A Proline Derivative-Enriched Fraction from Sideroxylon obtusifolium Protects the Hippocampus from Intracerebroventricular Pilocarpine-Induced Injury Associated with Status Epilepticus in Mice


 , , , , , ,
, , , , , ,  ,
,
Abstract
:1. Introduction
2. Results
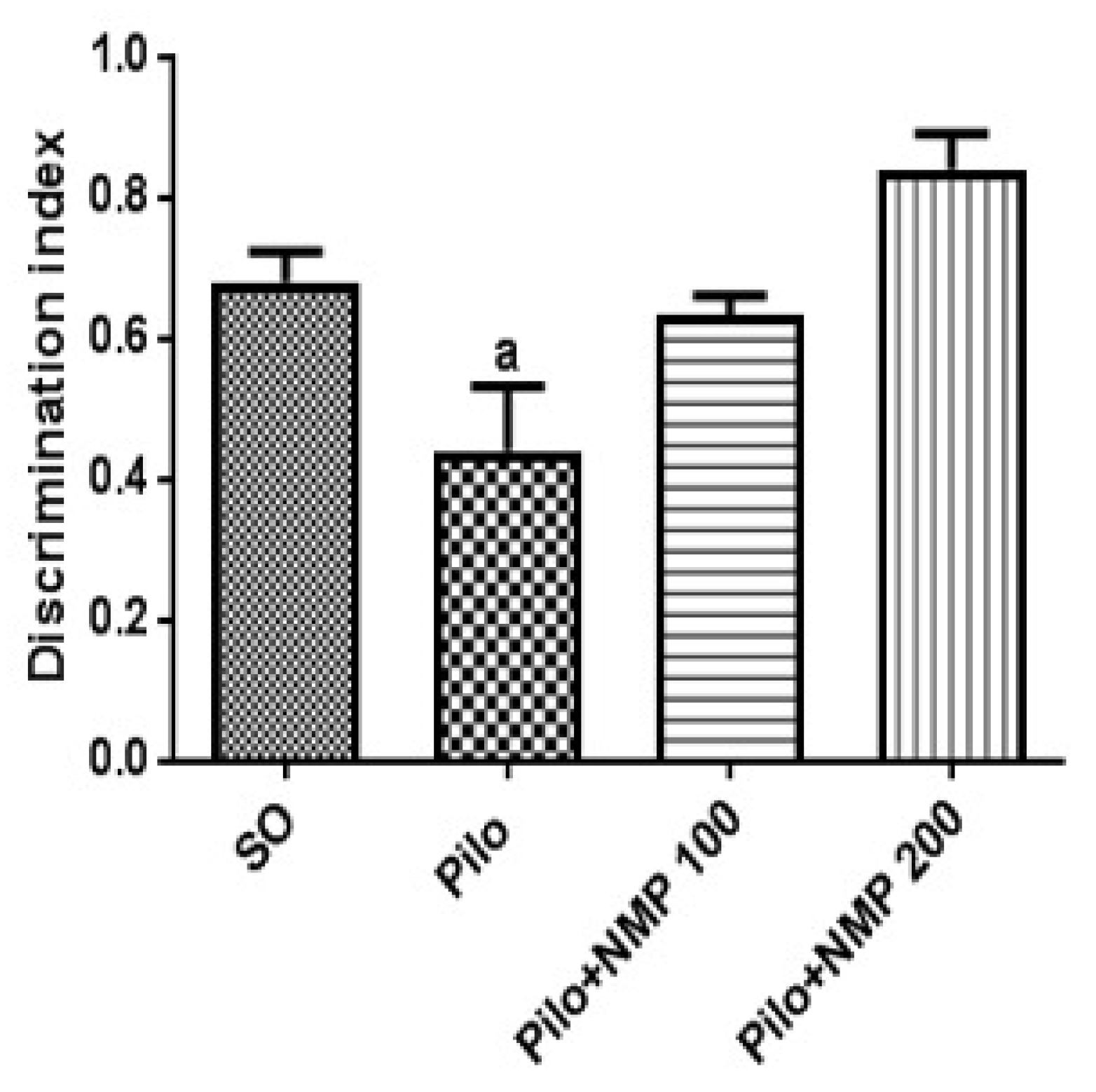
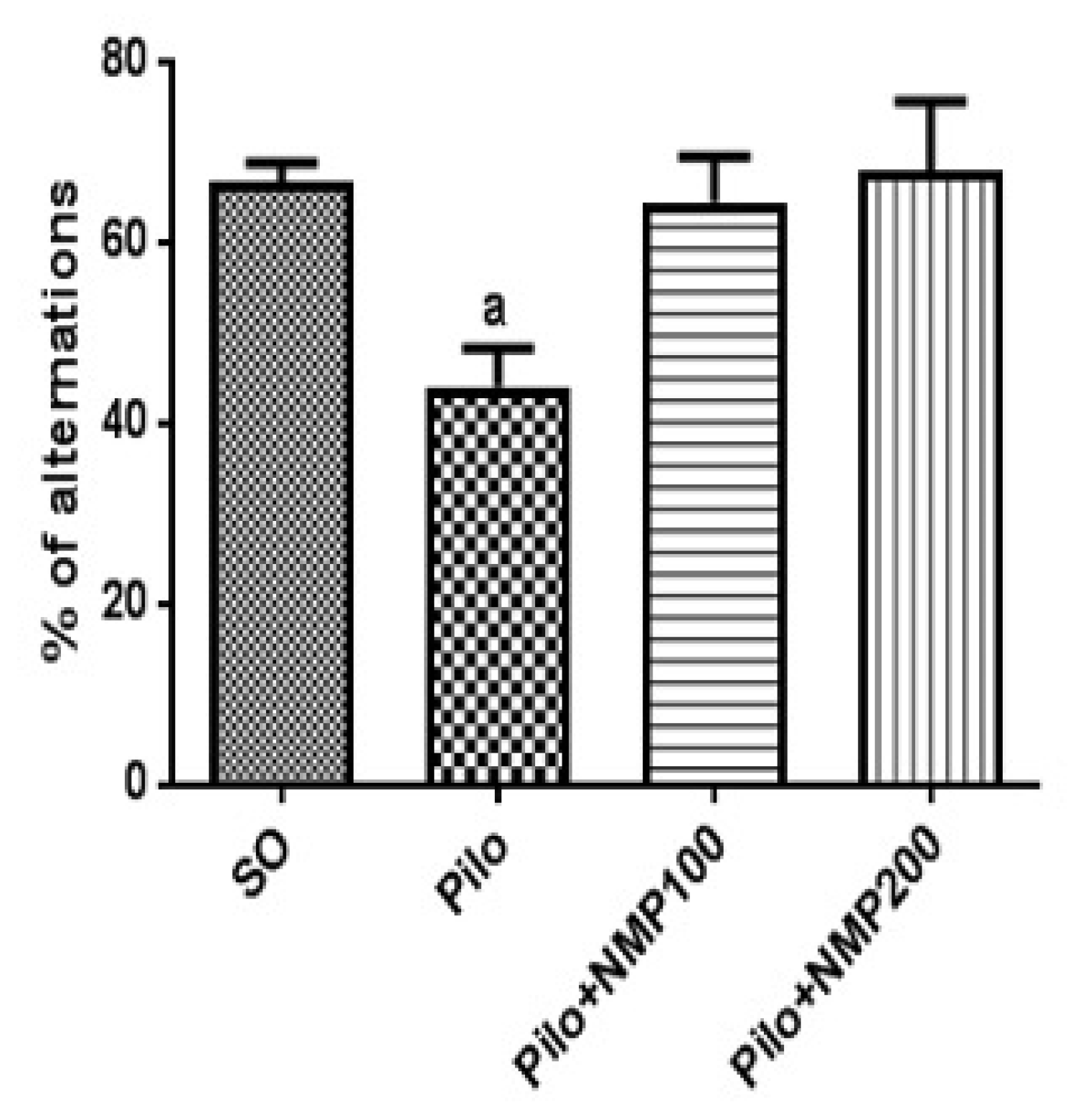
2.1. Behavioral Tests
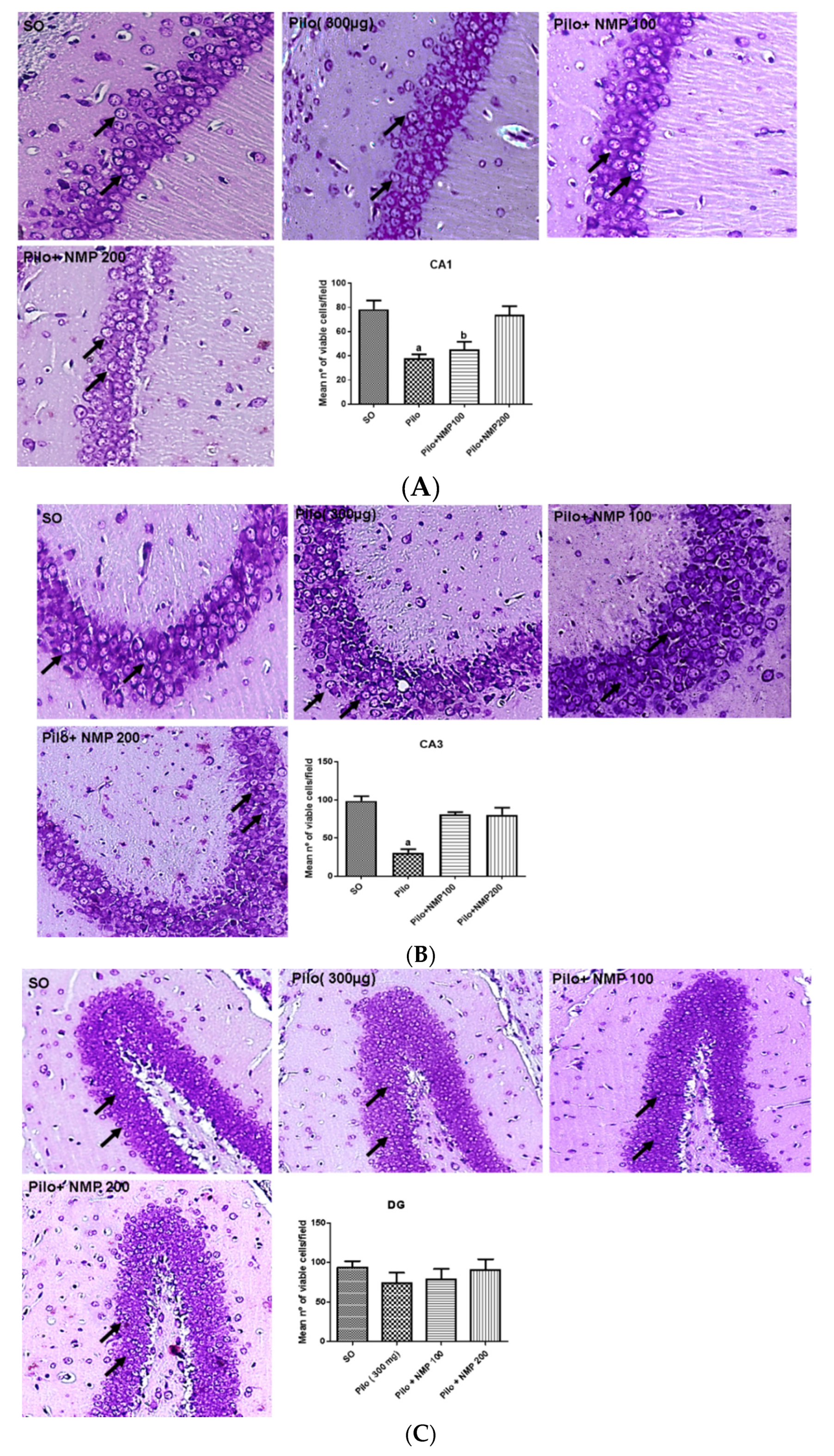
2.2. Nissl Staining and Neuronal Viability
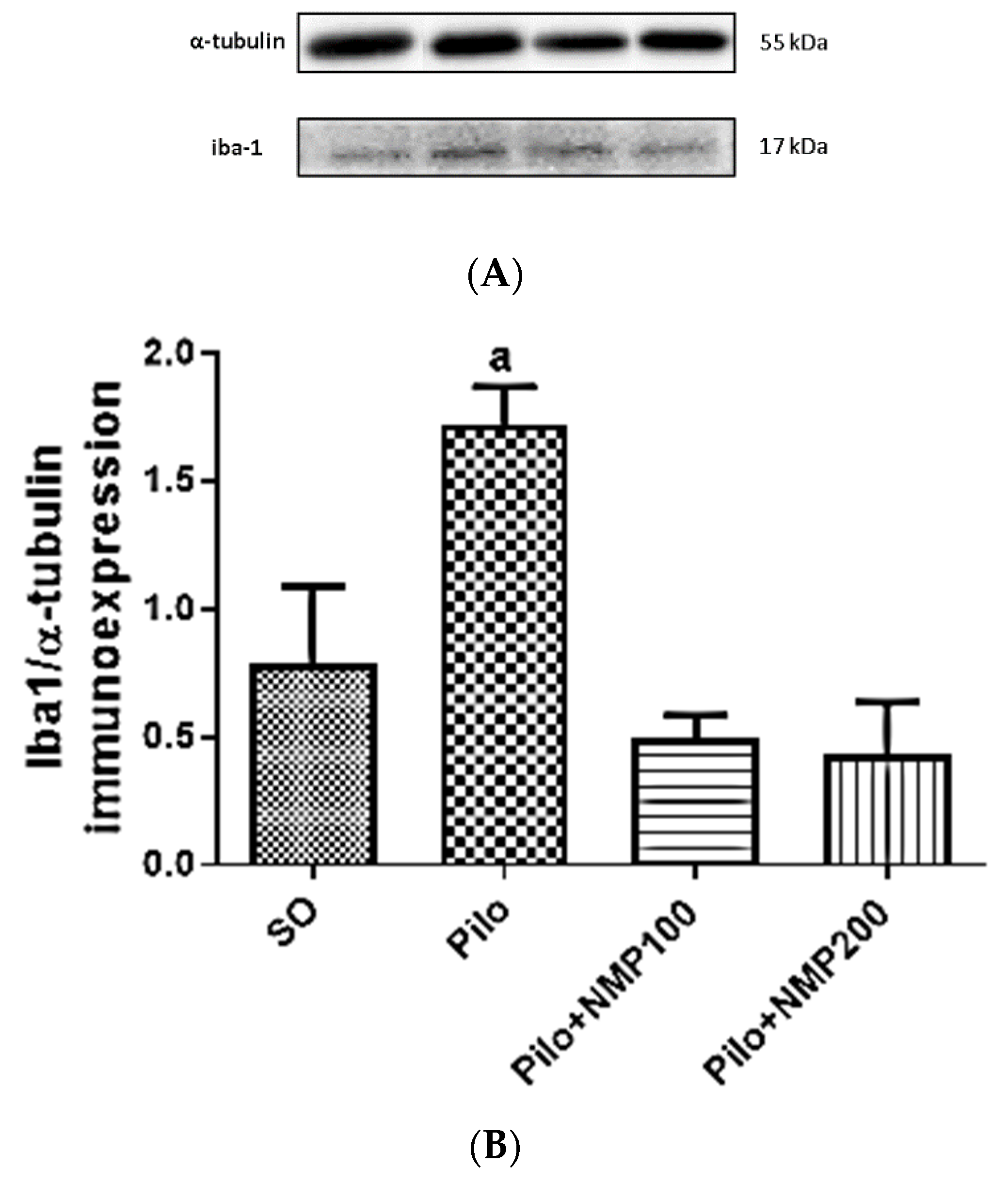
2.3. Ionized Calcium-Binding Adaptor Molecule 1 (Iba-1) Western Blot
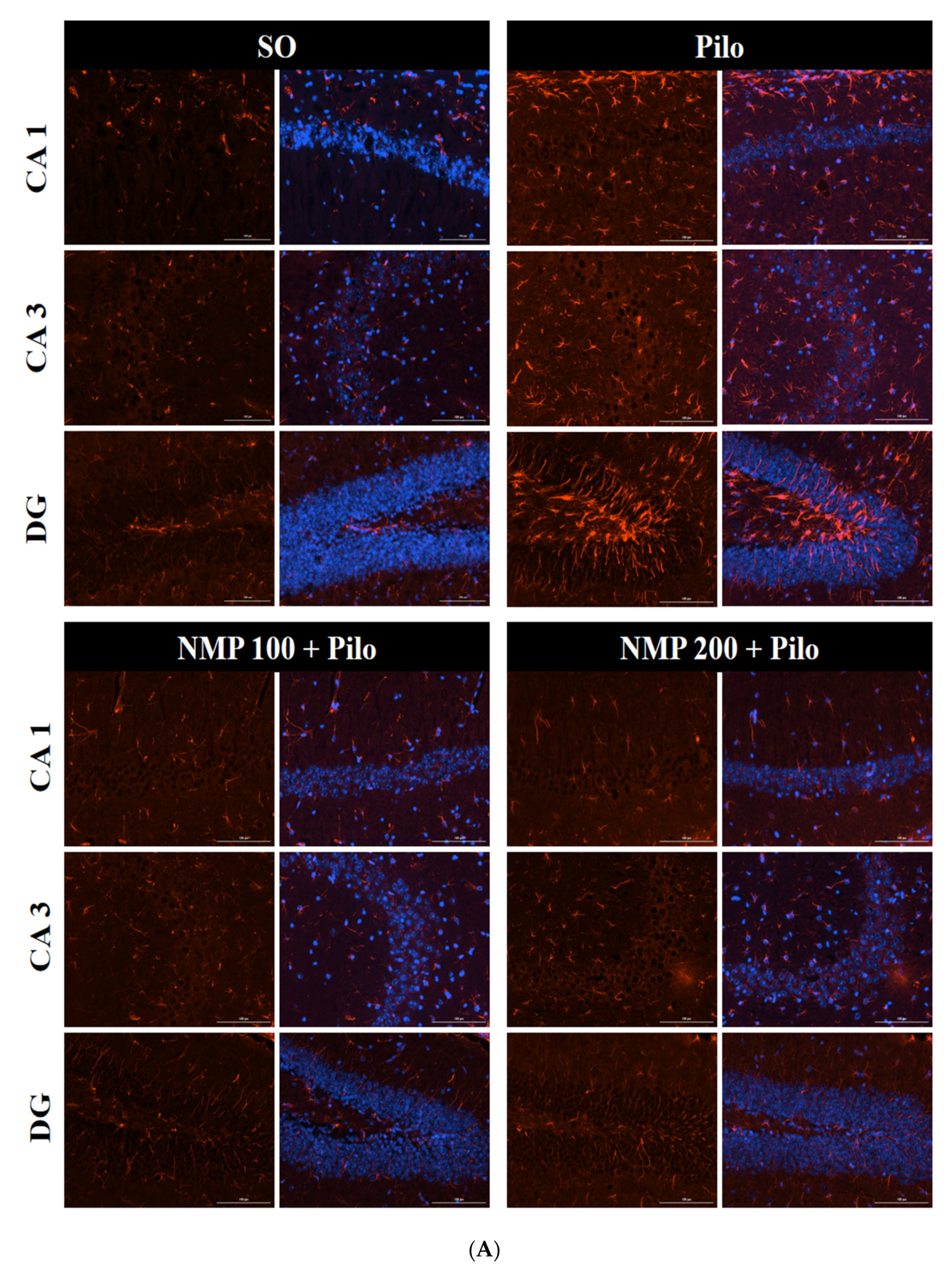
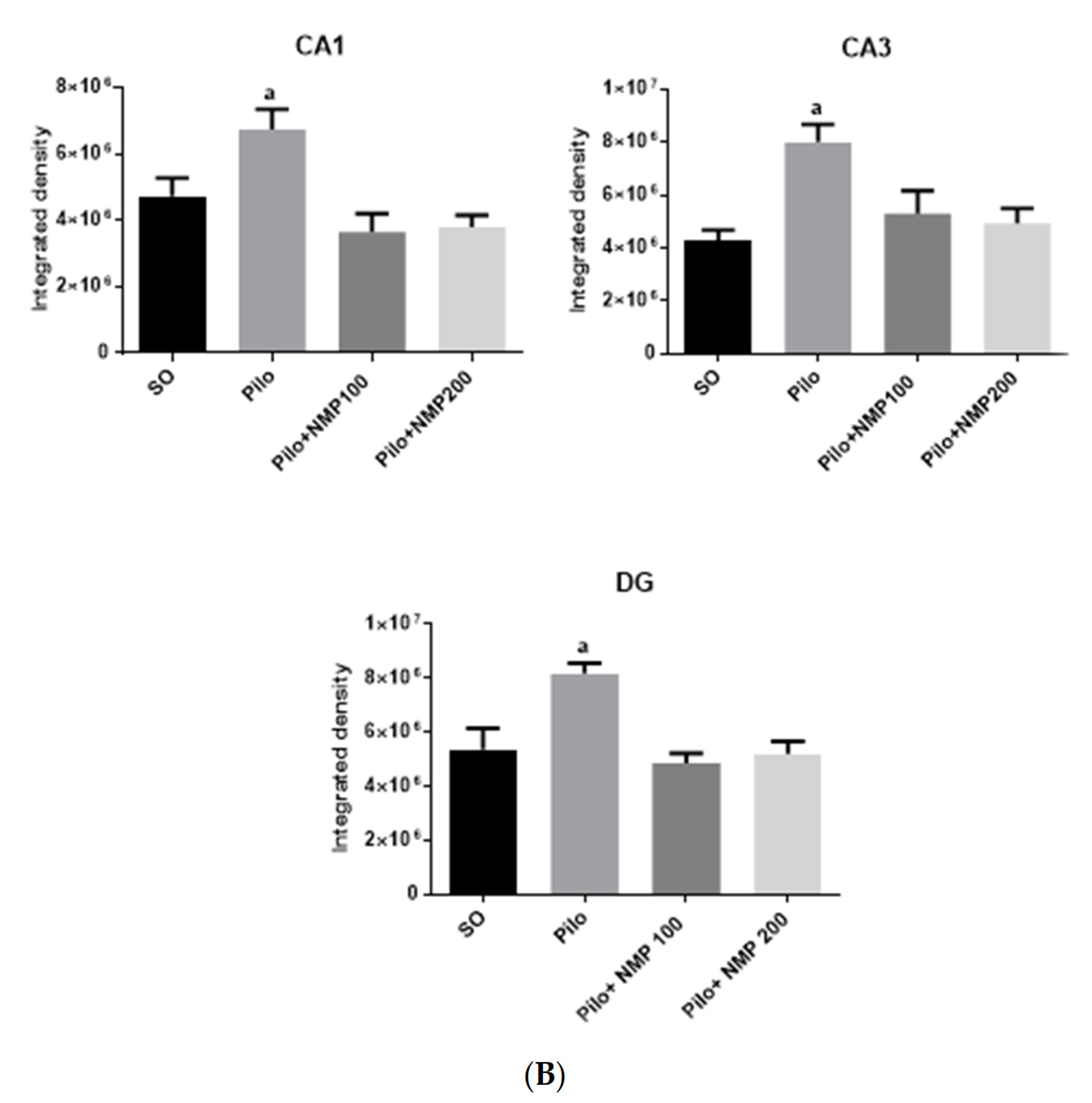
2.4. GFAP Immunofluorescence
2.5. Target Prediction Results for NMP
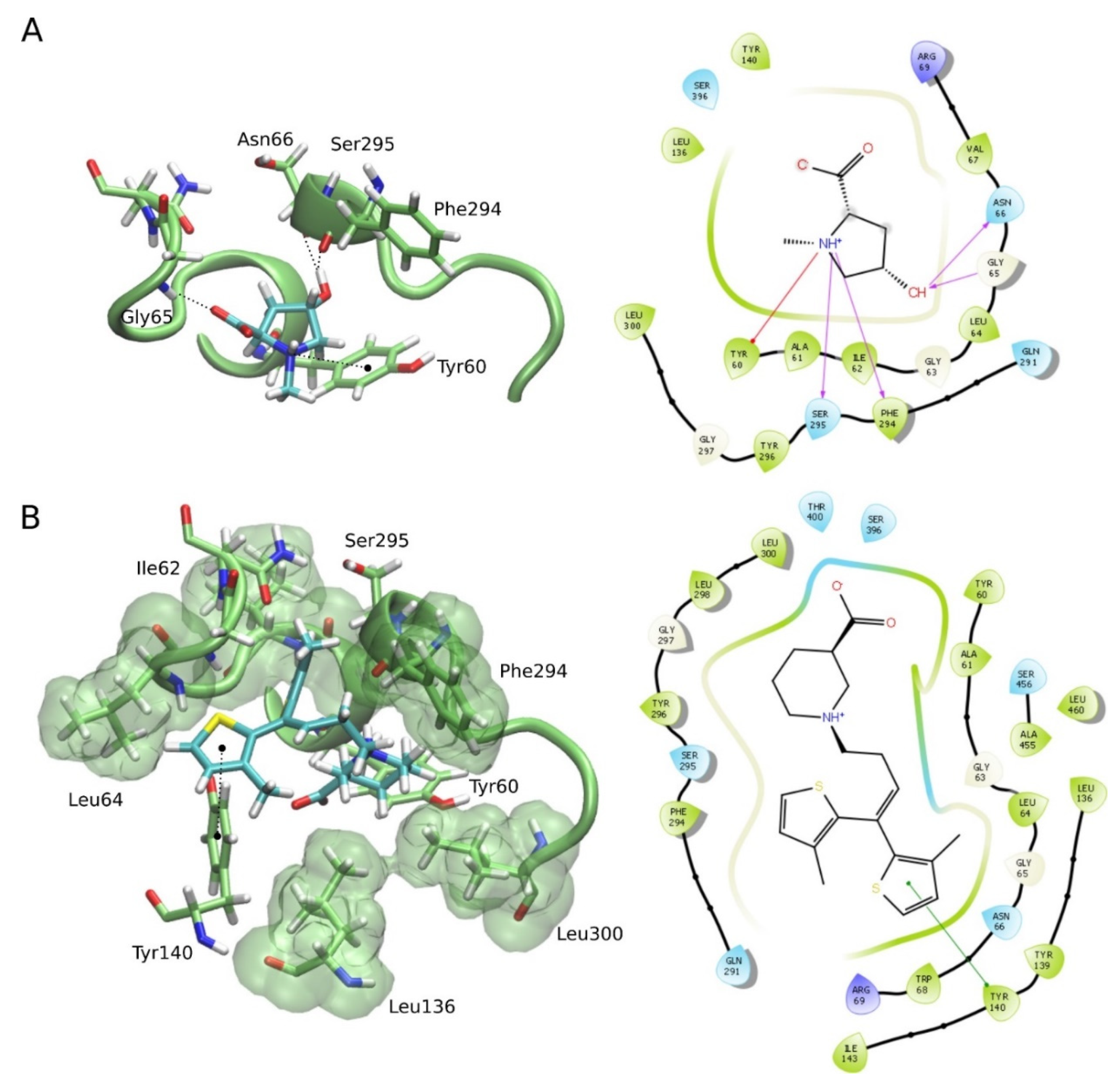
2.6. Molecular Docking for the NMP
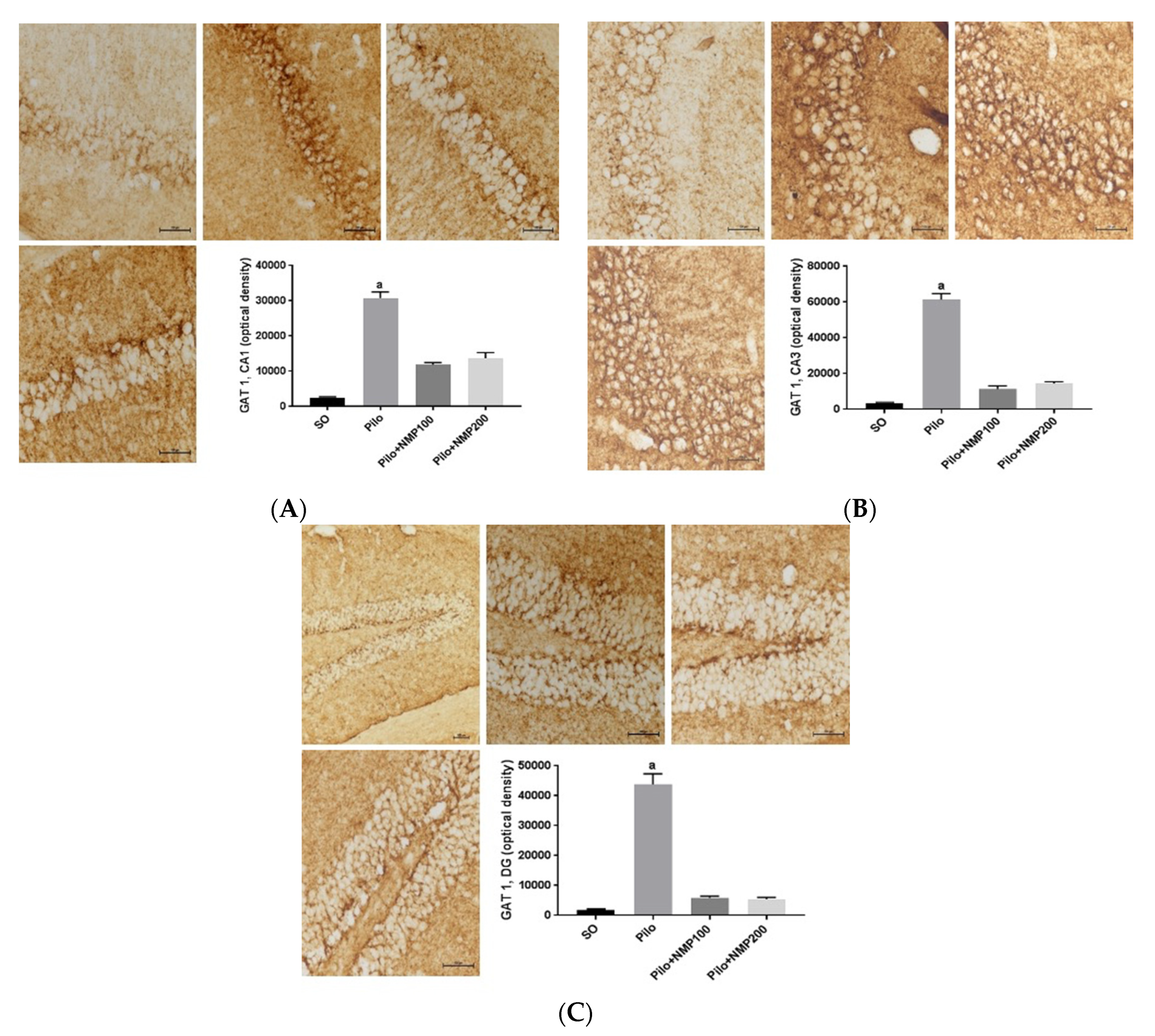
2.7. GAT1 Immunohistochemistry
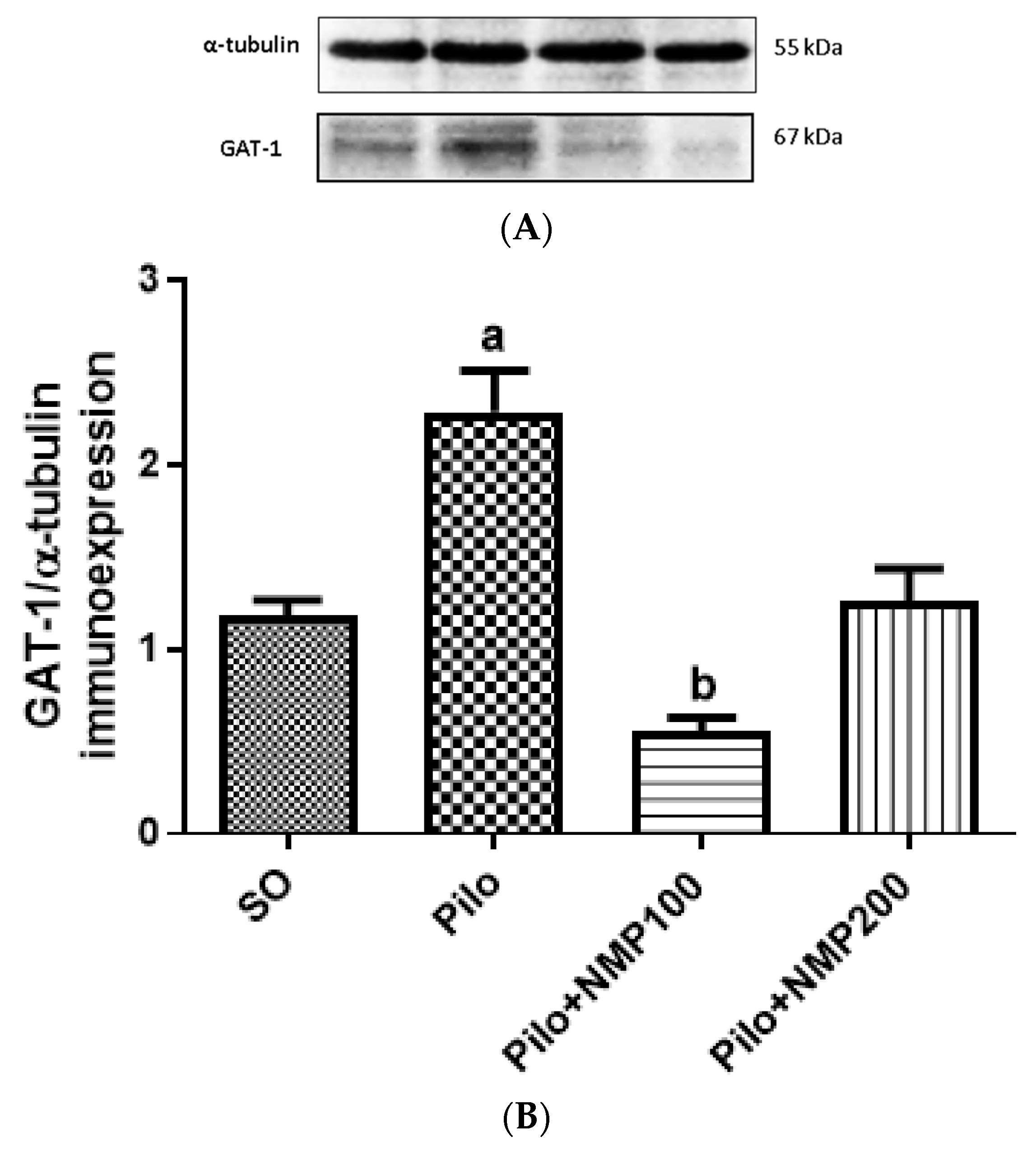
2.8. GAT1 Western Blot
3. Discussion
4. Materials and Methods
4.1. Plant Material Extraction and Purification
4.2. Animals
4.3. Drugs and Chemicals
4.4. Stereotaxic-Guided Injection of Pilo or aCSF in the Left Lateral Ventricle of the Brain, and Treatments
4.5. Behavioral Tests Performed on the 15th Postoperative Day
4.5.1. Novel ORT
4.5.2. Y-Maze Test
4.6. Euthanasia and Tissue Harvesting
4.7. Nissl Staizning
4.8. Drug Discovery Computational Tools
4.8.1. Target Prediction
4.8.2. Homology Modeling and Molecular Docking Calculations: Comparing the Interaction of NMP and Tiagabine with GAT-1
4.9. Immunohistochemistry for GAT-1
4.10. Western Blotting Assays for GAT1 and Iba-1
4.11. Immunofluorescence for GFAP
4.12. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| aCSF | artificial cerebrospinal fluid |
| ANOVA | analysis of variance |
| CA | cornu Ammonis |
| DG | dentate gyrus, dentate fascia |
| GAT1/GAT-1 | GABA transporter type 1, GABA-Na+ symporter 1 |
| GFA | glial fibrillary acidic protein |
| Iba-1 | Ionized calcium-binding adaptor molecule 1 |
| icv | intracerebroventricular route of administration |
| MTLE | mesial temporal lobe epilepsy |
| NMP | N-methyl-(2S,4R)-trans-4-hydroxy-l-proline |
