Meniscus-Derived Matrix Bioscaffolds: Effects of Concentration and Cross-Linking on Meniscus Cellular Responses and Tissue Repair

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
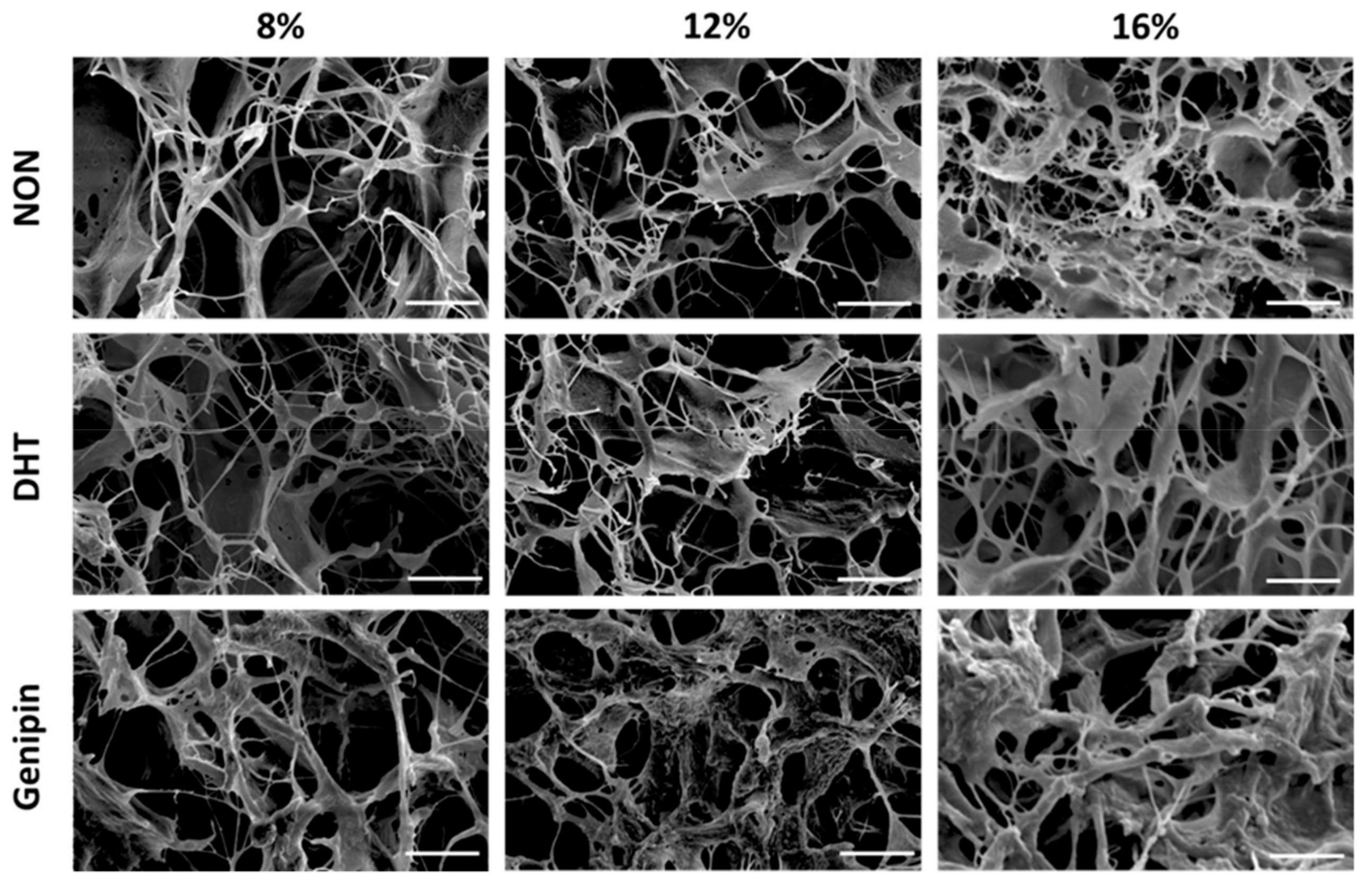
2.1. Scanning Electron Microscopy of MDM Scaffolds
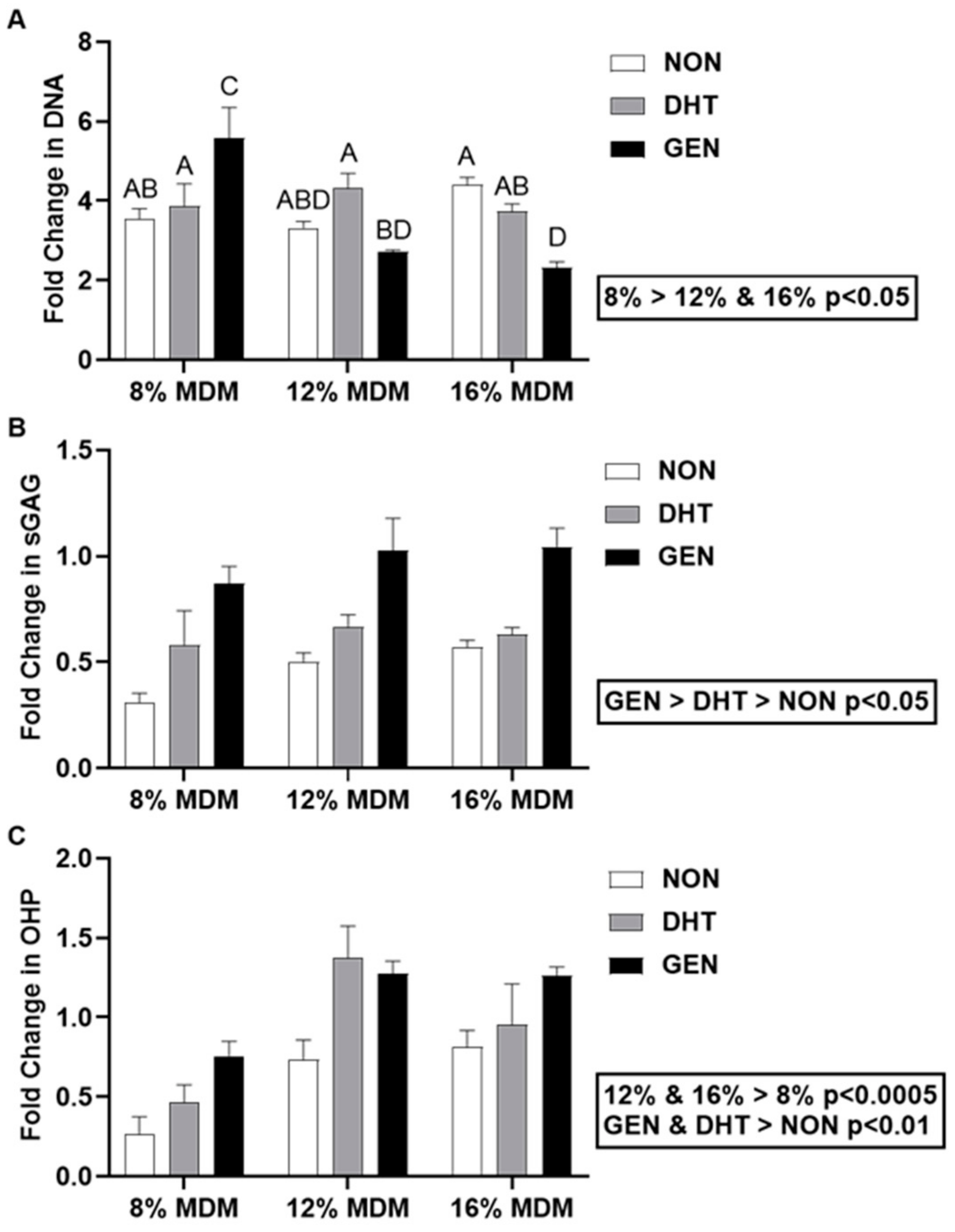
2.2. Biochemical Response of Primary Meniscus Cells Seeded on MDM Scaffolds
2.3. Meniscus Repair Model System
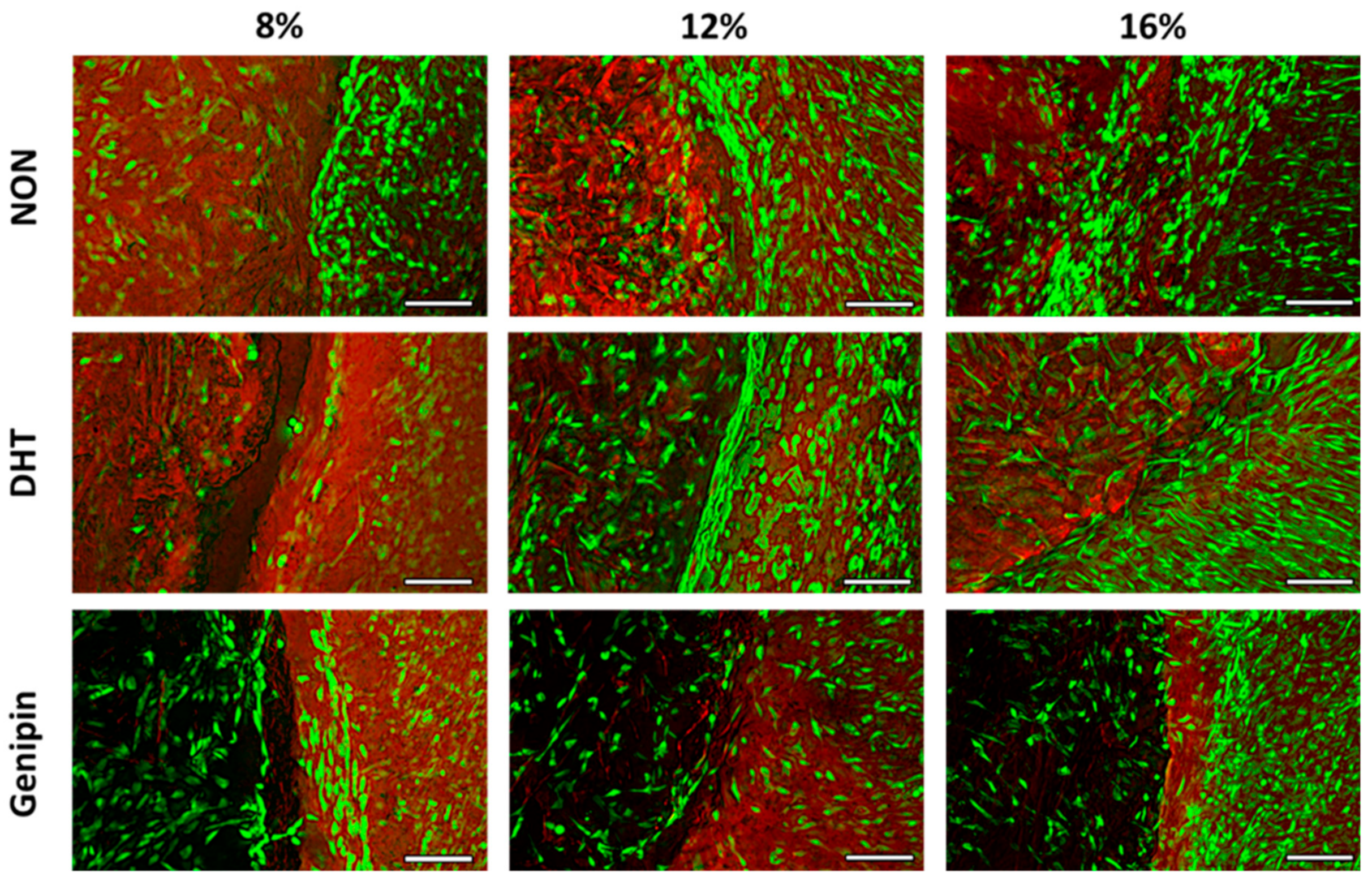
2.3.1. Fluorescence Imaging
2.3.2. Biochemical Analyses of MDM Scaffolds
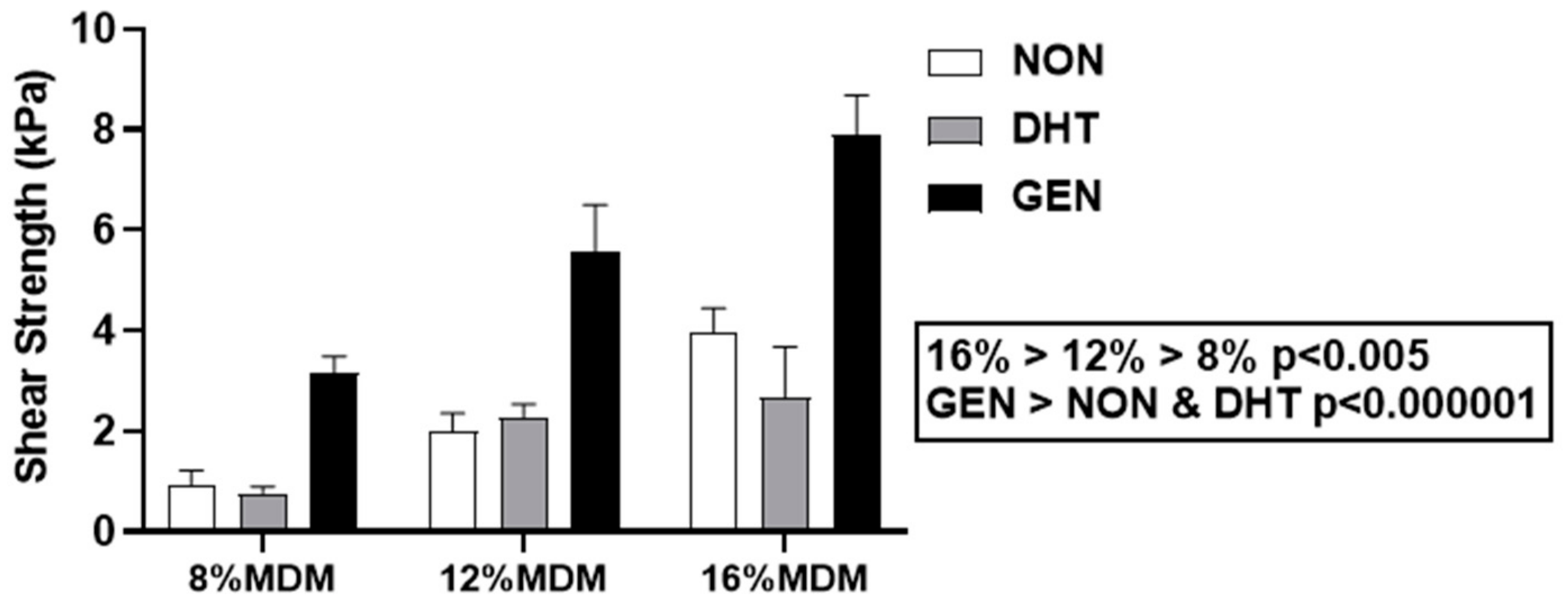
2.3.3. Integrative Shear Strength of Repair
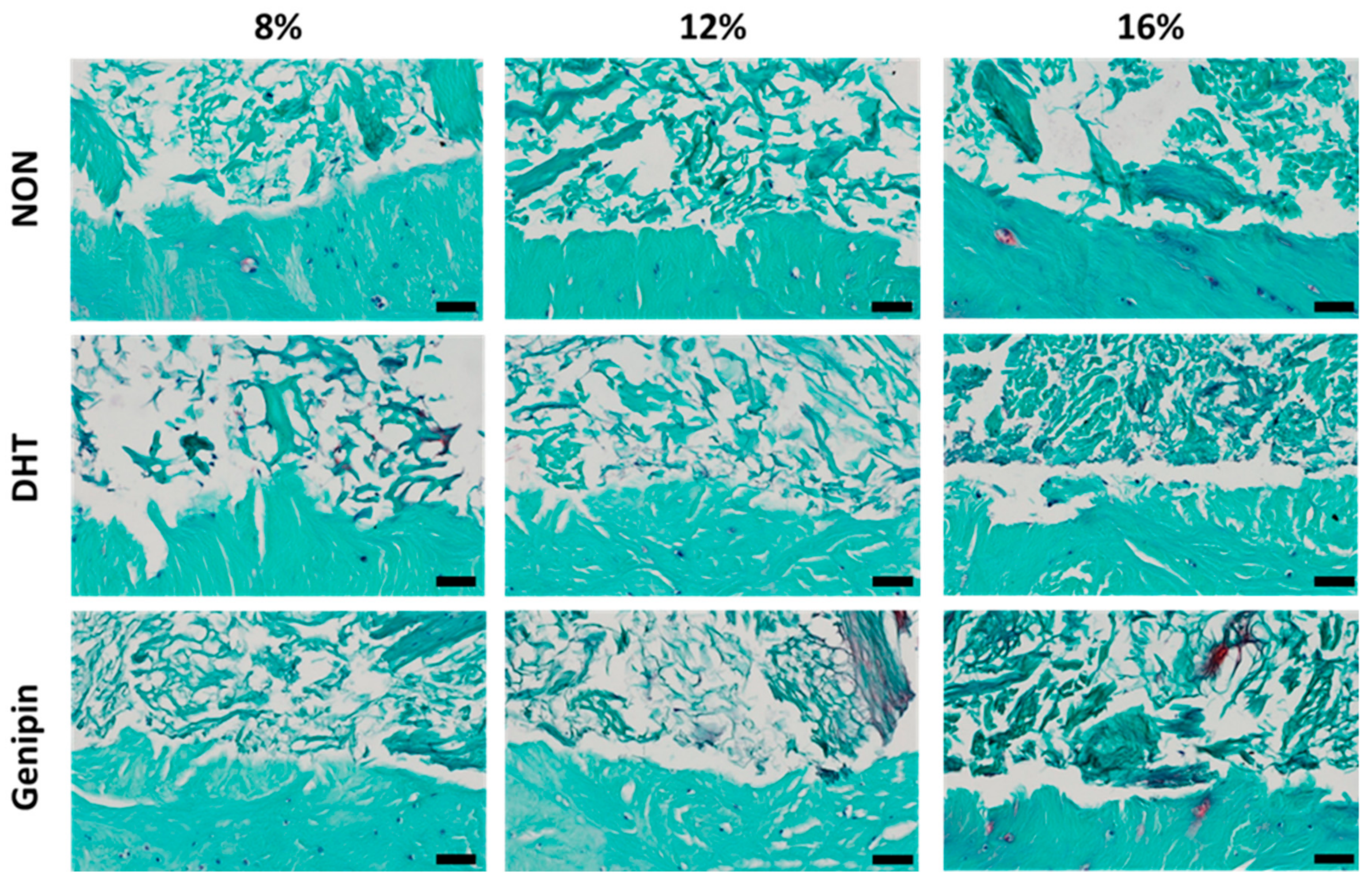
2.3.4. Histological Assessment
3. Discussion
4. Materials and Methods
4.1. Generation of MDM Scaffolds
4.2. Scanning Electron Microscopy
4.3. Isolation of Meniscal Cells
4.4. Primary Meniscus Cell Characterization on MDM Scaffolds
4.5. Meniscus Repair Model System
4.6. Fluorescent Imaging of Meniscus Repair Model Constructs
4.7. Integrative Shear Strength of Repair
4.8. Biochemical Analyses
4.9. Histological Analyses
4.10. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| ASC | adipose-derived stem cells |
| CDM | cartilage-derived matrix |
| DHT | dehydrothermal |
| DMEM-HG | Dulbecco’s modified eagle medium-high glucose |
| DMMB | dimethylmethylene blue |
| DNA | deoxyribonucleic acid |
| ECM | extracellular matrix |
| EDTA | ethylenediaminetetraacetic acid |
| EFI | extended focused imaging |
| FBS | fetal bovine serum |
| GEN | genipin cross-linked |
| HEPES | 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid |
| LSD | least significant difference |
| MDM | meniscus-derived matrix |
| hMSC | human mesenchymal stem cells |
| NON | non-cross-linked |
| OHP | hydroxyproline |
| PBS | phosphate buffered saline |
| PSF | penicillin streptomycin fungizone |
| SEM | standard error of the mean |
| sGAG | sulfated glycosaminoglycan |
| UV | ultraviolet |
References
- Blake, M.H.; Johnson, D.L. Knee Meniscus Injuries: Common Problems and Solutions. Clin. Sports Med. 2018, 37, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Carter, T.E.; Taylor, K.A.; Spritzer, C.E.; Utturkar, G.M.; Taylor, D.C.; Moorman, C.T.; Garrett, W.E.; Guilak, F.; McNulty, A.L.; DeFrate, L.E. In vivo cartilage strain increases following medial meniscal tear and correlates with synovial fluid matrix metalloproteinase activity. J. Biomech. 2015, 48, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Markolf, K.L.; Bargar, W.L.; Shoemaker, S.C.; Amstutz, H.C. The role of joint load in knee stability. J. Bone Joint Surg. Am. Vol. 1981, 63, 570–585. [Google Scholar] [CrossRef]
