Expression of Retinoid Acid Receptor-Responsive Genes in Rodent Models of Placental Pathology

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Auxology
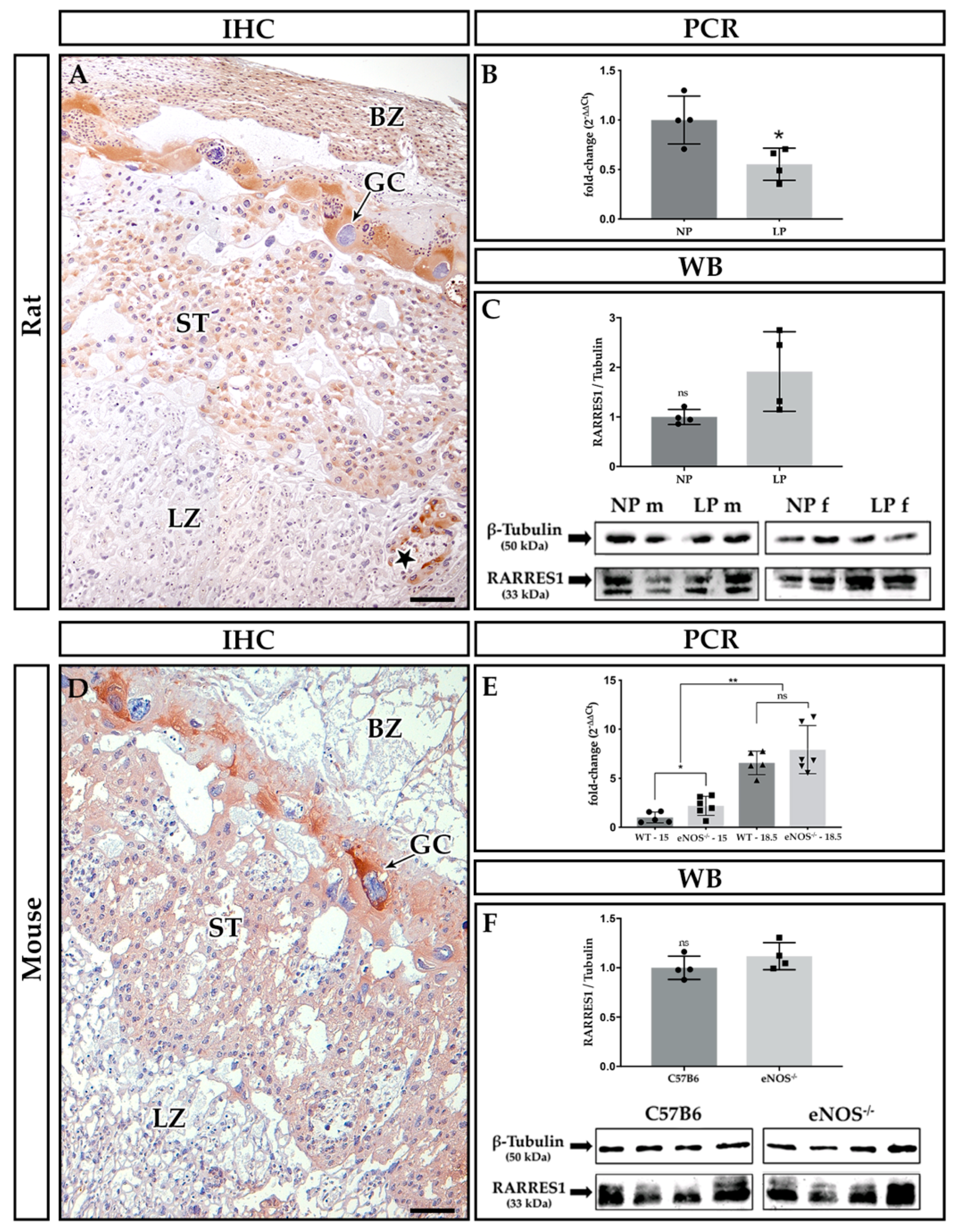
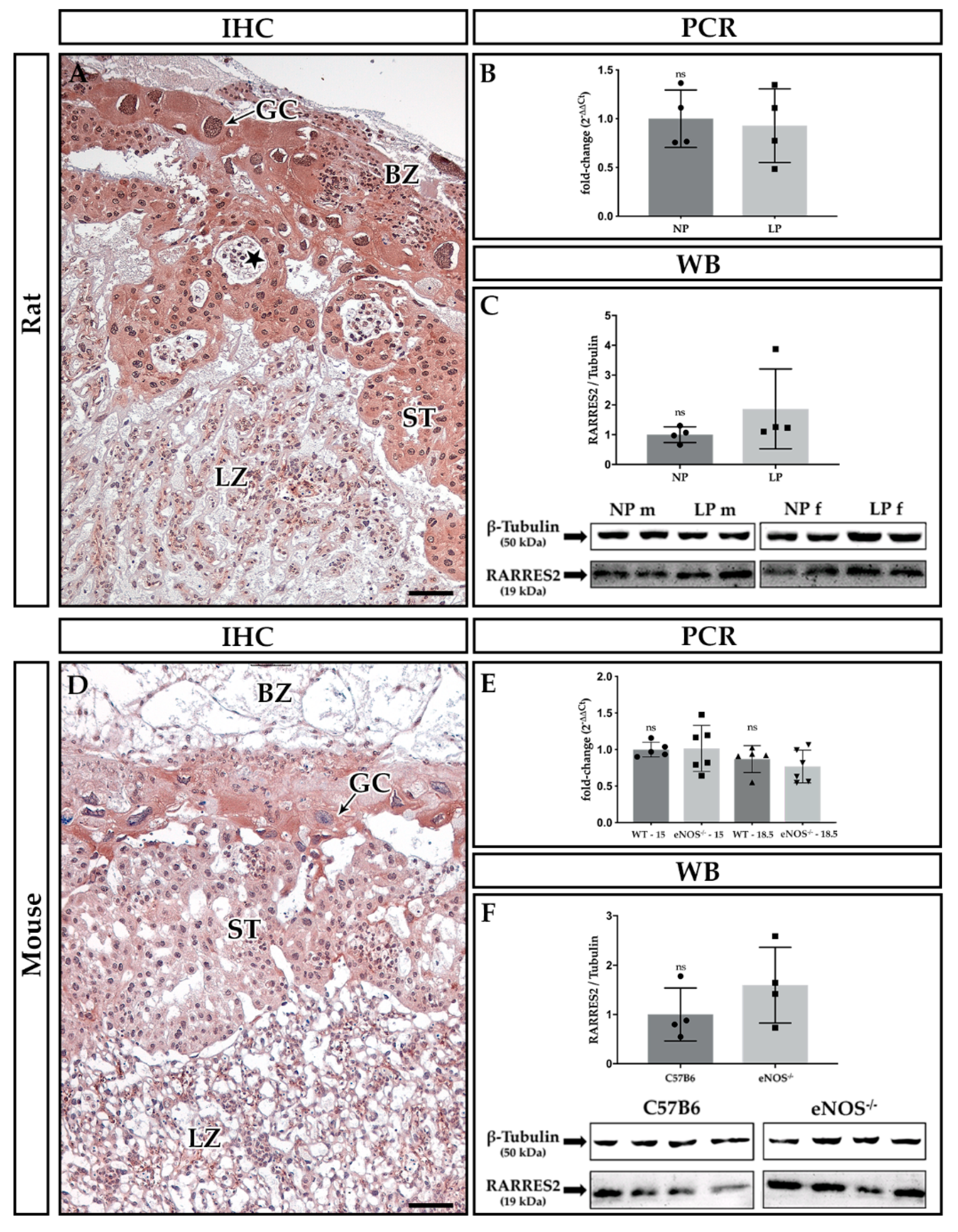
2.2. Localization of Rarres1 and 2
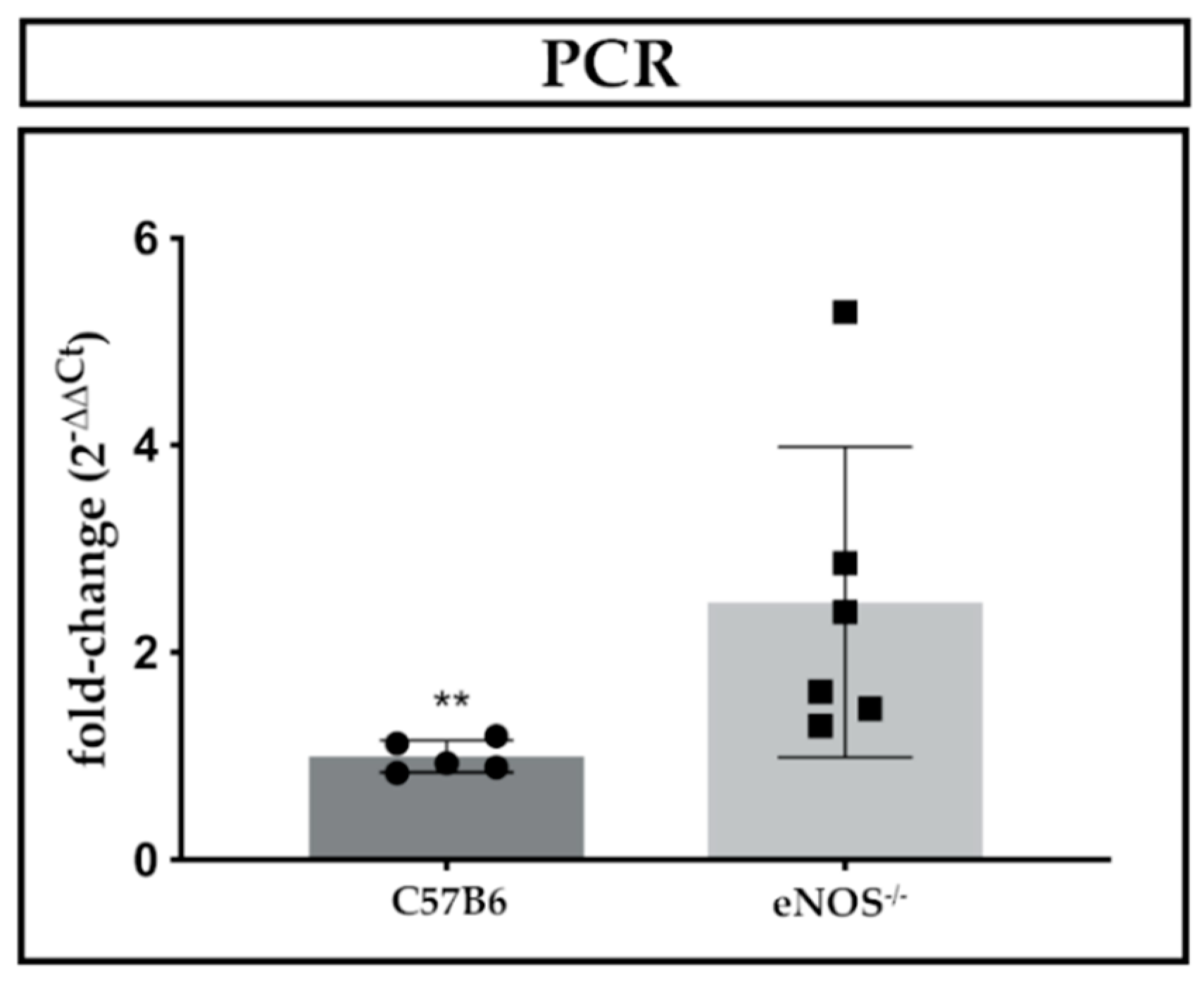
2.3. Expression Analyses of Rarres1/2, CmklR1 Receptor, and IL-11
3. Discussion
3.1. Expression of Rarres1 in Rodent Placenta
3.2. Expression of Rarres2 in Rodent Placenta
3.3. Expression of CmklR1 in Rodent Placenta
3.4. IL-11 as a Novel Regulatory Cytokine in eNOS−/− Mice
4. Materials and Methods
4.1. Animals and Diets
4.1.1. Alimentary Rat Model with IUGR-Like Features
4.1.2. Mouse Model with PE/IUGR-Like Features
4.2. RNA Extraction, RT-PCR, and Real-Time Quantitative PCR
4.3. Western Blot Analysis
4.4. Immunohistochemistry
4.5. Statistical Analysis
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huebner, H.; Hartner, A.; Rascher, W.; Strick, R.R.; Kehl, S.; Heindl, F.; Wachter, D.L.; Beckmann Md, M.W.; Fahlbusch, F.B.; Ruebner, M. Expression and Regulation of Retinoic Acid Receptor Responders in the Human Placenta. Reprod. Sci. 2018, 25, 1357–1370. [Google Scholar] [CrossRef] [PubMed]
- Huebner, H.; Strick, R.; Wachter, D.L.; Kehl, S.; Strissel, P.L.; Schneider-Stock, R.; Hartner, A.; Rascher, W.; Horn, L.C.; Beckmann, M.W.; et al. Hypermethylation and loss of retinoic acid receptor responder 1 expression in human choriocarcinoma. J. Exp. Clin. Cancer Res. 2017, 36, 165. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, T.; Ding, Y.; Zhong, Y.; Yu, L.; Peng, M. Chemerin Plays a Protective Role by Regulating Human Umbilical Vein Endothelial Cell-Induced Nitric Oxide Signaling in Preeclampsia. Endocrine 2015, 48, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, S.; Patel, S.; Asano, A.T.; Johnson, A.T.; Duvic, M.; Chandraratna, R.A. Tazarotene-Induced Gene 1 (TIG1), a Novel Retinoic Acid Receptor-Responsive Gene in Skin. J. Investig. Dermatol. 1996, 106, 269–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagpal, S.; Patel, S.; Jacobe, H.; Disepio, D.; Ghosn, C.; Malhotra, M.; Teng, M.; Duvic, M.; Chandraratna, R.A. Tazarotene-induced gene 2 (TIG2), a novel retinoid-responsive gene in skin. J. Investig. Dermatol. 1997, 109, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Garces, M.; Sánchez, E.; Acosta, B.; Angel, E.; Ruiz, A.; Rubio-Romero, J.; Diéguez, C.; Nogueiras, R.; Caminos, J. Expression and regulation of chemerin during rat pregnancy. Placenta 2012, 33, 373–378. [Google Scholar] [CrossRef]