| ORT | novel object recognition test |
| Pilo | pilocarpine |
| PBS | phosphate buffered saline |
References
- Moshé, S.L.; Perucca, E.; Ryvlin, P.; Tomson, T. Epilepsy: New advances. Lancet 2015, 385, 884–898. [Google Scholar] [CrossRef]
- GBD 2015 Neurological Disorders Collaborator Group. Global, regional, and national burden of neurological disorders during 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Neurol. 2017, 16, 877–897. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Cross, J.H.; Elger, C.E.; Engel, J.; Forsgren, L.; French, J.A.; Glynn, M.; et al. ILAE Official Report: A practical clinical definition of epilepsy. Epilepsia 2014, 55, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Téllez-Zenteno, J.F.; Hernández-Ronquillo, L. A review of the epidemiology of temporal lobe epilepsy. Epilepsy Res. Treat. 2012, 2012, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Blümcke, I.; Thom, M.; Aronica, E.; Armstrong, D.D.; Bartolomei, F.; Bernasconi, A.; Bernasconi, N.; Bien, C.G.; Cendes, F.; Coras, R.; et al. International consensus classification of hippocampal sclerosis in temporal lobe epilepsy: A Task Force report from the ILAE Commission on Diagnostic Methods. Epilepsia 2013, 54, 1315–1329. [Google Scholar] [CrossRef] [PubMed]
- Shorvon, S.D. The etiologic classification of epilepsy. Epilepsia 2011, 52, 1052–1057. [Google Scholar] [CrossRef]
- Kwan, P.; Arzimanoglou, A.; Berg, A.T.; Brodie, M.J.; Hauser, W.A.; Mathern, G.; Moshé, S.L.; Perucca, E.; Wiebe, S.; French, J. Definition of drug resistant epilepsy: Consensus proposal by the ad hoc Task Force of the ILAE Commission on Therapeutic Strategies. Epilepsia 2010, 51, 1069–1077. [Google Scholar] [CrossRef]
- Gilioli, I.; Vignoli, A.; Visani, E.; Casazza, M.; Canafoglia, L.; Chiesa, V.; Gardella, E.; La Briola, F.; Panzica, F.; Avanzini, G.; et al. Focal epilepsies in adult patients attending two epilepsy centers: Classification of drug-resistance, assessment of risk factors, and usefulness of “new” antiepileptic drugs. Epilepsia 2012, 53, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Wiebe, S.; Jette, N. Pharmacoresistance and the role of surgery in difficult to treat epilepsy. Nat. Rev. Neurol. 2012, 8, 669–677. [Google Scholar] [CrossRef]