- Mauck, R.L.; Burdick, J.A. From repair to regeneration: Biomaterials to reprogram the meniscus wound microenvironment. Ann. Biomed. Eng. 2015, 43, 529–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scotti, C.; Hirschmann, M.T.; Antinolfi, P.; Martin, I.; Peretti, G.M. Meniscus repair and regeneration: Review on current methods and research potential. Eur. Cell Mater. 2013, 26, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.J.S.; Wanivenhaus, F.; Burge, A.J.; Warren, R.F.; Rodeo, S.A. The human meniscus: A review of anatomy, function, injury, and advances in treatment. Clin. Anat. 2015, 28, 269–287. [Google Scholar] [CrossRef]
- Proctor, C.S.; Schmidt, M.B.; Whipple, R.R.; Kelly, M.A.; Mow, V.C. Material properties of the normal medial bovine meniscus. J. Orthop. Res. 1989, 7, 771–782. [Google Scholar] [CrossRef]
- Liu, B.; Lad, N.K.; Collins, A.T.; Ganapathy, P.K.; Utturkar, G.M.; McNulty, A.L.; Spritzer, C.E.; Moorman, C.T.; Sutter, E.G.; Garrett, W.E.; et al. In vivo tibial cartilage strains in regions of cartilage-to-cartilage contact and cartilage-to-meniscus contact in response to walking. Am. J. Sports Med. 2017, 45, 2817–2823. [Google Scholar] [CrossRef]
- Lohmander, L.S.; Englund, P.M.; Dahl, L.L.; Roos, E.M. The long-term consequence of anterior cruciate ligament and meniscus injuries: Osteoarthritis. Am. J. Sports Med. 2007, 35, 1756–1769. [Google Scholar] [CrossRef] [Green Version]
- Badlani, J.T.; Borrero, C.; Golla, S.; Harner, C.D.; Irrgang, J.J. The effects of meniscus injury on the development of knee osteoarthritis: Data from the osteoarthritis initiative. Am. J. Sports Med. 2013, 41, 1238–1244. [Google Scholar] [CrossRef]
- Chung, K.S.; Ha, J.K.; Yeom, C.H.; Ra, H.J.; Jang, H.S.; Choi, S.H.; Kim, J.G. Comparison of Clinical and Radiologic Results Between Partial Meniscectomy and Refixation of Medial Meniscus Posterior Root Tears: A Minimum 5-Year Follow-up. Arthrosc. J. Arthrosc. Surg. 2015, 31, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Englund, M.; Lohmander, L.S. Risk factors for symptomatic knee osteoarthritis fifteen to twenty-two years after meniscectomy. Arthritis Rheum. 