- Roy, A.; Ramalinga, M.; Kim, O.J.; Chijioke, J.; Lynch, S.; Byers, S.; Kumar, D. Multiple roles of RARRES1 in prostate cancer: Autophagy induction and angiogenesis inhibition. PLoS ONE 2017, 12, 0180344. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Saso, H.; Iwamoto, T.; Xia, W.; Gong, Y.; Pusztai, L.; Woodward, W.A.; Reuben, J.M.; Warner, S.L.; Bearss, D.J.; et al. TIG1 Promotes the Development and Progression of Inflammatory Breast Cancer through Activation of Axl Kinase. Cancer Res. 2013, 73, 6516–6525. [Google Scholar] [CrossRef] [Green Version]
- Liu-Chittenden, Y.; Jain, M.; Gaskins, K.; Wang, S.; Merino, M.J.; Kotian, S.; Kumar Gara, S.; Davis, S.; Zhang, L.; Kebebew, E. Rarres2 Functions as a Tumor Suppressor by Promoting Beta-Catenin Phosphorylation/Degradation and Inhibiting P38 Phosphorylation in Adrenocortical Carcinoma. Oncogene 2017, 36, 3541–3552. [Google Scholar] [CrossRef] [Green Version]
- Oldridge, E.E.; Walker, H.F.; Stower, M.J.; Simms, M.S.; Mann, V.M.; Collins, A.T.; Pellacani, D.; Maitland, N.J. Retinoic acid represses invasion and stem cell phenotype by induction of the metastasis suppressors RARRES1 and LXN. Oncogenesis 2013, 2, e45. [Google Scholar] [CrossRef] [Green Version]
- Sahab, Z.J.; Hall, M.D.; Me Sung, Y.; Dakshanamurthy, S.; Ji, Y.; Kumar, D.; Byers, S.W. Tumor Suppressor Rarres1 Interacts with Cytoplasmic Carboxypeptidase Agbl2 to Regulate the Alpha-Tubulin Tyrosination Cycle. Cancer Res. 2011, 71, 1219–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meder, W.; Wendland, M.; Busmann, A.; Kutzleb, C.; Spodsberg, N.; John, H.; Richter, R.; Schleuder, D.; Meyer, M.; Forssmann, W. Characterization of human circulating TIG2 as a ligand for the orphan receptor ChemR23. FEBS Lett. 2003, 555, 495–499. [Google Scholar] [CrossRef] [Green Version]
- Zabel, B.A.; Silverio, A.M.; Butcher, E.C. Chemokine-like receptor 1 expression and chemerin-directed chemotaxis distinguish plasmacytoid from myeloid dendritic cells in human blood. J. Immunol. 2005, 174, 244–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehrke, M.; Becker, A.; Greif, M.; Stark, R.; Laubender, R.P.; Von Ziegler, F.; Lebherz, C.; Tittus, J.; Reiser, M.; Becker, C.; et al. Chemerin is associated with markers of inflammation and components of the metabolic syndrome but does not predict coronary atherosclerosis. Eur. J. Endocrinol. 2009, 161, 339–344. [Google Scholar] [CrossRef]
- Wittamer, V.; Franssen, J.D.; Vulcano, M.; Mirjolet, J.F.; Le Poul, E.; Migeotte, I.; Brezillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific Recruitment of Antigen-Presenting Cells by Chemerin, a Novel Processed Ligand from Human Inflammatory Fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef]
- Bondue, B.; Wittamer, V.; Parmentier, M. Chemerin and its receptors in leukocyte trafficking, inflammation and metabolism. Cytokine Growth Factor Rev. 2011, 22, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Redman, C.; Sargent, I. Pre-eclampsia, the Placenta and the Maternal Systemic Inflammatory Response—A Review. Placenta 2003, 24 (Suppl. A), S21–S27. [Google Scholar] [CrossRef]