- Beghi, E. Addressing the burden of epilepsy: Many unmet needs. Pharmacol. Res. 2016, 107, 79–84. [Google Scholar] [CrossRef]
- Greenfield, L.J. Molecular mechanisms of antiseizure drug activity at GABAA receptors. Seizure 2013, 22, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scimemi, A. Structure, function, and plasticity of GABA transporters. Front. Cell. Neurosci. 2014, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinet, J.; Vainchtein, I.D.; Spano, C.; Giordano, C.; Bordini, D.; Curia, G.; Dominici, M.; Boddeke, H.W.G.M.; Eggen, B.J.L.; Biagini, G. Microglia are less pro-inflammatory than myeloid infiltrates in the hippocampus of mice exposed to status epilepticus. Glia 2016, 64, 1350–1362. [Google Scholar] [CrossRef]
- Vinet, J.; Costa, A.-M.; Salinas-Navarro, M.; Leo, G.; Moons, L.; Arckens, L.; Biagini, G. A Hydroxypyrone-based inhibitor of metalloproteinase-12 displays neuroprotective properties in both status epilepticus and optic nerve crush animal models. Int. J. Mol. Sci. 2018, 19, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Setkowicz, Z.; Kosonowska, E.; Janeczko, K. Inflammation in the developing rat modulates astroglial reactivity to seizures in the mature brain. J. Anat. 2017, 231, 366–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steward, O.; Torre, E.R.; Tomasulo, R.; Lothman, E. Seizures and the regulation of astroglial gene expression. Epilepsy Res. Suppl. 1992, 7, 197–209. [Google Scholar]
- Steward, O.; Kelley, M.S.; Schauwecker, P.E. Signals that regulate astroglial gene expression: Induction of gfap mrna following seizures or injury is blocked by protein synthesis inhibitors. Exp. Neurol. 1997, 148, 100–109. [Google Scholar] [CrossRef]
- Mula, M.; Sander, J.W. Psychosocial aspects of epilepsy: A wider approach. Br. J. Psychiatry Open 2016, 2, 270–274. [Google Scholar] [CrossRef] [Green Version]
- Helmstaedter, C.; Kockelmann, E. Cognitive outcomes in patients with chronic temporal lobe epilepsy. Epilepsia 2006, 47, 96–98. [Google Scholar] [CrossRef]