2004, 50, 2811–2819. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Choi, C.H.; Chun, Y.M.; Kim, S.H.; Lee, S.K.; Jang, J.; Jeong, H.; Jung, M. Relationship Between Preoperative Extrusion of the Medial Meniscus and Surgical Outcomes After Partial Meniscectomy. Am. J. Sport Med. 2017, 45, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.E.; Swiontkowski, M.F.; Weinstein, J.N.; Callaghan, J.; Rosier, R.N.; Berry, D.J.; Harrast, J.; Derosa, G.P. American board of orthopaedic surgery practice of the orthopaedic surgeon: Part-II, certification examination case mix. J. Bone Joint Surg. Am. Vol. 2006, 88, 660–667. [Google Scholar] [CrossRef]
- Arnoczky, S.P.; Warren, R.F. Microvasculature of the human meniscus. Am. J. Sports Med. 1982, 10, 90–95. [Google Scholar] [CrossRef]
- Noyes, F.R.; Barber-Westin, S.D. Repair of complex and avascular meniscal tears and meniscal transplantation. J. Bone Joint Surg. Am. Vol. 2010, 92, 1012–1029. [Google Scholar]
- Badylak, S.F. The extracellular matrix as a biologic scaffold material. Biomaterials 2007, 28, 3587–3593. [Google Scholar] [CrossRef]
- Hidalgo Perea, S.; Lyons, L.P.; Nishimuta, J.F.; Weinberg, J.B.; McNulty, A.L. Evaluation of culture conditions for in vitro meniscus repair model systems using bone marrow-derived mesenchymal stem cells. Connect. Tissue Res. 2019, 1–16. [Google Scholar] [CrossRef]
- Ruprecht, J.C.; Waanders, T.D.; Rowland, C.R.; Nishimuta, J.F.; Glass, K.A.; Stencel, J.; DeFrate, L.E.; Guilak, F.; Weinberg, J.B.; McNulty, A.L. Meniscus-Derived Matrix Scaffolds Promote the Integrative Repair of Meniscal Defects. Sci. Rep. 2019, 9, 8719. [Google Scholar] [CrossRef]
- Cheng, N.C.; Estes, B.T.; Awad, H.A.; Guilak, F. Chondrogenic differentiation of adipose-derived adult stem cells by a porous scaffold derived from native articular cartilage extracellular matrix. Tissue Eng. Part A 2009, 15, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Cheng, N.C.; Estes, B.T.; Young, T.H.; Guilak, F. Engineered cartilage using primary chondrocytes cultured in a porous cartilage-derived matrix. Regen. Med. 2011, 6, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, N.S.; Varghese, S.; Puleo, C.; Zhang, Z.; Elisseeff, J. Morphogenetic signals from chondrocytes promote chondrogenic and osteogenic differentiation of mesenchymal stem cells. J. Cell. Physiol. 