- Podjarny, E.; Losonczy, G.; Baylis, C. Animal Models of Preeclampsia. Semin. Nephrol. 2004, 24, 596–606. [Google Scholar] [CrossRef]
- Sladek, S.M.; Magness, R.R.; Conrad, K.P. Nitric Oxide and Pregnancy. Am. J. Physiol. 1997, 272, R441–R463. [Google Scholar] [CrossRef] [Green Version]
- Depoortere, I.; Thijs, T.; Keith, J.; Peeters, T.L. Treatment with interleukin-11 affects plasma leptin levels in inflamed and non-inflamed rabbits. Regul. Pept. 2004, 122, 149–156. [Google Scholar] [CrossRef]
- Makrilakis, K.; Fragiadaki, K.; Smith, J.; Sfikakis, P.P.; Kitas, G.D. Interrelated Reduction of Chemerin and Plasminogen Activator Inhibitor-1 Serum Levels in Rheumatoid Arthritis after Interleukin-6 Receptor Blockade. Clin. Rheumatol. 2015, 34, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Fatima, S.S.; Alam, F.; Chaudhry, B.; Khan, T.A. Elevated Levels of Chemerin, Leptin, and Interleukin-18 in Gestational Diabetes Mellitus. J. Matern. Fetal Neonatal Med. 2017, 30, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, I.; Ohsumi, J.; Mita-Honjo, K.; Shimoda-Takano, K.; Ishikawa, H.; Sakakibara, S.; Miyadai, K.; Takiguchi, Y. Molecular cloning of cDNA encoding adipogenesis inhibitory factor and identity with interleukin-11. FEBS Lett. 1991, 283, 199–202. [Google Scholar] [CrossRef] [Green Version]
- Winship, A.; Dimitriadis, E. Interleukin 11 is upregulated in preeclampsia and leads to inflammation and preeclampsia features in mice. J. Reprod. Immunol. 2018, 125, 32–38. [Google Scholar] [CrossRef]
- Winship, A.L.; Koga, K.; Menkhorst, E.; Van Sinderen, M.; Rainczuk, K.; Nagai, M.; Cuman, C.; Yap, J.; Zhang, J.-G.; Simmons, D.; et al. Interleukin-11 alters placentation and causes preeclampsia features in mice. Proc. Natl. Acad. Sci.USA 2015, 112, 15928–15933. [Google Scholar] [CrossRef] [Green Version]
- Kusinski, L.C.; Stanley, J.L.; Dilworth, M.R.; Hirt, C.J.; Andersson, I.J.; Renshall, L.J.; Baker, B.C.; Baker, P.N.; Sibley, C.P.; Wareing, M.; et al. eNOS knockout mouse as a model of fetal growth restriction with an impaired uterine artery function and placental transport phenotype. Am. J. Physiol. Integr. Comp. Physiol. 2012, 303, R86–R93. [Google Scholar] [CrossRef]
- Anumba, D.O.C.; Robson, S.C.; Boys, R.J.; Ford, G.A. Nitric oxide activity in the peripheral vasculature during normotensive and preeclamptic pregnancy. Am. J. Physiol. Content 1999, 277, H848–H854. [Google Scholar] [CrossRef]
- Hefler, L.A.; Reyes, C.A.; O’Brien, W.E.; Gregg, A.R. Perinatal development of endothelial nitric oxide synthase-deficient mice. Boil. Reprod. 2001, 64, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Hefler, L.A.; Tempfer, C.B.; Moreno, R.M.; O’Brien, W.E.; Gregg, A.R. Endothelial-derived nitric oxide and angiotensinogen: Blood pressure and metabolism during mouse pregnancy. Am. J. Physiol. Integr. Comp. Physiol. 2001, 280, R174–R182. [Google Scholar] [CrossRef]