- Drinovac, M.; Wagner, H.; Agrawal, N.; Cock, H.R.; Mitchell, A.J.; von Oertzen, T.J. Screening for depression in epilepsy: A model of an enhanced screening tool. Epilepsy Behav. 2015, 44, 67–72. [Google Scholar] [CrossRef]
- Josephson, C.B.; Lowerison, M.; Vallerand, I.; Sajobi, T.T.; Patten, S.; Jette, N.; Wiebe, S. Association of depression and treated depression with epilepsy and seizure outcomes a multicohort analysis. JAMA Neurol. 2017, 74, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Van Elst, L.T.; Woermann, F.G.; Lemieux, L.; Thompson, P.J.; Trimble, M.R. Affective aggression in patients with temporal lobe epilepsy: A quantitative MRI study of the amygdala. Brain 2000, 123, 234–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.; Bankstahl, J.P.; Löscher, W.; Bankstahl, M. Sucrose consumption test reveals pharmacoresistant depression-associated behavior in two mouse models of temporal lobe epilepsy. Exp. Neurol. 2015, 263, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Epps, S.A.; Kahn, A.B.; Holmes, P.V.; Boss-Williams, K.A.; Weiss, J.M.; Weinshenker, D. Antidepressant and anticonvulsant effects of exercise in a rat model of epilepsy and depression comorbidity. Epilepsy Behav. 2013, 29, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brillatz, T.; Lauritano, C.; Jacmin, M.; Khamma, S.; Marcourt, L.; Righi, D.; Romano, G.; Esposito, F.; Ianora, A.; Queiroz, E.F.; et al. Zebrafish-based identification of the antiseizure nucleoside inosine from the marine diatom Skeletonema marinoi. PLoS ONE 2018, 13, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohni, N.; Cordero-Maldonado, M.L.; Maes, J.; Siverio-Mota, D.; Marcourt, L.; Munck, S.; Kamuhabwa, A.R.; Moshi, M.J.; Esguerra, C.V.; de Witte, P.A.M.; et al. Integration of microfractionation, qNMR and Zebrafish screening for the in vivo bioassay-guided isolation and quantitative bioactivity analysis of natural products. PLoS ONE 2013, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Araujo-Neto, V.; Bomfim, R.R.; Oliveira, V.O.B.; Passos, A.M.P.R.; Oliveira, J.P.R.; Lima, C.A.; Mendes, S.S.; Estevam, C.S.; Thomazzi, S.M. Therapeutic benefits of Sideroxylon obtusifolium (Humb. ex Roem. & Schult.) T.D. Penn., Sapotaceae, in experimental models of pain and inflammation. Rev. Bras. Farmacogn. 2010, 20, 933–938. [Google Scholar]