2007, 212, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Rowland, C.R.; Lennon, D.P.; Caplan, A.I.; Guilak, F. The effects of crosslinking of scaffolds engineered from cartilage ECM on the chondrogenic differentiation of MSCs. Biomaterials 2013, 34, 5802–5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, N.C.; Estes, B.T.; Young, T.H.; Guilak, F. Genipin-crosslinked cartilage-derived matrix as a scaffold for human adipose-derived stem cell chondrogenesis. Tissue Eng. Part A 2013, 19, 484–496. [Google Scholar] [CrossRef] [Green Version]
- Cornwell, K.G.; Lei, P.; Andreadis, S.T.; Pins, G.D. Crosslinking of discrete self-assembled collagen threads: Effects on mechanical strength and cell–matrix interactions. J. Biomed. Mater. Res. A 2007, 80A, 362–371. [Google Scholar] [CrossRef]
- Lew, D.-H.; Liu, P.H.-T.; Orgill, D.P. Optimization of UV cross-linking density for durable and nontoxic collagen GAG dermal substitute. J. Biomed. Mater. Res. B Appl. Biomater. 2007, 82B, 51–56. [Google Scholar] [CrossRef]
- Schulz Torres, D.; Freyman, T.M.; Yannas, I.V.; Spector, M. Tendon cell contraction of collagen–GAG matrices in vitro: Effect of cross-linking. Biomaterials 2000, 21, 1607–1619. [Google Scholar] [CrossRef]
- Sung, H.-W.; Huang, D.-M.; Chang, W.-H.; Huang, R.-N.; Hsu, J.-C. Evaluation of gelatin hydrogel crosslinked with various crosslinking agents as bioadhesives: In vitro study. J. Biomed. Mater. Res. 1999, 46, 520–530. [Google Scholar] [CrossRef]
- Weadock, K.S.; Miller, E.J.; Bellincampi, L.D.; Zawadsky, J.P.; Dunn, M.G. Physical crosslinking of collagen fibers: Comparison of ultraviolet irradiation and dehydrothermal treatment. J. Biomed. Mater. Res. 1995, 29, 1373–1379. [Google Scholar] [CrossRef]
- Yan, L.-P.; Wang, Y.-J.; Ren, L.; Wu, G.; Caridade, S.G.; Fan, J.-B.; Wang, L.-Y.; Ji, P.-H.; Oliveira, J.M.; Oliveira, J.T.; et al. Genipin-cross-linked collagen/chitosan biomimetic scaffolds for articular cartilage tissue engineering applications. J. Biomed. Mater. Res. A 2010, 95A, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.W.; Huang, R.N.; Huang, L.L.; Tsai, C.C.; Chiu, C.T. Feasibility study of a natural crosslinking reagent for biological tissue fixation. J. Biomed. Mater. Res. 1998, 42, 560–567. [Google Scholar] [CrossRef]
- Cauich-Rodriguez, J.V.; Deb, S.; Smith, R. Effect of cross-linking agents on the dynamic mechanical properties of hydrogel blends of poly(acrylic acid)-poly(vinyl alcohol-vinyl acetate). Biomaterials 1996, 17, 2259–2264. [Google Scholar] [CrossRef]
