Gadolinium Complexes as Contrast Agent for Cellular NMR Spectroscopy

Abstract
:
1. Introduction
2. Results
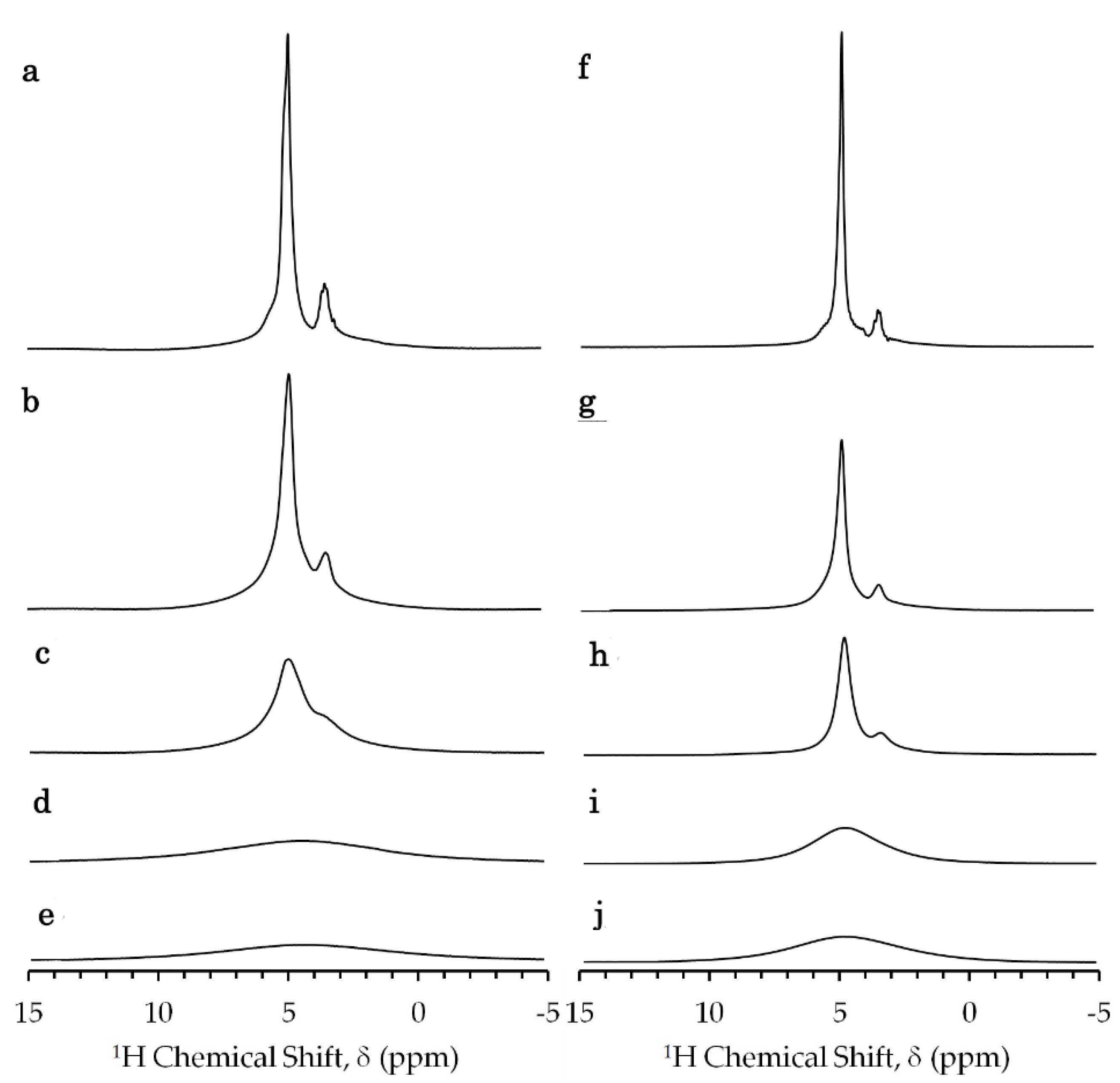
2.1. NMR Spectra of E. coli Sample in Absense of Gadolinium Complex
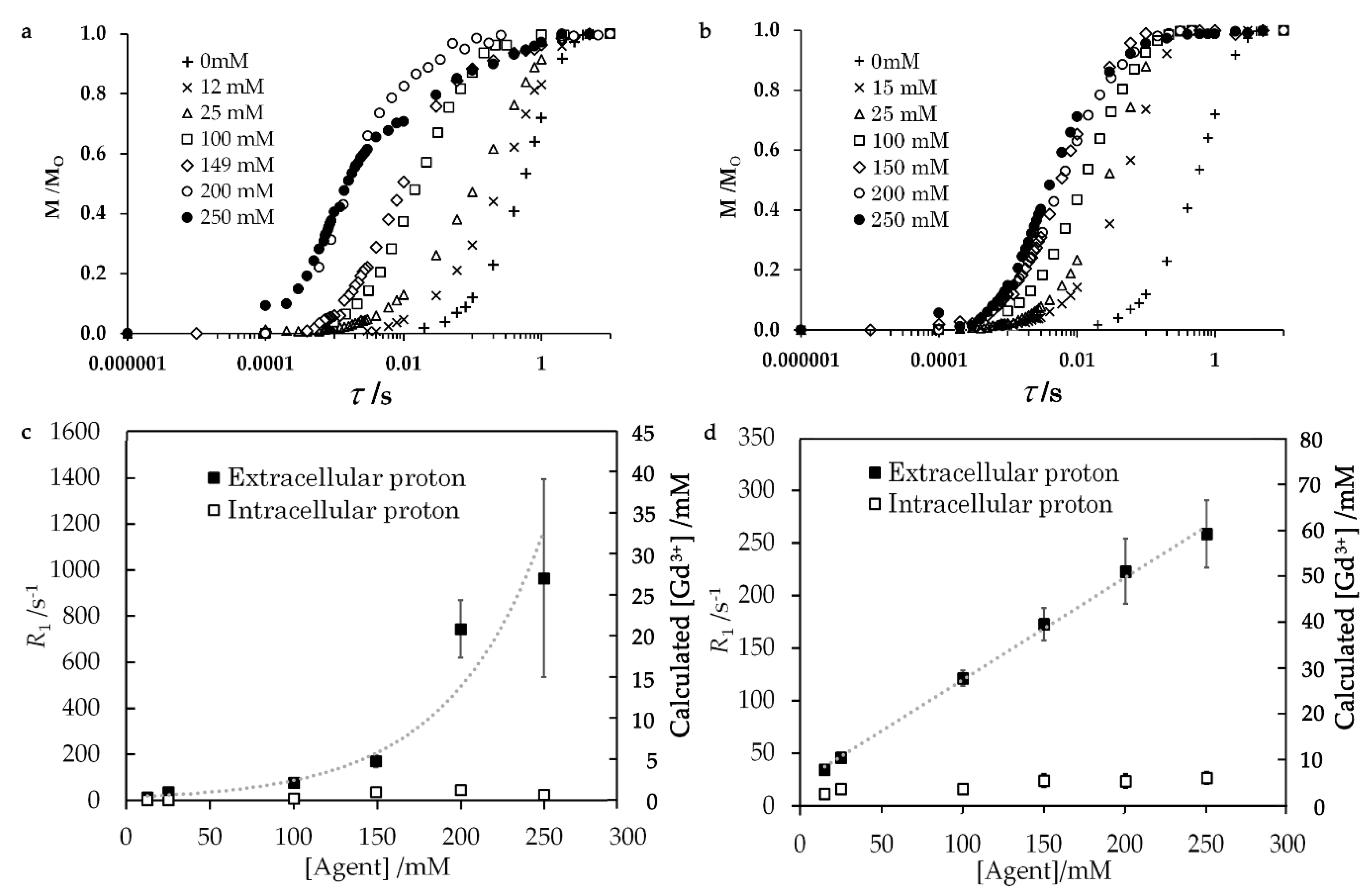
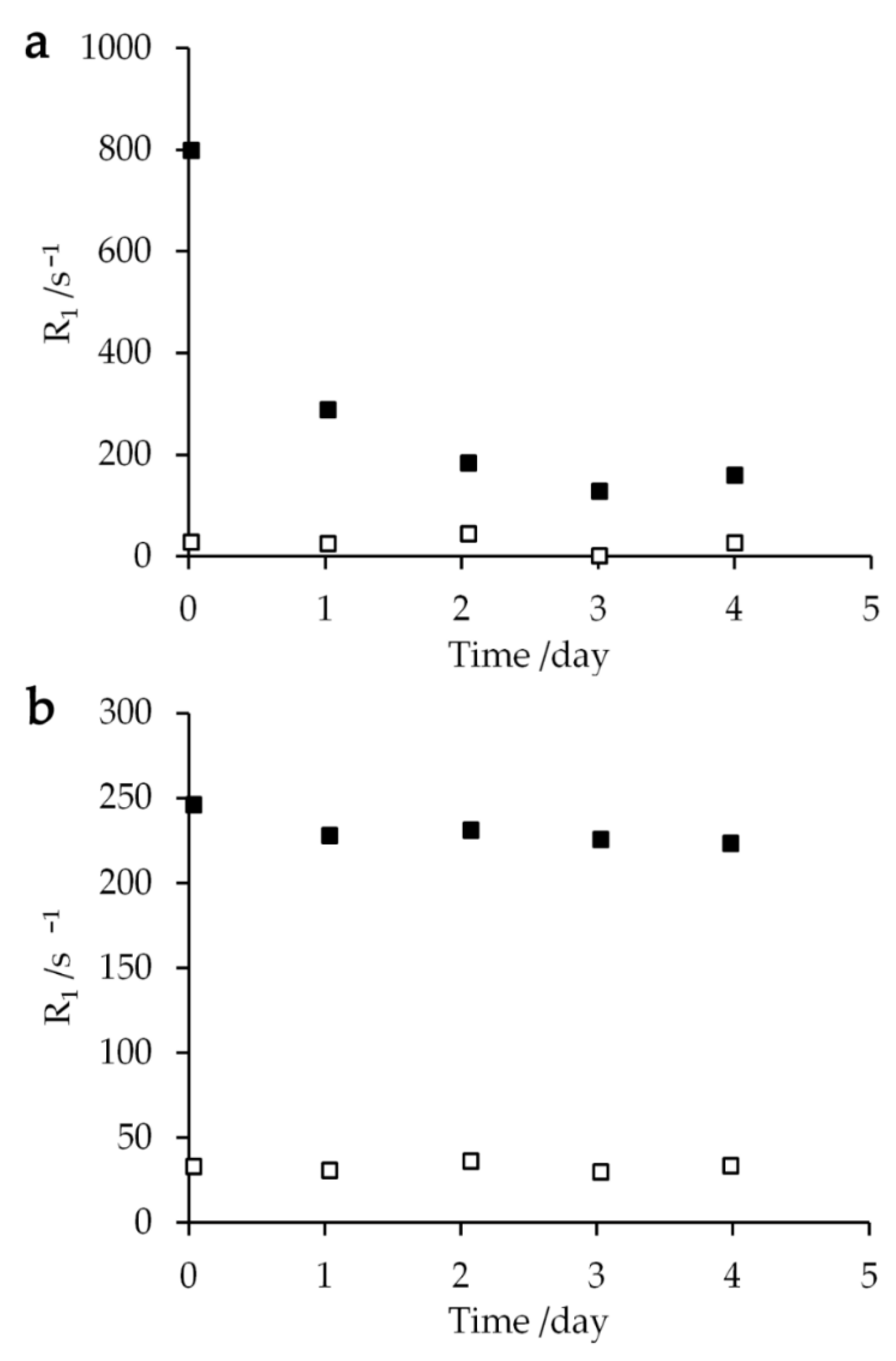
2.2. PRE for Gadolinium Solutions
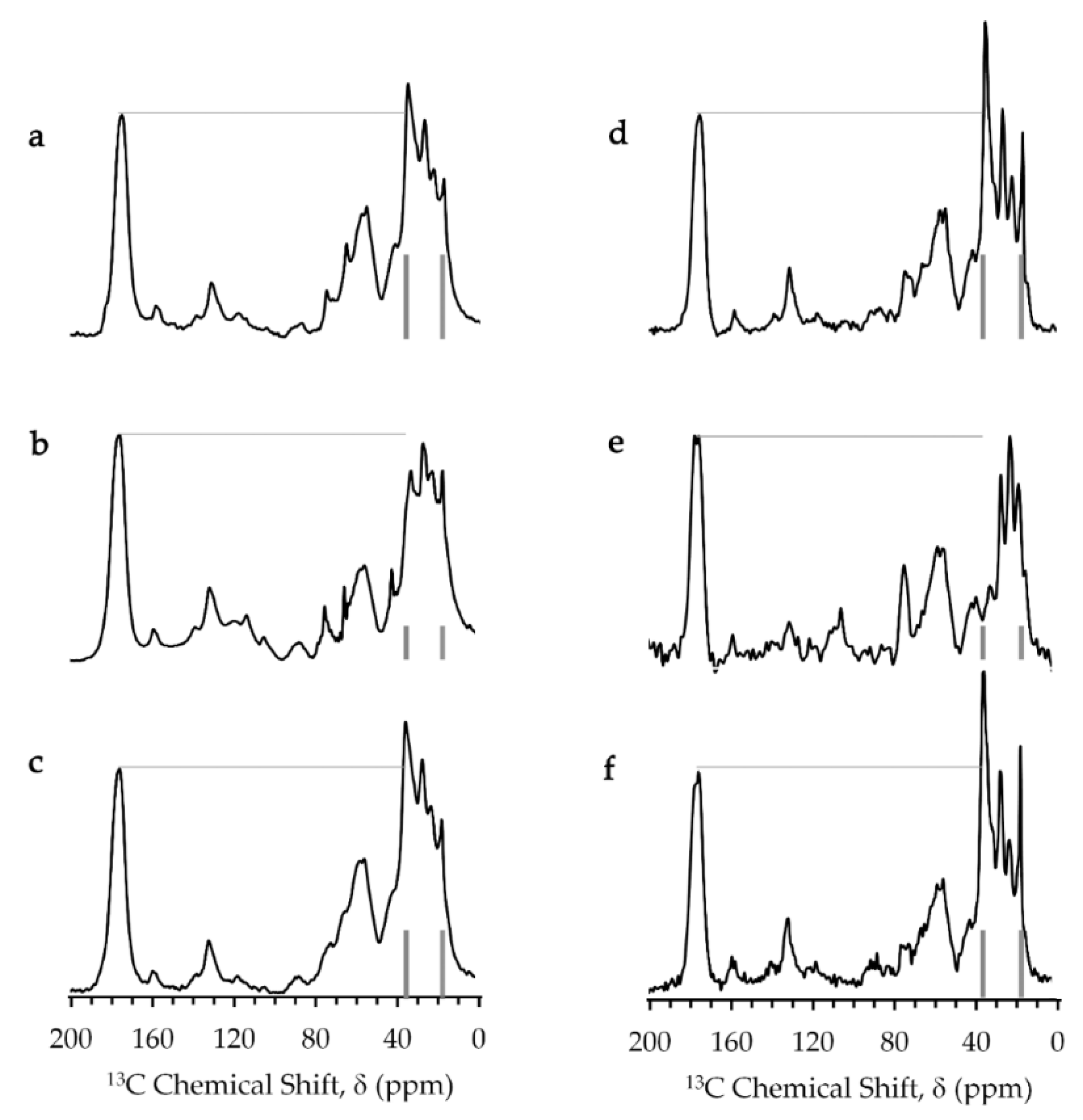
2.3. High-Resolution Solid-State 13C NMR Spectra
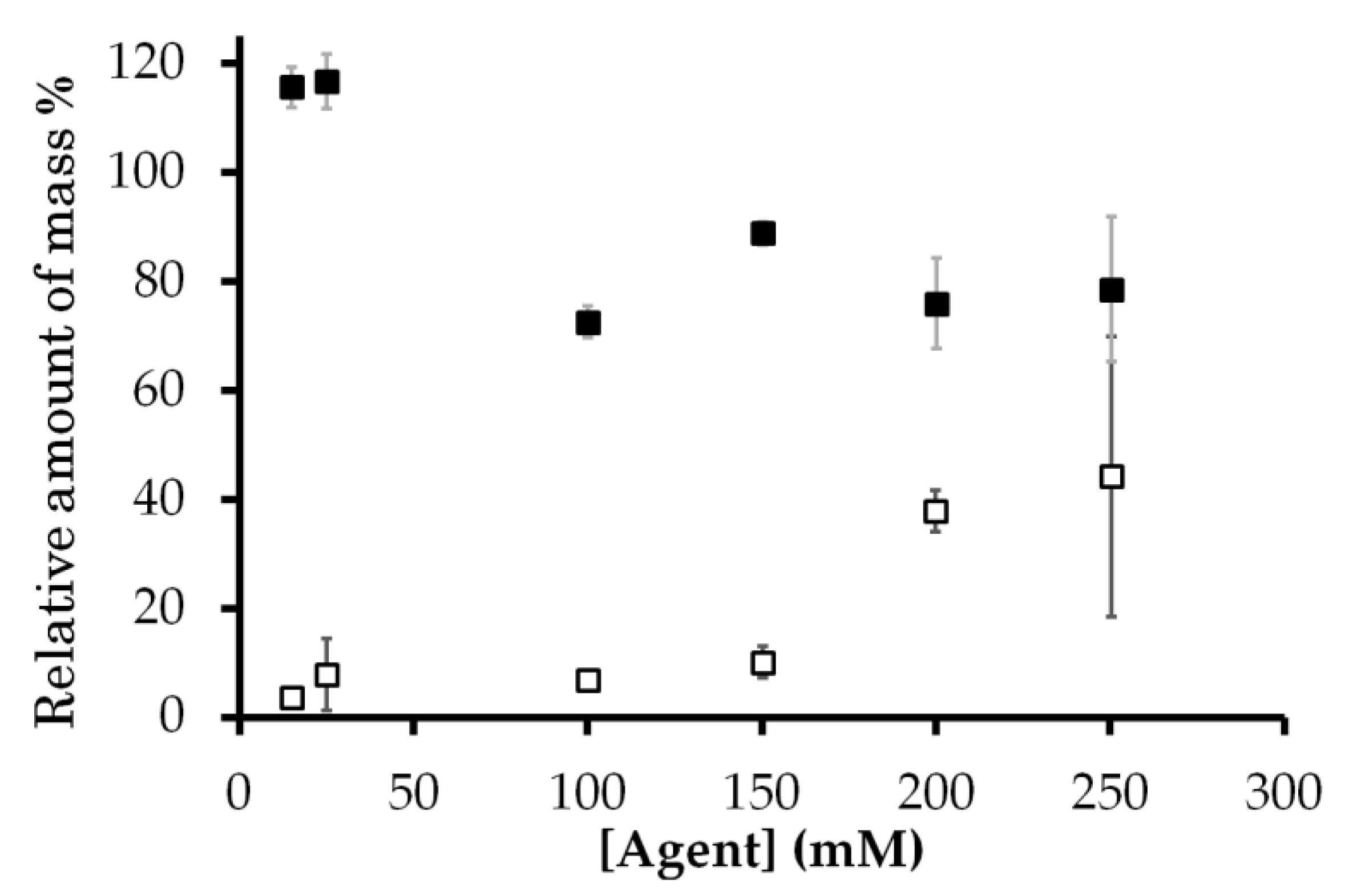
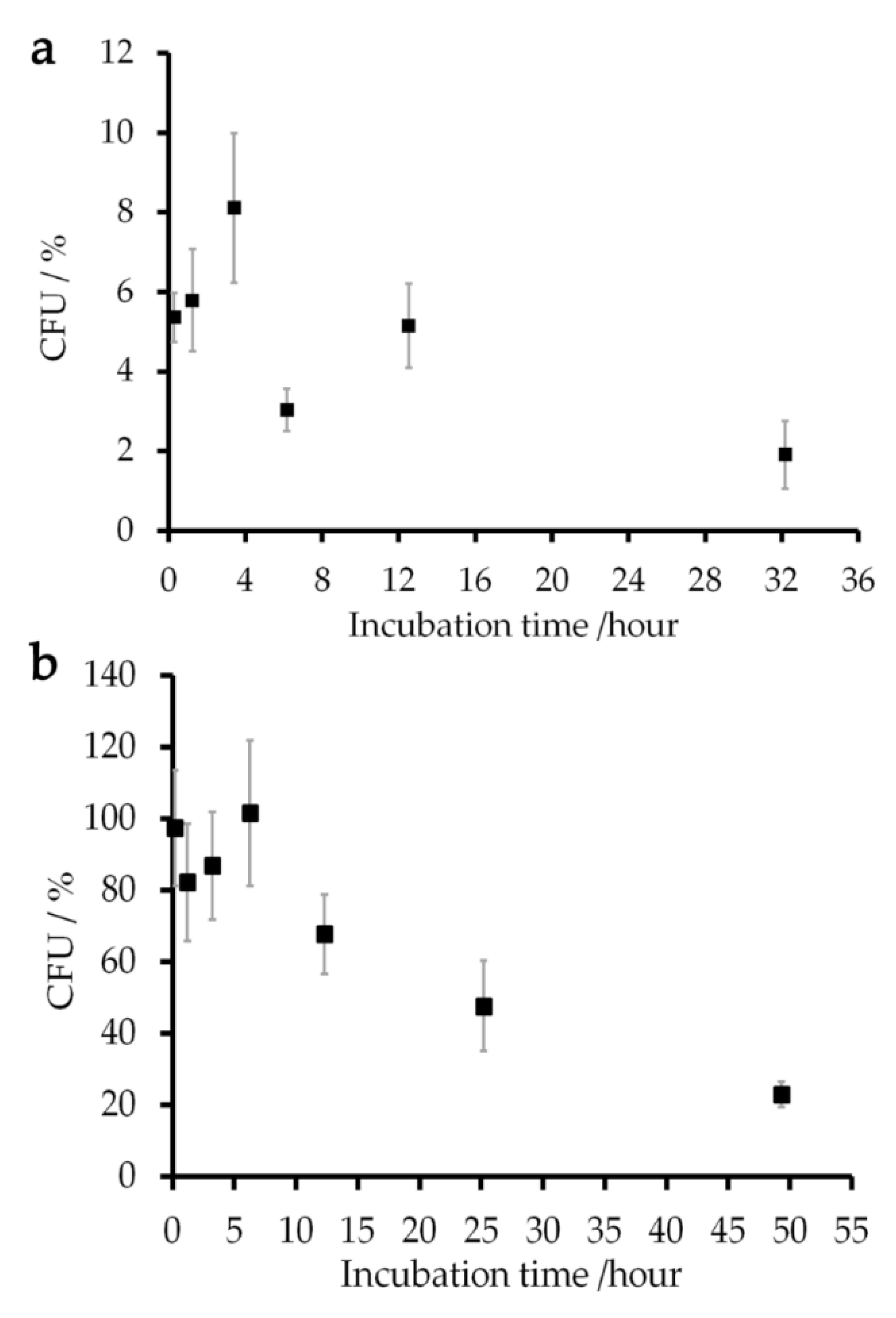
2.4. Viability of E. coli Cells and Magnetic Relxation during the Experiments
3. Discussion
3.1. Dependence of 1H-NMR Relaxation Rates R1 on Gd Complex Concentration
3.2. Interaction of Gd Complexes with the Cells and Cytotoxicity
3.3. Water Exchange Across the Cell Membranes and Its Effect on Water 1H T1 Relaxation
3.4. Cell Membrane Permeability to Gd Complexes
4. Materials and Methods
4.1. Data Analysis for the Longitudinal 1H Relaxation Rate R1 at High Magnetic Fields
4.2. Aqua Gd3+ and Gd-DOTA Solutions
4.3. E. coli Samples
4.4. Relaxation Measurement of the 1H Longitudinal Magnetization
4.5. Solid-State 13C-NMR Experiments
4.6. Viability of E. coli Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Gd-DOTA | Gadolinium-1,4,7,10-tetraazacyclododecane-1,4,7,10-tetraacete complex |
| NMR | Nuclear Magnetic Resonance |
| MRI | Magnetic Resonance Imaging |
| R1 | Longitudinal relaxation rate |
| T1 | Longitudinal relaxation time |
| R1p | Longitudinal relaxivity |
| PRE | Paramagnetic Relaxation Enhancement |
| CPMAS | Cross Polarization Magic-Angle Spinning |
| CFU | Colony-forming unit |
References
- Bertini, I.; Luchinat, C.; Parigi, G.; Ravera, E. NMR of Paramagnetic Molecules, Applications to Metallobiomolecules and Models, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 277–312. [Google Scholar]
- Pell, A.J.; Pintacuda, G.; Grey, C.P. Paramagnetic NMR in solution and the solid state. Prog. Nucl. Magn. Reson. Spectrosc. 2018, 111, 1–271. [Google Scholar] [CrossRef]
- Koehler, J.; Meiler, J. Expanding the utility of NMR restraints with paramagnetic compounds: Background and practical aspects. Prog. Nucl. Magn. Reson. Spectrosc. 2011, 59, 360–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.-Y.; Keizers, P.H.; Liu, W.-M.; Lohr, F.; Skinner, S.; Heeneman, E.A.; Schwalbe, H.; Ubbink, M.; Siegal, G. Small-molecule binding sites on proteins established by paramagnetic NMR spectroscopy. J. Am. Chem. Soc. 2013, 135, 5859–5868. [Google Scholar] [CrossRef]