- Veerareddy, S.; Cooke, C.-L.M.; Baker, P.N.; Davidge, S.T. Vascular adaptations to pregnancy in mice: Effects on myogenic tone. Am. J. Physiol. Circ. Physiol. 2002, 283, H2226–H2233. [Google Scholar] [CrossRef] [Green Version]
- Flaherty, M.P.; Brown, M.; Grupp, I.L.; El Schultz, J.; Murphree, S.S.; Jones, W.K. eNOS Deficient Mice Develop Progressive Cardiac Hypertrophy with Altered Cytokine and Calcium Handling Protein Expression. Cardiovasc. Toxicol. 2007, 7, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Bucci, M.; Gratton, J.-P.; Rudic, R.D.; Acevedo, L.; Roviezzo, F.; Cirino, G.; Sessa, W.C. In vivo delivery of the caveolin-1 scaffolding domain inhibits nitric oxide synthesis and reduces inflammation. Nat. Med. 2000, 6, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- Bucci, M.; Roviezzo, F.; Posadas, I.; Yu, J.; Parente, L.; Sessa, W.C.; Ignarro, L.J.; Cirino, G. Endothelial nitric oxide synthase activation is critical for vascular leakage during acute inflammation in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 904–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Zhang, M.; Liang, X.; Jia, X.; Jia, J.; Zhao, M.; Fan, Y. Interleukin-33 Promotes Inflammation-Induced Lymphangiogenesis via St2/Traf6-Mediated Akt/Enos/No Signalling Pathway. Sci. Rep. 2017, 7, 10602. [Google Scholar] [CrossRef] [Green Version]
- Beinder, L.; Faehrmann, N.; Wachtveitl, R.; Winterfeld, I.; Hartner, A.; Menendez-Castro, C.; Rauh, M.; Ruebner, M.; Huebner, H.; Noegel, S.C.; et al. Detection of Expressional Changes Induced by Intrauterine Growth Restriction in the Developing Rat Mammary Gland via Exploratory Pathways Analysis. PLoS ONE 2014, 9, e100504. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Rauh, M.; Schmid, M.C.; Huebner, H.; Ruebner, M.; Wachtveitl, R.; Cordasic, N.; Rascher, W.; Menendez-Castro, C.; Hartner, A.; et al. Influence of Low Protein Diet-Induced Fetal Growth Restriction on the Neuroplacental Corticosterone Axis in the Rat. Front. Endocrinol. 2019, 10, 124. [Google Scholar] [CrossRef] [Green Version]
- Watts, S.W.; Darios, E.S.; Mullick, A.E.; Garver, H.; Saunders, T.L.; Hughes, E.D.; Filipiak, W.E.; Zeidler, M.G.; McMullen, N.; Sinal, C.J.; et al. The Chemerin Knockout Rat Reveals Chemerin Dependence in Female, but Not Male, Experimental Hypertension. FASEB J. 2018, fj201800479. [Google Scholar] [CrossRef]
- Langbein, M.; Strick, R.; Strissel, P.L.; Vogt, N.; Parsch, H.; Beckmann, M.W.; Schild, R.L. Impaired Cytotrophoblast Cell-Cell Fusion Is Associated with Reduced Syncytin and Increased Apoptosis in Patients with Placental Dysfunction. Mol. Reprod. Dev. 2008, 75, 175–183. [Google Scholar] [CrossRef]