- Aquino, P.E.A.; Magalhães, T.R.; Nicolau, L.A.D.; Leal, L.A.K.M.A.M.; de Aquino, N.C.; dos Santos, S.M.; Neves, K.R.T.; Silveira, E.R.; de Barros Viana, G.S. The anti-inflammatory effects of N-methyl-(2S,4R)-trans-4-hydroxy-l-proline from Sideroxylon obtusifolium are related to its inhibition of TNF-alpha and inflammatory enzymes. Phytomedicine 2016, 24, 14–23. [Google Scholar] [CrossRef]
- Curia, G.; Longo, D.; Biagini, G.; Jones, R.S.G.; Avoli, M. The pilocarpine model of temporal lobe epilepsy. J. Neurosci. Methods 2008, 172, 143–157. [Google Scholar] [CrossRef]
- Aquino, P.E.A.; Paes, L.C.F.; Costa, R.O.; Lima, L.A.R.; Siqueira, E.A.; Carvalho, M.A.J.; Cavalcante, T.M.B.; Silveira, E.R.; Viana, G.S.B. N-methyl-trans-4-hydroxy-l-proline isolated from Sideroxylon obtusifolium leaves has antioxidant and anticonvulsivant activity. In Proceedings of the 2nd International Neuroscience Symposium, Fortaleza, Brazil, 18 December 2018. [Google Scholar]
- Medina-Ceja, L.; Pardo-Peña, K.; Ventura-Mejía, C. Evaluation of behavioral parameters and mortality in a model of temporal lobe epilepsy induced by intracerebroventricular pilocarpine administration. Neuroreport 2014, 25, 875–879. [Google Scholar] [CrossRef]
- Devinsky, O.; Vazquez, B. Behavioral Changes Associated with Epilepsy. Neurol. Clin. 1993, 11, 127–149. [Google Scholar] [CrossRef]
- Cornaggia, C.M.; Beghi, M.; Provenzi, M.; Beghi, E. Correlation between Cognition and Behavior in Epilepsy. Epilepsia 2006, 47, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Lueptow, L.M. Novel object recognition test for the investigation of learning and memory in mice. J. Vis. Exp. 2017, 126, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kraeuter, A.; Guest, P.C.; Sarnyai, Z. The Y-Maze for Assessment of Spatial Working and Reference Memory in Mice. In Pre-Clinical Models: Techniques and Protocols, Methods in Molecular BIOLOGY; Guest, P.C., Ed.; Humana Press: New York, NY, USA, 2019; Volume 1916, pp. 105–111. [Google Scholar]