- Maltsev, S.; Hudson, S.M.; Sahu, I.D.; Liu, L.; Lorigan, G.A. Solid-state NMR 31P paramagnetic relaxation enhancement membrane protein immersion depth measurements. J. Phys. Chem. B 2014, 118, 4370–4377. [Google Scholar] [CrossRef]
- Su, Y.; Mani, R.; Hong, M. Asymmetric insertion of membrane proteins in lipid bilayers by solid-state NMR paramagnetic relaxation enhancement: A cell-penetrating peptide example. J. Am. Chem. Soc. 2008, 130, 8856–8864. [Google Scholar] [CrossRef] [Green Version]
- Respondek, M.; Madl, T.; Göbl, C.; Golser, R.; Zangger, K. Mapping the orientation of helices in micelle-bound peptides by paramagnetic relaxation waves. J. Am. Chem. Soc. 2007, 129, 5228–5234. [Google Scholar] [CrossRef] [PubMed]
- Luchinat, C.; Parigi, G.; Ravera, E.; Rinaldelli, M. Solid-state NMR crystallography through paramagnetic restraints. J. Am. Chem. Soc. 2012, 134, 5006–5009. [Google Scholar] [CrossRef] [PubMed]
- Rotz, M.W.; Culver, K.S.B.; Parigi, G.; MacRenaris, K.W.; Luchinat, C.; Odom, T.W.; Meade, T.J. High relaxivity Gd(III)–DNA gold nanostars: Investigation of shape effects on proton relaxation. ACS Nano 2015, 9, 3385–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianolio, E.; Porto, S.; Napolitano, R.; Baroni, S.; Giovenzana, G.B.; Aime, S. Relaxometric investigations and MRI evaluation of a liposome-loaded pH-responsive Gadolinium(III) complex. Inorg. Chem. 2012, 51, 7210–7217. [Google Scholar] [CrossRef]
- Morrow, J.R.; Tóth, É. Next-generation magnetic resonance imaging contrast agents. Inorg. Chem. 2017, 56, 6029–6034. [Google Scholar] [CrossRef] [Green Version]
- Boros, E.; Gale, E.M.; Caravan, P. MR imaging probes: Design and applications. Dalton Trans. 2015, 44, 4804–4818. [Google Scholar] [CrossRef] [PubMed]
- De Leon-Rodriguez, L.M.; Martins, A.F.; Pinho, M.; Rofsky, N.M.; Sherry, A.D. Basic MR relaxation mechanisms and contrast agent design. J. Magn. Reson. Imaging 2015, 42, 545–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherry, A.D.; Caravan, P.; Lenkinski, R. Primer on gadolinium chemistry. J. Magn. Reson. Imaging 2009, 30, 1240–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanana, H.; Turcotte, P.; André, C.; Gagnon, C.; Gagné, F. Comparative study of the effects of gadolinium chloride and gadolinium—Based magnetic resonance imaging contrast agent on freshwater mussel, Dreissena polymorpha. Chemosphere 2017, 181, 197–207. [Google Scholar] [CrossRef]