- McNulty, A.L.; Guilak, F. Integrative repair of the meniscus: Lessons from in vitro studies. Biorheology 2008, 45, 487–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilusz, R.E.; Weinberg, J.B.; Guilak, F.; McNulty, A.L. Inhibition of integrative repair of the meniscus following acute exposure to interleukin-1 in vitro. J. Orthop. Res. 2008, 26, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Riera, K.M.; Rothfusz, N.E.; Wilusz, R.E.; Weinberg, J.B.; Guilak, F.; McNulty, A.L. Interleukin-1, tumor necrosis factor-alpha, and transforming growth factor-beta 1 and integrative meniscal repair: Influences on meniscal cell proliferation and migration. Arthritis Res. Ther. 2011, 13, R187. [Google Scholar] [CrossRef] [Green Version]
- McNulty, A.L.; Moutos, F.T.; Weinberg, J.B.; Guilak, F. Enhanced integrative repair of the porcine meniscus in vitro by inhibition of interleukin-1 or tumor necrosis factor alpha. Arthritis Rheum. 2007, 56, 3033–3042. [Google Scholar] [CrossRef]
- Hennerbichler, A.; Moutos, F.T.; Hennerbichler, D.; Weinberg, J.B.; Guilak, F. Interleukin-1 and tumor necrosis factor alpha inhibit repair of the porcine meniscus in vitro. Osteoarthr. Cartil. 2007, 15, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- McNulty, A.L.; Estes, B.T.; Wilusz, R.E.; Weinberg, J.B.; Guilak, F. Dynamic loading enhances integrative meniscal repair in the presence of interleukin-1. Osteoarthr. Cartil. 2010, 18, 830–838. [Google Scholar] [CrossRef] [Green Version]
- McNulty, A.L.; Weinberg, J.B.; Guilak, F. Inhibition of matrix metalloproteinases enhances in vitro repair of the meniscus. Clin. Orthop. Res. 2009, 467, 1557–1567. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Qu, F.; Han, B.; Wang, C.; Li, H.; Mauck, R.L.; Han, L. Micromechanical anisotropy and heterogeneity of the meniscus extracellular matrix. Acta Biomater. 2017, 54, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Rowland, C.R.; Colucci, L.A.; Guilak, F. Fabrication of anatomically-shaped cartilage constructs using decellularized cartilage-derived matrix scaffolds. Biomaterials 2016, 91, 57–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puetzer, J.L.; Bonassar, L.J. Physiologically Distributed Loading Patterns Drive the Formation of Zonally Organized Collagen Structures in Tissue-Engineered Meniscus. Tissue Eng. Part A 2016, 22, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.L.H.; Sung, H.-W.; Tsai, C.-C.; Huang, D.-M. Biocompatibility study of a biological tissue fixed with a naturally occurring crosslinking reagent. J. Biomed. Mater. Res. 1998, 42, 568–576. [Google Scholar] [CrossRef]