- Sarhan, S.; Seiler, N. Proline and proline derivatives as anticonvulsants. Gen. Pharmac. 1989, 20, 53–60. [Google Scholar] [CrossRef]
- De, A.; Furtado, M.; Braga, G.K.; Oliveira, J.A.C.; Del Vecchio, F.; Garcia-Cairasco, N. Behavioral, morphologic, and electroencephalographic evaluation of seizures induced by intrahippocampal microinjection of pilocarpine. Epilepsia 2002, 43, 37–39. [Google Scholar]
- Furtado, M.A.; Castro, O.W.; Del Vecchio, F.; de Oliveira, J.A.C.; Garcia-Cairasco, N. Study of spontaneous recurrent seizures and morphological alterations after status epilepticus induced by intrahippocampal injection of pilocarpine. Epilepsy Behav. 2011, 20, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, B.; Wheal, H.V. A comparative histological and electrophysiological study of some neurotoxins in the rat hippocampus. J. Comp. Neurol. 1982, 211, 105–114. [Google Scholar] [CrossRef]
- Victor Nadler, J.; Cuthbertson, G.J. Kainic acid neurotoxicity toward hippocampal formation: Dependence on specific excitatory pathways. Brain Res. 1980, 195, 47–56. [Google Scholar] [CrossRef]
- Binder, D.K.; Carson, M.J. Glial cells as primary therapeutic targets for epilepsy. Neurochem. Int. 2013, 63, 635–637. [Google Scholar] [CrossRef]
- Diaz Verdugo, C.; Myren-Svelstad, S.; Aydin, E.; Van Hoeymissen, E.; Deneubourg, C.; Vanderhaeghe, S.; Vancraeynest, J.; Pelgrims, R.; IlyasCosacak, M.; Muto, A.; et al. Glia-neuron interactions underlie state transitions to generalized seizures. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Liu, Z.R.; Chen, J.; Zhang, S.J.; Quan, Q.Y.; Huang, Y.G.; Jiang, W. Roles of astrocytes and microglia in seizure-induced aberrant neurogenesis in the hippocampus of adult rats. J. Neurosci. Res. 2010, 88, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Schartz, N.D.; Herr, S.A.; Madsen, L.; Butts, S.J.; Torres, C.; Mendez, L.B.; Brewster, A.L. Spatiotemporal profile of Map2 and microglial changes in the hippocampal CA1 region following pilocarpine-induced status epilepticus. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiragi, T.; Ikegaya, Y.; Koyama, R.; Hiragi, T.; Ikegaya, Y.; Koyama, R. Microglia after seizures and in epilepsy. Cells 2018, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Liao, Y.; Morgan, S.; Mathur, R.; Feustel, P.; Mazurkiewicz, J.; Qian, J.; Chang, J.; Mathern, G.W.; Adamo, M.A.; et al. Noninflammatory changes of microglia are sufficient to cause epilepsy. Cell Rep. 2018, 22, 2080–2093. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, L.A.; Wang, L.; Ribak, C.E. Rapid astrocyte and microglial activation following pilocarpine-induced seizures in rats. Epilepsia 2008, 49, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt-Johnson, S.K.; Herr, S.A.; Brewster, A.L. Status epilepticus triggers time-dependent alterations in microglia abundance and morphological phenotypes in the hippocampus. Front. Neurol. 2017, 8, 700. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Sun, Q.; Fan, J.; Jiang, Y.; Yang, W.; Cui, Y.; Yu, Z.; Jiang, H.; Li, B. role of astrocytes in post-traumatic epilepsy. Front. Neurol. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Steward, O.; Torre, E.R.; Tomasulo, R.; Lothman, E. Neuronal activity up-regulates astroglial gene expression. Proc. Natl. Acad. Sci. USA 1991, 88, 6819–6823. [Google Scholar] [CrossRef] [Green Version]
- Arisi, G.M.; Ruch, M.; Foresti, M.L.; Mukherjee, S.; Ribak, C.E.; Shapiro, L.A. Astrocyte alterations in the hippocampus following pilocarpine-induced seizures in aged rats. Aging Dis. 2011, 2, 294–300. [Google Scholar]
- Coulter, D.A.; Steinhäuser, C. Role of astrocytes in epilepsy. In Cold Spring Harbor Perspectives in Medicine; Holmes, G.L., Noebels, J.L., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2015; Volume 5, pp. 1–12. [Google Scholar]