- Ariyani, W.; Khairinisa, M.A.; Perrotta, G.; Manto, M.; Koibuchi, N. The effects of gadolinium-based contrast agents on the cerebellum: From basic research to neurological practice and from pregnancy to adulthood. Cerebellum 2017, 17, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Jones, C.M.; Chan, W.T.K.; Pham, T.A.; Ling, X.; Gale, E.M.; Rotile, N.J.; Tai, W.C.-S.; Anderson, C.J.; Caravan, P.; et al. Chiral DOTA chelators as an improved platform for biomedical imaging and therapy applications. Nat. Commun. 2018, 9, 857. [Google Scholar] [CrossRef] [Green Version]
- Granata, V.; Cascella, M.; Fusco, R.; Dell’Aprovitola, N.; Catalano, O.; Filice, S.; Schiavone, V.; Izzo, F.; Cuomo, A.; Petrillo, A. Immediate adverse reactions to gadolinium-based MR contrast media: A retrospective analysis on 10,608 examinations. Biomed. Res. Int. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Ramalho, J.; Semelka, R.C.; Ramalho, M.; Nunes, R.H.; Alobaidy, M.; Castillo, M. Gadolinium-based contrast agent accumulation and toxicity: An update. Am. J. Neuroradiol. 2015, 37, 1192–1198. [Google Scholar] [CrossRef] [Green Version]
- Cabella, C.; Crich, S.G.; Corpillo, D.; Barge, A.; Ghirelli, C.; Bruno, E.; Lorusso, V.; Uggeri, F.; Aime, S. Cellular labeling with Gd(III) chelates: Only high thermodynamic stabilities prevent the cells acting as ‘sponges’ of Gd3+ ions. Contrast Media Mol. Imaging 2006, 1, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Crich, S.G.; Biancone, L.; Cantaluppi, V.; Duò, D.; Esposito, G.; Russo, S.; Camussi, G.; Aime, S. Improved route for the visualization of stem cells labeled with a Gd-/Eu-chelate as dual (MRI and fluorescence) agent. Magn. Reson. Med. 2004, 51, 938–944. [Google Scholar] [CrossRef]
- Modo, M.; Mellodew, K.; Cash, D.; Fraser, S.E.; Meade, T.J.; Price, J.R.; Williams, S.C. Mapping transplanted stem cell migration after a stroke: A serial, in vivo magnetic resonance imaging study. NeuroImage 2004, 21, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Vuu, K.; Xie, J.; McDonald, M.A.; Bernardo, M.; Hunter, F.; Zhang, Y.; Li, K.; Bednarski, M.; Guccione, S. Gadolinium-rhodamine nanoparticles for cell labeling and tracking via magnetic resonance and optical imaging. Bioconjugate Chem. 2005, 16, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Luchinat, E.; Banci, L. In-cell NMR in human cells: Direct protein expression allows structural studies of protein folding and maturation. Acc. Chem. Res. 2018, 51, 1550–1557. [Google Scholar] [CrossRef]
- Sakakibara, D.; Sasaki, A.; Ikeya, T.; Hamatsu, J.; Hanashima, T.; Mishima, M.; Yoshimasu, M.; Hayashi, N.; Mikawa, T.; Wälchli, M.; et al. Protein structure determination in living cells by in-cell NMR spectroscopy. Nature 2009, 458, 102–105. [Google Scholar] [CrossRef]