- Burton, G.J.; Jones, C.J. Syncytial Knots, Sprouts, Apoptosis, and Trophoblast Deportation from the Human Placenta. Taiwan. J. Obstet. Gynecol. 2009, 48, 28–37. [Google Scholar] [CrossRef]
- Dilworth, M.; Sibley, C. Review: Transport across the placenta of mice and women. Placenta 2013, 34, S34–S39. [Google Scholar] [CrossRef]
- Silva, J.F.; Serakides, R. Intrauterine trophoblast migration: A comparative view of humans and rodents. Cell Adhes. Migr. 2016, 10, 88–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemberger, M.; Cross, J.C. Genes governing placental development. Trends Endocrinol. Metab. 2001, 12, 162–168. [Google Scholar] [CrossRef]
- Cross, J.; Baczyk, D.; Dobric, N.; Hemberger, M.; Hughes, M.; Simmons, D.; Yamamoto, H.; Kingdom, J. Genes, Development and Evolution of the Placenta. Placenta 2003, 24, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Georgiades, P.; Ferguson-Smith, A.; Burton, G. Comparative Developmental Anatomy of the Murine and Human Definitive Placentae. Placenta 2002, 23, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.G.; Cross, J.C. Determinants of trophoblast lineage and cell subtype specification in the mouse placenta. Dev. Boil. 2005, 284, 12–24. [Google Scholar] [CrossRef] [Green Version]
- Woods, L.; Perez-Garcia, V.; Hemberger, M. Regulation of Placental Development and Its Impact on Fetal Growth—New Insights from Mouse Models. Front. Endocrinol. 2018, 9, 570. [Google Scholar] [CrossRef] [Green Version]
- Abreu, R.D.S.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global signatures of protein and mRNA expression levels. Mol. BioSyst. 2009, 5, 1512–1526. [Google Scholar]
- Ruebner, M.; Strissel, P.L.; Langbein, M.; Fahlbusch, F.; Wachter, D.L.; Faschingbauer, F.; Beckmann, M.W.; Strick, R. Impaired cell fusion and differentiation in placentae from patients with intrauterine growth restriction correlate with reduced levels of HERV envelope genes. J. Mol. Med. 2010, 88, 1143–1156. [Google Scholar] [CrossRef]
- Ruebner, M.; Strissel, P.L.; Ekici, A.B.; Stiegler, E.; Dammer, U.; Goecke, T.W.; Faschingbauer, F.; Fahlbusch, F.B.; Beckmann, M.W.; Strick, R. Reduced Syncytin-1 Expression Levels in Placental Syndromes Correlates with Epigenetic Hypermethylation of the ERVW-1 Promoter Region. PLoS ONE 2013, 8, 56145. [Google Scholar] [CrossRef] [Green Version]
- Garces, M.F.; Sanchez, E.; Ruíz-Parra, A.I.; Rubio-Romero, J.A.; Angel-Müller, E.; Suarez, M.A.; Bohórquez, L.F.; Bravo, S.B.; Nogueiras, R.; Diéguez, C.; et al. Serum chemerin levels during normal human pregnancy. Peptides 2013, 42, 138–143. [Google Scholar] [CrossRef]