- Akao, T.; Kobashi, K.; Aburada, M. Enzymic Studies on the Animal and Intestinal Bacterial Metabolism of Geniposide. Biol. Pharm. Bull. 1994, 17, 1573–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, L.; Cao, Z.; Hu, Y.; Song, Y.; Yu, L.; Yang, B.; Mu, J.; Huang, Z.; Han, Y. Effects of different cross-linking conditions on the properties of genipin-cross-linked chitosan/collagen scaffolds for cartilage tissue engineering. J. Mater. Sci. Mater. Med. 2011, 22, 51–62. [Google Scholar] [CrossRef]
- Dimida, S.; Barca, A.; Cancelli, N.; De Benedictis, V.; Raucci, M.G.; Demitri, C. Effects of Genipin Concentration on Cross-Linked Chitosan Scaffolds for Bone Tissue Engineering: Structural Characterization and Evidence of Biocompatibility Features. Int. J. Polym. Sci. 2017, 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.; Lee, M.H.; Liang, H.C.; Hsu, C.K.; Sung, H.W. Acellular bovine pericardia with distinct porous structures fixed with genipin as an extracellular matrix. Tissue Eng. 2004, 10, 881–892. [Google Scholar] [CrossRef]
- Crapo, P.M.; Gilbert, T.W.; Badylak, S.F. An overview of tissue and whole organ decellularization processes. Biomaterials 2011, 32, 3233–3243. [Google Scholar] [CrossRef] [Green Version]
- Londono, R.; Dziki, J.L.; Haljasmaa, E.; Turner, N.J.; Leifer, C.A.; Badylak, S.F. The effect of cell debris within biologic scaffolds upon the macrophage response. J. Biomed. Mater. Res. Part A 2017, 105, 2109–2118. [Google Scholar] [CrossRef]
- Shimomura, K.; Rothrauff, B.B.; Tuan, R.S. Region-Specific Effect of the Decellularized Meniscus Extracellular Matrix on Mesenchymal Stem Cell-Based Meniscus Tissue Engineering. Am. J. Sports Med. 2017, 45, 604–611. [Google Scholar] [CrossRef]
- Rothrauff, B.B.; Shimomura, K.; Gottardi, R.; Alexander, P.G.; Tuan, R.S. Anatomical region-dependent enhancement of 3-dimensional chondrogenic differentiation of human mesenchymal stem cells by soluble meniscus extracellular matrix. Acta Biomater. 