- Rahman, S.; Byun, Y.; Hassan, I.; Kim, J.; Kumar, V. Towards understanding cellular structure biology: In-cell NMR. Biochim. Biophys. Acta 2017, 1865, 547–557. [Google Scholar] [CrossRef]
- Lippens, G.; Cahoreau, E.; Millard, P.; Charlier, C.; Lopez, J.M.; Hanoulle, X.; Portais, J.-C. In-cell NMR: From metabolites to macromolecules. Analyst 2018, 143, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Luchinat, E.; Banci, L. In-cell NMR: A topical review. IUCrJ 2017, 4, 108–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebau, J.; Mäler, L. Immersion depths of lipid carbons in bicelles measured by paramagnetic relaxation enhancement. J. Phys. Chem. B 2017, 121, 7660–7670. [Google Scholar] [CrossRef]
- Wang, S.; Gopinath, T.; Veglia, G. Application of paramagnetic relaxation enhancements to accelerate the acquisition of 2D and 3D solid-state NMR spectra of orientedmembrane proteins. Methods 2018, 138–139, 54–61. [Google Scholar] [CrossRef]
- Phillips, R.; Kondev, J.; Theriot, J.; Garcia, H.G.; Orme, N. Physical Biology of the Cell, 2nd ed.; Garland Science: New York, NY, USA, 2012; pp. 3–85. [Google Scholar]
- Sundararaj, S.; Guo, A.; Habibi-Nazhad, B.; Rouani, M.; Stothard, P.; Ellison, M.; Wishart, D.S. The CyberCell Database (CCDB): A comprehensive, self-updating, relational database to coordinate and facilitate in silico modeling of Escherichia coli. Nucleic Acids Res. 2004, 32, D293–D295. [Google Scholar] [CrossRef] [Green Version]
- Persson, E.; Halle, B. Cell water dynamics on multiple time scales. Proc. Natl. Acad. Sci. USA 2008, 105, 6266–6271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qvist, J.; Persson, E.; Mattea, C.; Halle, B. Time scales of water dynamics at biological interfaces: Peptides, proteins and cells. Faraday Discuss. 2009, 141, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Renault, M.A.M.; Pawsey, S.; Bos, M.P.; Koers, E.; Nand, D.; Boxtel, R.T.; Rosay, M.; Tommassen, J.; Maas, W.E.; Baldus, M.; et al. Solid-state NMR spectroscopy on cellular preparations enhanced by dynamic nuclear polarization. Angew. Chem. Int. Ed. 2012, 51, 2998–3001. [Google Scholar] [CrossRef] [PubMed]
- van Dalen, A.; de Kruijff, B. The role of lipids in membrane insertion and translocation of bacterial proteins. Biochim. Biophys. Acta 2004, 1694, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, W.E. Nuclear spin-lattice relaxation caused by paramagnetic impurities. Phys. Rev. 1960, 119, 79–84. [Google Scholar] [CrossRef]