- Snoeck, A.; Hoet, J.J.; Remacle, C.; Reusens, B. Effect of a Low Protein Diet during Pregnancy on the Fetal Rat Endocrine Pancreas. Biol. Neonate 1990, 57, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Blondeau, B.; Lesage, J.; Czernichow, P.; Dupouy, J.P.; Bréant, B. Glucocorticoids impair fetal beta-cell development in rats. Am. J. Physiol. Metab. 2001, 281, 592–599. [Google Scholar]
- Mühlhäusler, B.S. Programming of the Appetite-Regulating Neural Network: A Link Between Maternal Overnutrition and the Programming of Obesity? J. Neuroendocr. 2007, 19, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Valsamakis, G.; Kumar, S.; Creatsas, G.; Mastorakos, G. The effects of adipose tissue and adipocytokines in human pregnancy. Ann. N. Y. Acad. Sci. 2010, 1205, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Ritterath, C.; Rad, N.T.; Siegmund, T.; Heinze, T.; Siebert, G.; Buhling, K.J. Adiponectin During Pregnancy: Correlation with Fat Metabolism, but Not with Carbohydrate Metabolism. Arch. Gynecol. Obstet. 2010, 281, 91–96. [Google Scholar] [CrossRef]
- Briana, D.D.; Malamitsi-Puchner, A. The role of adipocytokines in fetal growth. Ann. N. Y. Acad. Sci. 2010, 1205, 82–87. [Google Scholar] [CrossRef]
- Barker, G.; Lim, R.; Rice, G.E.; Lappas, M. Increased chemerin concentrations in fetuses of obese mothers and correlation with maternal insulin sensitivity. J. Matern. Neonatal Med. 2012, 25, 2274–2280. [Google Scholar] [CrossRef]
- Sanchez-Rebordelo, E.; Cunarro, J.; Perez-Sieira, S.; Seoane, L.M.; Diéguez, C.; Nogueiras, R.; Tovar, S. Regulation of Chemerin and CMKLR1 Expression by Nutritional Status, Postnatal Development, and Gender. Int. J. Mol. Sci. 2018, 19, 2905. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.J.; Yang, P.; Read, C.; Kuc, R.E.; Yang, L.; Taylor, E.J.; Taylor, C.W.; Maguire, J.J.; Davenport, A.P. Chemerin Elicits Potent Constrictor Actions Via Chemokine-Like Receptor 1 (Cmklr1), Not G-Protein-Coupled Receptor 1 (Gpr1), in Human and Rat Vasculature. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Kulandavelu, S.; Whiteley, K.J.; Qu, D.; Mu, J.; Bainbridge, S.A.; Adamson, S.L. Endothelial Nitric Oxide Synthase Deficiency Reduces Uterine Blood Flow, Spiral Artery Elongation, and Placental Oxygenation in Pregnant Mice. Hypertension 2012, 60, 231–238. [Google Scholar] [CrossRef]
- Yang, X.; Yao, J.; Wei, Q.; Ye, J.; Yin, X.; Quan, X.; Lan, Y.; Xing, H. Role of chemerin/CMKLR1 in the maintenance of early pregnancy. Front. Med. 2018, 12, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Castro, C.; Fahlbusch, F.; Cordasic, N.; Amann, K.; Münzel, K.; Plank, C.; Wachtveitl, R.; Rascher, W.; Hilgers, K.F.; Hartner, A. Early and Late Postnatal Myocardial and Vascular Changes in a Protein Restriction Rat Model of Intrauterine Growth Restriction. PLoS ONE 2011, 6, e20369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayward, C.E.; Lean, S.; Sibley, C.P.; Jones, R.L.; Wareing, M.; Greenwood, S.L.; Dilworth, M.R. Placental Adaptation: What Can We Learn from Birthweight: Placental Weight Ratio? Front. Physiol. 