon cancer-initiating cells through p21. Cancer Cell 2012, 21, 777–792. [Google Scholar] [CrossRef]
- Kakiuchi, S.; Daigo, Y.; Ishikawa, N.; Furukawa, C.; Tsunoda, T.; Yano, S.; Nakagawa, K.; Tsuruo, T.; Kohno, N.; Fukuoka, M.; et al. Prediction of sensitivity of advanced non-small cell lung cancers to gefitinib (Iressa, ZD1839). Hum. Mol. Genet. 2004, 13, 3029–3043. [Google Scholar] [CrossRef]
- Knoblich, J.A. Asymmetric cell division: Recent developments and their implications for tumour biology. Nat. Rev. Mol. Cell Biol. 2010, 11, 849–860. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Protein Names | Molecular Weight | Intracellular Localization | Functions | References |
|---|---|---|---|---|
| Giantin | 372 kDa | Golgi | Golgi organization; ER to Golgi transport | [6] |
| Golgin160 | 160 kDa | Golgi | Maintenance of Golgi structure | [7,8] |
| Golgin45 | 45 kDa | Golgi and nucleus | Required for normal Golgi structure and for protein transport from the ER through the Golgi to the cell surface | [11] |
| Numb | 72 kDa | plasma membrane, nucleus and cytosol | Plays a role in the determination of cell fates during development | [19] |
| PI4KB | 92 kDa | Golgi, endosome and cytosol | May regulate Golgi disintegration/reorganization during mitosis; Involved in Golgi-to-plasma membrane trafficking | [10,17,21,42,44,45] |
| TUG | 60 kDa | plasma membrane, nucleus and cytosol | Tethering protein that sequesters GLUT4-containing vesicles in the cytoplasm in the absence of insulin. Modulates the amount of GLUT4 that is available at the cell surface. Enhances VCP methylation catalyzed by VCPKMT. | [12] |
| Htt | 35 kDa | cytoskeleton, nucleus, cytosol, ER, Golgi and endosome | Huntingtin is a disease gene linked to Huntington’s disease; May play a role in microtubule-mediated transport or vesicle function | [13] |
| PBR | 18 kDa | mitochondria | Promotes the transport of cholesterol across mitochondrial membranes and may play a role in lipid metabolism | [26] |
| PRKAR1A | 43 kDa | plasma membrane and cytosol | Regulatory subunit of the cAMP-dependent protein kinases involved in cAMP signaling in cells | [26] |
| 3A protein | Nonstructural viral protein required for RNA replication | [14,23,24,42,44,45] | ||
| SseF and SseG | Both are (Salmonella pathogenicity island 2) SPI-2-encoded effectors which are necessary for the retention of tightly clustered bacterial microcolonies in close proximity to the MTOC and the Golgi | [22] | ||
| DMT1 | 62 kDa | plasma membrane, mitochondria, nucleus lysosome, endosome and Golgi | Important in metal transport, in particular iron | [16] |
| PARP1 | 110 kDa | Mitochondria and nucleus | Involved in the base excision repair (BER) pathway, by catalyzing the poly(ADP-ribosyl)ation of a limited number of acceptor proteins involved in chromatin architecture and in DNA metabolism | [20] |
| SREBP1 | 120 kDa | ER, nucleus, cytosol and Golgi | Transcriptional activator required for lipid homeostasis | [28] |
| FAPP2 | 58 kDa | nucleus and Golgi | Cargo transport protein that is required for apical transport from the Golgi complex; Mediates the non-vesicular transport of glucosylceramide (GlcCer) from the trans-Golgi network (TGN) to the plasma membrane and plays a pivotal role in the synthesis of complex glycosphingolipids | [9] |
| TBC1D22A | 47 kDa | Perinuclear, Golgi | GTPase activating protein (GAP) for Rab33b; membrane trafficking | [14] |
| PPM1L | 41 kDa | ER | Helps regulate ceramide transport from the ER to the Golgi; Acts as a suppressor of the SAPK signaling pathways | [15] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, X.; Qian, Y.; Gim, B.; Lee, I. Acyl-CoA-Binding Domain-Containing 3 (ACBD3; PAP7; GCP60): A Multi-Functional Membrane Domain Organizer. Int. J. Mol. Sci. 2019, 20, 2028. https://doi.org/10.3390/ijms20082028
Yue X, Qian Y, Gim B, Lee I. Acyl-CoA-Binding Domain-Containing 3 (ACBD3; PAP7; GCP60): A Multi-Functional Membrane Domain Organizer. International Journal of Molecular Sciences. 2019; 20(8):2028. https://doi.org/10.3390/ijms20082028
Chicago/Turabian StyleYue, Xihua, Yi Qian, Bopil Gim, and Intaek Lee. 2019. "Acyl-CoA-Binding Domain-Containing 3 (ACBD3; PAP7; GCP60): A Multi-Functional Membrane Domain Organizer" International Journal of Molecular Sciences 20, no. 8: 2028. https://doi.org/10.3390/ijms20082028
APA StyleYue, X., Qian, Y., Gim, B., & Lee, I. (2019). Acyl-CoA-Binding Domain-Containing 3 (ACBD3; PAP7; GCP60): A Multi-Functional Membrane Domain Organizer. International Journal of Molecular Sciences, 20(8), 2028. https://doi.org/10.3390/ijms20082028