- Nadaud, P.S.; Helmus, J.J.; Kall, S.L.; Jaroniec, C.P. Paramagnetic ions enable tuning of nuclear relaxation rates and provide long-range structural restraints in solid-state NMR of proteins. J. Am. Chem. Soc. 2009, 131, 8108–8120. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C. Solid state 13C NMR of unlabeled phosphatidylcholine bilayers: Spectral assignments and measurement of carbon-phosphorus dipolar couplings and 13C chemical shift anisotropies. Biophys. J. 1993, 64, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Laage, S.; Tao, Y.; McDermott, A.E. Cardiolipin interaction with subunit c of ATP synthase: Solid-state NMR characterization. Biochim. Biophys. Acta 2014, 1848, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boettcher, J.M.; Davis-Harrison, R.L.; Clay, M.C.; Nieuwkoop, A.J.; Ohkubo, Y.Z.; Tajkhorshid, E.; Morrissey, J.H.; Rienstra, C.M. Atomic view of calcium-induced clustering of phosphatidylserine in mixed lipid bilayers. Biochemistry 2011, 50, 2264–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, J.E.; LeComte, J.T.J.; Lee, A.L.; Young, G.B.; Pielak, G.J. Protein dynamics in living cells. Biochemistry 2005, 44, 9275–9279. [Google Scholar] [CrossRef] [PubMed]
- Jucker, B.A.; Harms, H.; Zehnder, A.J. Adhesion of the positively charged bacterium Stenotrophomonas (Xanthomonas) maltophilia 70401 to glass and Teflon. J. Bacteriol. 1996, 178, 5472–5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, K.; Elwing, H.; Hermansson, M. Effect of ionic strength on initial interactions of Escherichia coli with surfaces, studied on-line by a novel quartz crystal microbalance technique. J. Bacteriol. 1999, 181, 5210–5218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagle, J.F.; Nagle, J.F. Lipid bilayer structure. Curr. Opin. Struct. Biol. 2000, 10, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, B.R.; Maruyama, K.; Edula, J.R.; Tougan, T.; Lin, Y.; Lee, Y.-H.; Horii, T.; Fujiwara, T. Mechanistic and structural basis of bioengineered bovine Cathelicidin-5 with optimized therapeutic activity. Sci. Rep. 2017, 7, 44781. [Google Scholar] [CrossRef] [Green Version]
- Fuma, S.; Takeda, H.; Miyamoto, K.; Yanagisawa, K.; Inoue, Y.; Ishii, N.; Sugai, K.; Ishii, C.; Kawabata, Z. Ecological evaluation of gadolinium toxicity compared with other heavy metals using an aquatic microcosm. Bull. Environ. Contam. Toxicol. 2001, 66, 231–238. [Google Scholar] [CrossRef]