2016, 7, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Heijden, O.W.; Essers, Y.P.; Fazzi, G.; Peeters, L.L.; De Mey, J.G.R.; Van Eys, G.J. Uterine Artery Remodeling and Reproductive Performance Are Impaired in Endothelial Nitric Oxide Synthase-Deficient Mice1. Boil. Reprod. 2005, 72, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Pallares, P.; Gonzalez-Bulnes, A. The effect of embryo and maternal genotypes on prolificacy, intrauterine growth retardation and postnatal development of Nos3-knockout mice. Reprod. Boil. 2010, 10, 241–248. [Google Scholar] [CrossRef]
- Herdl, S.; Huebner, H.; Volkert, G.; Marek, I.; Menendez-Castro, C.; Noegel, S.C.; Ruebner, M.; Rascher, W.; Hartner, A.; Fahlbusch, F.B. Integrin Alpha8 Is Abundant in Human, Rat, and Mouse Trophoblasts. Reprod. Sci. 2017, 24, 1426–1437. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Castro, C.; Nitz, D.; Cordasic, N.; Jordan, J.; Bäuerle, T.; Fahlbusch, F.B.; Rascher, W.; Hilgers, K.F.; Hartner, A. Neonatal nephron loss during active nephrogenesis—Detrimental impact with long-term renal consequences. Sci. Rep. 2018, 8, 4542. [Google Scholar] [CrossRef] [Green Version]
- Motulsky, H.J.; E Brown, R. Detecting outliers when fitting data with nonlinear regression—A new method based on robust nonlinear regression and the false discovery rate. BMC Bioinform. 2006, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Hughes, B.G.; Hekimi, S. Different Mechanisms of Longevity in Long-Lived Mouse andCaenorhabditis elegansMutants Revealed by Statistical Analysis of Mortality Rates. Genetics 2016, 204, 905–920. [Google Scholar] [CrossRef] [Green Version]
- Pallares, P.; Gonzalez-Bulnes, A. Intrauterine Growth Retardation in Endothelial Nitric Oxide Synthase-Deficient Mice Is Established from Early Stages of Pregnancy. Boil. Reprod. 2008, 78, 1002–1006. [Google Scholar] [CrossRef] [Green Version]
- Shesely, E.G.; Gilbert, C.; Granderson, G.; Carretero, C.; Carretero, O.A.; Beierwaltes, W.H. Nitric oxide synthase gene knockout mice do not become hypertensive during pregnancy. Am. J. Obstet. Gynecol. 2001, 185, 1198–1203. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Rat † | E18.5 NP | E18.5 LP | p Value | |||
| fetal weight (fw) | 1.38 ± 0.09 | 0.86 ± 0.05 | 0.03 * | |||
| placental weight (pw) | 0.34 ± 0.04 | 0.30 ± 0.01 | 0.11 * | |||
| pw/fw ratio | 0.25 ± 0.02 | 0.35 ± 0.02 | 0.03 * | |||
| Mouse ‡ | E15 C57BL/6 | E15 eNOS−/− | p Value | E18.5 C57BL/6 | E18.5 eNOS−/− | p Value |
| fetal weight (fw) | 0.34 ± 0.07 | 0.28 ± 0.03 | 0.08 * | 1.19 ± 0.15 | 0.97 ± 0.08 | 0.006 * |
| placental weight (pw) | 0.10 ± 0.01 | 0.08 ± 0.01 | <0.001 * | 0.09 ± 0.01 | 0.07 ± 0.01 | <0.001 * |