2017, 49, 140–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Chen, R.N.; Jhan, H.J.; Liu, D.Z.; Ho, H.O.; Mao, Y.; Kohn, J.; Sheu, M.T. Development and Characterization of Acellular Extracellular Matrix Scaffolds from Porcine Menisci for Use in Cartilage Tissue Engineering. Tissue Eng. Part C Methods 2015, 21, 971–986. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.; Levett, P.A.; te Moller, N.C.; Besems, J.; Boere, K.W.; van Rijen, M.H.; de Grauw, J.C.; Dhert, W.J.; van Weeren, P.R.; Malda, J. Crosslinkable hydrogels derived from cartilage, meniscus, and tendon tissue. Tissue Eng. Part A 2015, 21, 1195–1206. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ding, Q.; Dutta, A.; Wang, Y.; Huang, Y.H.; Weng, H.; Tang, L.; Hong, Y. An injectable extracellular matrix derived hydrogel for meniscus repair and regeneration. Acta Biomater. 2015, 16, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Nishimuta, J.F.; Levenston, M.E. Response of cartilage and meniscus tissue explants to in vitro compressive overload. Osteoarthr. Cartil. 2012, 20, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, F.; Guilak, F.; Mauck, R.L. Cell migration: Implications for repair and regeneration in joint disease. Nat. Rev. Rheumatol. 2019, 15, 167–179. [Google Scholar] [CrossRef]
- Abdelgaied, A.; Stanley, M.; Galfe, M.; Berry, H.; Ingham, E.; Fisher, J. Comparison of the biomechanical tensile and compressive properties of decellularised and natural porcine meniscus. J. Biomech. 2015, 48, 1389–1396. [Google Scholar] [CrossRef] [Green Version]
- Stabile, K.J.; Odom, D.; Smith, T.L.; Northam, C.; Whitlock, P.W.; Smith, B.P.; Van Dyke, M.E.; Ferguson, C.M. An acellular, allograft-derived meniscus scaffold in an ovine model. Arthroscopy 2010, 26, 936–948. [Google Scholar] [CrossRef]
- Maier, D.; Braeun, K.; Steinhauser, E.; Ueblacker, P.; Oberst, M.; Kreuz, P.C.; Roos, N.; Martinek, V.; Imhoff, A.B. In vitro analysis of an allogenic scaffold for tissue-engineered meniscus replacement. J. Orthop. Res. 2007, 25, 1598–1608. [Google Scholar] [CrossRef]
- Gorham, S.D.; Light, N.D.; Diamond, A.M.; Willins, M.J.; Bailey, A.J.; Wess, T.J.; Leslie, N.J. Effect of chemical modifications on the susceptibility of collagen to proteolysis. II. Dehydrothermal crosslinking. Int. J. Biol. Macromol. 1992, 14, 129–138. [Google Scholar] [CrossRef]