- Ishiguchi, T.; Takahashi, S. Safety of gadoterate meglumine (Gd-DOTA) as a contrast agent for magnetic resonance imaging: Results of a post-marketing surveillance study in Japan. Drugs R D 2010, 10, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Hazlewood, C.F.; Chang, D.C.; Nichols, B.L.; Woessner, N.E. Nuclear magnetic resonance transverse relaxation times of water protons in skeletal muscle. Biophys. J. 1974, 14, 583–606. [Google Scholar] [CrossRef] [Green Version]
- Donahue, K.M.; Weisskoff, R.M.; Chesler, D.A.; Kwong, K.K.; Bogdanov, A.A.; Mandeville, J.B.; Rosen, B.R. Improving MR quantification of regional blood volume with intravascularT1 contrast agents: Accuracy, precision, and water exchange. Magn. Reson. Med. 1996, 36, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Fick, A. On liquid diffusion. J. Membr. Sci. 1995, 100, 33–38. [Google Scholar] [CrossRef]
- Donahue, K.M.; Weisskoff, R.M.; Burstein, D. Water diffusion and exchange as they influence contrast enhancement. J. Magn. Reson. Imaging 1997, 7, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Labadie, C.; Lee, J.; Vetek, G.; Springer, C. Relaxographic Imaging. J. Magn. Reson. Ser. B 1994, 105, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Aslund, I.; Nowacka, I.A.; Nilsson, M.; Topgaard, D.; Hürlimann, M.D.; Song, Y.Q.; Fantazzini, P.; Bortolotti, V. Molecular exchange between intra- and extracellular compartments in a cell suspension. AIP Conf. Proc. 2008, 1081, 79–82. [Google Scholar]
- Iwahara, J.; Schwieters, C.D.; Clore, G.M. Ensemble approach for NMR structure refinement against1H paramagnetic relaxation enhancement data arising from a flexible paramagnetic group attached to a macromolecule. J. Am. Chem. Soc. 2004, 126, 5879–5896. [Google Scholar] [CrossRef] [PubMed]
- Iwahara, J.; Tang, C.; Clore, G.M. Practical aspects of 1H transverse paramagnetic relaxation enhancement measurements on macromolecules. J. Magn. Reson. 2006, 184, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazyev, O.V.; Helm, L.; Malkin, V.G.; Malkina, O.L. Quantum chemical investigation of hyperfine coupling constants on first coordination sphere water molecule of Gadolinium(III) aqua complexes. J. Phys. Chem. A 2005, 109, 10997–11005. [Google Scholar] [CrossRef] [PubMed]
- Luz, Z.; Meiboom, S. Proton relaxation in dilute solutions of Cobalt(II) and Nickel(II) ions in methanol and the rate of methanol exchange of the solvation sphere. J. Chem. Phys. 1964, 40, 2686. [Google Scholar] [CrossRef]