| pw/fw ratio | 0.31 ± 0.06 | 0.29 ± 0.06 | 0.58 * | 0.08 ± 0.02 | 0.07 ± 0.01 | 0.63 * |
| Rat | NP m | NP f | p Value | LP m | LP f | p Value |
|---|---|---|---|---|---|---|
| Rarres1 | 1.00 ± 0.36 | 0.97 ± 0.17 | 0.89 * | 0.48 ± 0.26 | 0.58 ± 0.19 | 0.69 * |
| Rarres2 | 1.00 ± 0.30 | 1.30 ± 0.41 | 0.49 * | 0.90 ± 0.75 | 1.52 ± 0.42 | 0.23 * |
| CmklR1 | 1.00 ± 0.44 | 0.89 ± 0.60 | 0.99 * | 1.10 ± 0.51 | 0.95 ± 0.29 | 0.99 * |
| Rat † | E18.5 NP | E18.5 LP | p Value | |||
| 1.00 ± 0.53 | 1.08 ± 0.32 | 0.99 * | ||||
| Mouse ‡ | E15 C57BL/6 | E15 eNOS−/− | p Value | E18.5 C57BL/6 | E18.5 eNOS−/− | p Value |
| 1.00 ± 0.43 | 0.9 ± 0.42 | 0.93 * | 0.51 ± 0.1 | 0.41 ± 0.18 | 0.18 * |
| Rat | Forward | Reverse |
| Rarres1 | 5′-AGGTGGACCTGGTGTTTAGCA-3′ | 5′-AACACCCTCGCAGAACATTTG-3’ |
| Rarres2 | 5′-AAATGGGAGGAAGCGGAAAT-3′ | 5′-CCATCCGGCCTAGAACTTTACC-3′ |
| CmlkR1 | 5′-AAGAGATGGAGTACGAGGGTTACAA-3′ | 5′-GATGTAGTCCGAGCCGTCAGA-3′ |
| r18s | 5′-TTGATTAAGTCCCTGCCCTTTGT-3′ | 5′-CGATCCGAGGGCCTCACTA-3′ |
| Mouse | ||
| Rarres1 | 5′-AGCGGCTGAAAACGGATGA-3′ | 5′-CCAAGTGAATACGGCAGGGA-3′ |
| Rarres2 | 5′-CACTGCCCAATTCTGAAGCAA-3′ | 5′-CGCCAGCCTGTGCTATCTTAA-3′ |
| Cmlkr1 | 5′- CAACGGTGAACAGTGAAAGGTC-3‘ | 5′- TTGTAAGCGTCGTACTCCATCTCT-3‘ |
| eNos | 5′-CACCAGGAAGAAGACCTTTAAGGA-3′ | 5′-CACCGTGCCCATGAGTGA-3′ |
| IL-11 | 5′-GCTCCCCTCGAGTCTCTTCA-3′ | 5′-TGTCTCTCATCTGTGCAGCTAGTTG-3′ |
| r18s | 5′-TTGATTAAGTCCCTGCCCTTTGT-3′ | 5′-CGATCCGAGGGCCTCACTA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mocker, A.; Schmidt, M.; Huebner, H.; Wachtveitl, R.; Cordasic, N.; Menendez-Castro, C.; Hartner, A.; Fahlbusch, F.B. Expression of Retinoid Acid Receptor-Responsive Genes in Rodent Models of Placental Pathology. Int. J. Mol. Sci. 2020, 21, 242. https://doi.org/10.3390/ijms21010242
Mocker A, Schmidt M, Huebner H, Wachtveitl R, Cordasic N, Menendez-Castro C, Hartner A, Fahlbusch FB. Expression of Retinoid Acid Receptor-Responsive Genes in Rodent Models of Placental Pathology. International Journal of Molecular Sciences. 2020; 21(1):242. https://doi.org/10.3390/ijms21010242
Chicago/Turabian StyleMocker, Alexander, Marius Schmidt, Hanna Huebner, Rainer Wachtveitl, Nada Cordasic, Carlos Menendez-Castro, Andrea Hartner, and Fabian B. Fahlbusch. 2020. "Expression of Retinoid Acid Receptor-Responsive Genes in Rodent Models of Placental Pathology" International Journal of Molecular Sciences 21, no. 1: 242. https://doi.org/10.3390/ijms21010242
APA StyleMocker, A., Schmidt, M., Huebner, H., Wachtveitl, R., Cordasic, N., Menendez-Castro, C., Hartner, A., & Fahlbusch, F. B. (2020). Expression of Retinoid Acid Receptor-Responsive Genes in Rodent Models of Placental Pathology. International Journal of Molecular Sciences, 21(1), 242. https://doi.org/10.3390/ijms21010242