- McNulty, A.L.; Rothfusz, N.E.; Leddy, H.A.; Guilak, F. Synovial fluid concentrations and relative potency of interleukin-1 alpha and beta in cartilage and meniscus degradation. J. Orthop. Res. 2013, 31, 1039–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.T.; Hatcher, C.C.; Kim, S.Y.; Ziemian, S.N.; Spritzer, C.E.; Guilak, F.; DeFrate, L.E.; McNulty, A.L. Selective Enzymatic Digestion of Proteoglycans and Collagens Alters Cartilage T1rho and T2 Relaxation Times. Ann. Biomed. Eng. 2019, 47, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, C.C.; Collins, A.T.; Kim, S.Y.; Michel, L.C.; Mostertz, W.C.; Ziemian, S.N.; Spritzer, C.E.; Guilak, F.; DeFrate, L.E.; McNulty, A.L. Relationship between T1rho magnetic resonance imaging, synovial fluid biomarkers, and the biochemical and biomechanical properties of cartilage. J. Biomech. 2017, 55, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farndale, R.W.; Sayers, C.A.; Barrett, A.J. A direct spectrophotometric microassay for sulfated glycosaminoglycans in cartilage cultures. Connect. Tissue Res. 1982, 9, 247–248. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.K.; Enwemeka, C.S. A simplified method for the analysis of hydroxyproline in biological tissues. Clin. Biochem. 1996, 29, 225–229. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyons, L.P.; Hidalgo Perea, S.; Weinberg, J.B.; Wittstein, J.R.; McNulty, A.L. Meniscus-Derived Matrix Bioscaffolds: Effects of Concentration and Cross-Linking on Meniscus Cellular Responses and Tissue Repair. Int. J. Mol. Sci. 2020, 21, 44. https://doi.org/10.3390/ijms21010044
Lyons LP, Hidalgo Perea S, Weinberg JB, Wittstein JR, McNulty AL. Meniscus-Derived Matrix Bioscaffolds: Effects of Concentration and Cross-Linking on Meniscus Cellular Responses and Tissue Repair. International Journal of Molecular Sciences. 2020; 21(1):44. https://doi.org/10.3390/ijms21010044
Chicago/Turabian StyleLyons, Lucas P., Sofia Hidalgo Perea, J. Brice Weinberg, Jocelyn R. Wittstein, and Amy L. McNulty. 2020. "Meniscus-Derived Matrix Bioscaffolds: Effects of Concentration and Cross-Linking on Meniscus Cellular Responses and Tissue Repair" International Journal of Molecular Sciences 21, no. 1: 44. https://doi.org/10.3390/ijms21010044
APA StyleLyons, L. P., Hidalgo Perea, S., Weinberg, J. B., Wittstein, J. R., & McNulty, A. L. (2020). Meniscus-Derived Matrix Bioscaffolds: Effects of Concentration and Cross-Linking on Meniscus Cellular Responses and Tissue Repair. International Journal of Molecular Sciences, 21(1), 44. https://doi.org/10.3390/ijms21010044