- Swift, T.J.; Connick, R.E. NMR-relaxation mechanisms of O 17 in aqueous solutions of paramagnetic cations and the lifetime of water molecules in the first coordination sphere. J. Chem. Phys. 1962, 37, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Caravan, P. Strategies for increasing the sensitivity of gadolinium based MRI contrast agents. Chem. Soc. Rev. 2006, 35, 512–523. [Google Scholar] [CrossRef]
- Benmelouka, M.; Borel, A.; Moriggi, L.; Helm, L.; Merbach, A.E. Design of Gd(III)-based magnetic resonance imaging contrast agents: Static and transient zero-field splitting contributions to the electronic relaxation and their impact on relaxivity. J. Phys. Chem. B 2007, 111, 832–840. [Google Scholar] [CrossRef]
- Banci, L.; Bertini, I.; Luchinat, C. 1H NMRD studies of solutions of paramagnetic metal ions in ethyleneglycol. Inorg. Chim. Acta 1985, 100, 173–181. [Google Scholar] [CrossRef]
- Taupitz, M.; Stolzenburg, N.; Ebert, M.; Schnorr, J.; Hauptmann, R.; Kratz, H.; Hamm, B.; Wagner, S. Gadolinium-containing magnetic resonance contrast media: Investigation on the possible transchelation of Gd3+to the glycosaminoglycan heparin. Contrast Media Mol. Imaging 2012, 8, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Micskei, K.; Powell, D.H.; Helm, L.; Brücher, E.; Merbach, A.E. Water exchange on [Gd(H2O)8]3+ and [Gd(PDTA)(H2O)] in aqueous solution: A variable-pressure, -temperature and -magnetic field17O NMR study. Magn. Reson. Chem. 1993, 31, 1011–1020. [Google Scholar] [CrossRef]
- Powell, D.; González, G.; Tissières, V.; Micskei, K.; Brücher, E.; Helm, L.; Merbach, A. Gd3+ chelates of interest in magnetic resonance imaging (MRI): Studies using 17O NMR and EPR at several magnetic fields. J. Alloy. Compd. 1994, 207, 20–24. [Google Scholar] [CrossRef]
- Hindman, J.C. Relaxation processes in water. A study of the proton spin-lattice relaxation time. J. Chem. Phys. 1973, 59, 1517. [Google Scholar] [CrossRef]
- Morokuma, K. Molecular orbital studies of hydrogen bonds: Dimeric H2O with the slater minimal basis set. J. Chem. Phys. 1970, 52, 1301. [Google Scholar] [CrossRef]
- Fogel’Son, R.L.; Likhachev, E.R. Temperature dependence of viscosity. Tech. Phys. 2001, 46, 1056–1059. [Google Scholar] [CrossRef]
- Baldwin, W.W.; Myer, R.; Powell, N.; Anderson, E.; Koch, A.L. Buoyant density of Escherichia coli is determined solely by the osmolarity of the culture medium. Arch. Microbiol. 1995, 164, 155–157. [Google Scholar] [CrossRef]
- Doerrler, W.T. Density gradient enrichment of escherichia coli conditional msbA mutants. Appl. Environ. Microbiol. 2007, 73, 7992–7996. [Google Scholar] [CrossRef] [Green Version]
- Vishnevskaya, G.P.; Frolova, E.N.; Gataullin, A.M.; Gumerov, F.M.; Fakhrutdinov, A.R. Structural transformations and phase transitions in aqueous solutions of gadolinium nitrate in the course of freezing. Phys. Solid State 2003, 45, 1322–1326. [Google Scholar] [CrossRef]
- Koenig, S.H.; Epstein, M. Ambiguities in the interpretation of proton magnetic relaxation data in water solutions of Gd3+ ions. J. Chem. Phys. 1975, 63, 2279. [Google Scholar] [CrossRef]
- Tei, L.; Baranyai, Z.; Gaino, L.; Forgács, A.; Vágner, A.; Botta, M. Thermodynamic stability, kinetic inertness and relaxometric properties of monoamide derivatives of lanthanide(iii) DOTA complexes. Dalton Trans. 2015, 44, 5467–5478. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
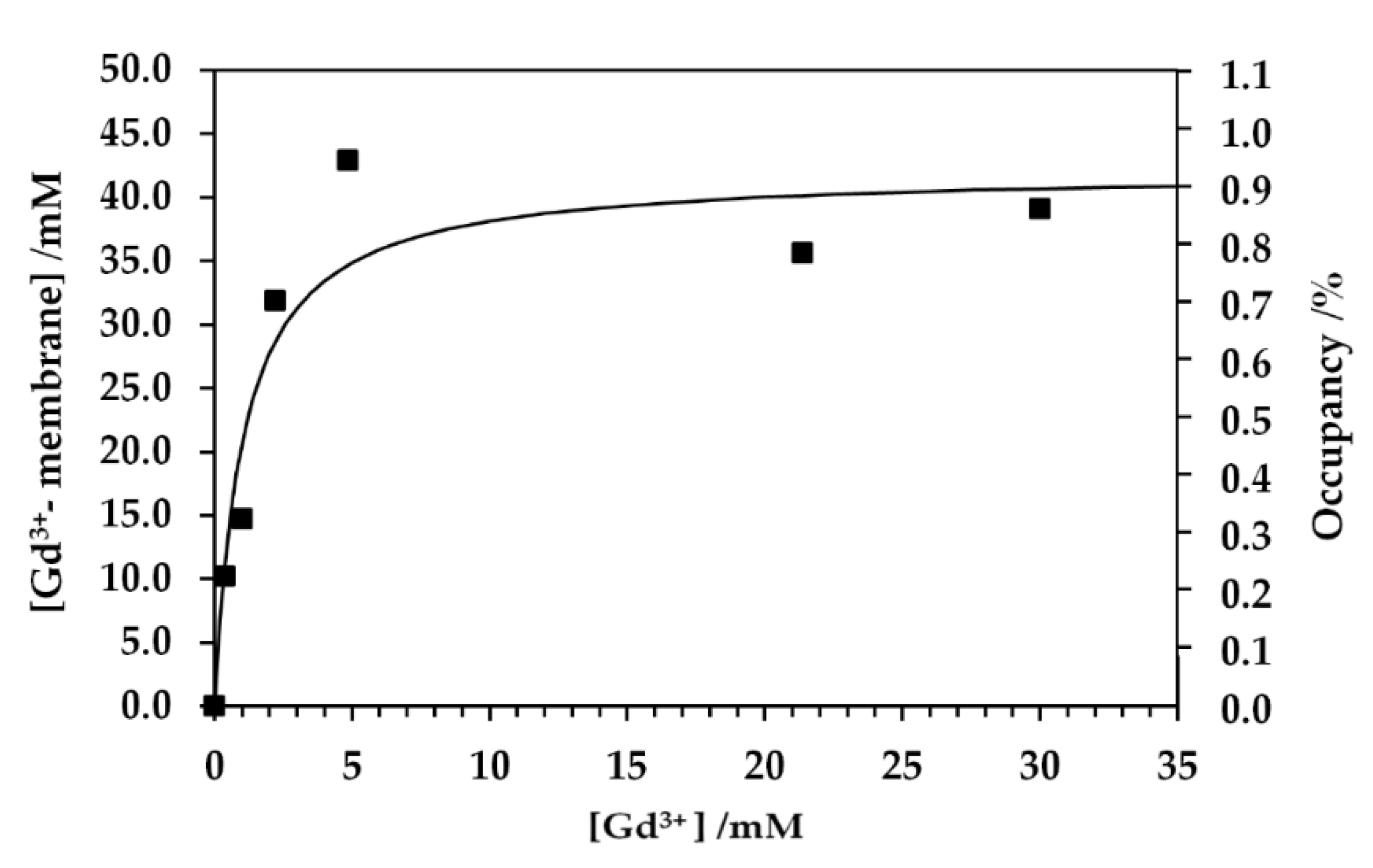{kind=link}
| Agent | [Agent] | Intracellular Solution | Extracellular Solution | ||
|---|---|---|---|---|---|
| (mM) | R1 (s−1) | Amplitude (%) | R1 (s−1) | Amplitude (%) | |
| 0 | 1.5 ± 0.3 | 32 ± 25 | 0.89 ± 0.03 | 68 ± 25 | |
| 15 | 12 ± 0 | 84 ± 1 | 34 ± 1 | 16 ± 1 | |
| 25 | 16 ± 0 | 63 ± 5 | 45 ± 1 | 37 ± 5 | |
| Gd-DOTA | 100 | 16 ± 3 | 44 ± 0 | 121 ± 7 | 56 ± 0 |
| 150 | 34 ± 6 | 32 ± 4 | 182 ± 15 | 68 ± 4 | |
| 200 | 24 ± 7 | 35 ± 3 | 223 ± 30 | 65 ± 3 | |
| 250 | 26 ± 6 | 27 ± 1 | 259 ± 32 | 73 ± 1 | |
| 12 | 1.7 ± 0.2 | 75 ± 3 | 14 ± 3 | 25 ± 3 | |
| 25 | 3.6 ± 2.3 | 65 ± 10 | 36 ± 21 | 35 ± 10 | |
| [Gd(H2O)8]3+ | 100 | 10 ± 1 | 38 ± 3 | 77 ± 2 | 62 ± 3 |
| 149 | 37 ± 11 | 44 ± 16 | 170 ± 26 | 55 ± 16 | |
| 200 | 47 ± 21 | 26 ± 6 | 744 ± 124 | 74 ± 6 | |
| 250 | 25 ± 7 | 25 ± 3 | 965 ± 429 | 75 ± 3 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakol, N.; Egawa, A.; Fujiwara, T. Gadolinium Complexes as Contrast Agent for Cellular NMR Spectroscopy. Int. J. Mol. Sci. 2020, 21, 4042. https://doi.org/10.3390/ijms21114042
Sakol N, Egawa A, Fujiwara T. Gadolinium Complexes as Contrast Agent for Cellular NMR Spectroscopy. International Journal of Molecular Sciences. 2020; 21(11):4042. https://doi.org/10.3390/ijms21114042
Chicago/Turabian StyleSakol, Nat, Ayako Egawa, and Toshimichi Fujiwara. 2020. "Gadolinium Complexes as Contrast Agent for Cellular NMR Spectroscopy" International Journal of Molecular Sciences 21, no. 11: 4042. https://doi.org/10.3390/ijms21114042
APA StyleSakol, N., Egawa, A., & Fujiwara, T. (2020). Gadolinium Complexes as Contrast Agent for Cellular NMR Spectroscopy. International Journal of Molecular Sciences, 21(11), 4042. https://doi.org/10.3390/ijms21114042