- Robel, S.; Buckingham, S.C.; Boni, J.L.; Campbell, S.L.; Danbolt, N.C.; Riedemann, T.; Sutor, B.; Sontheimer, H. Reactive astrogliosis causes the development of spontaneous seizures. J. Neurosci. 2015, 35, 3330–3345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, D.K.; Steinhäuser, C. Role of astrocyte dysfunction in epilepsy. In The Curated Reference Collection in Neuroscience and Biobehavioral Psychology; Stein, J., Ed.; Elsevier: Amsterdan, Holland, 2017; pp. 412–417. [Google Scholar]
- Challal, S.; Bohni, N.; Buenafe, O.E.; Esguerra, C.V.; De Witte, P.A.M.; Wolfender, J.L.; Crawford, A.D. Zebrafish bioassay-guided microfractionation for the rapid in vivo identification of pharmacologically active natural products. Chimia (Aarau) 2012, 66, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Schachter, S.C. Pharmacology and clinical experience with tiagabine. Expert Opin. Pharmacother. 2001, 2, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.F.; Esplin, B.; Čapek, R. Inhibitory nature of tiagabine-augmented GABA (A) receptor-mediated depolarizing responses in hippocampal pyramidal cells. J. Neurophysiol. 1999, 81, 1192–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, E.C.; Wu, K.C.; Kao, F.C.; Wu, S.N. Effects of midazolam on ion currents and membrane potential in differentiated motor neuron-like NSC-34 and NG108-15 cells. Eur. J. Pharmacol. 2014, 724, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Vonderlin, N.; Fischer, F.; Zitron, E.; Seyler, C.; Scherer, D.; Thomas, D.; Katus, H.A.; Scholz, E.P. Anesthetic drug midazolam inhibits cardiac human ether-à-go-go-related gene channels: Mode of action. Drug Des. Devel. Ther. 2015, 9, 867–877. [Google Scholar] [PubMed] [Green Version]
- Kim, D.-S.; Kim, J.-E.; Kwak, S.-E.; Choi, H.-C.; Song, H.-K.; Kimg, Y.-I.; Choi, S.-Y.; Kang, T.-C. Up-regulated astroglial TWIK-related acid-sensitive K+ channel-1 (TASK-1) in the hippocampus of seizure-sensitive gerbils: A target of anti-epileptic drugs. Brain Res. 2007, 1185, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.-Y.; Huang, H.-Y.I.; Wu, S.-N.; Huang, C.-W. Depressive effectiveness of vigabatrin (y-Vinyl-GABA), an antiepileptic drug, in intermediate-conductance calcium-sctivated potassium channels in human glioma cells. unpublished. [CrossRef]
- Tescarollo, F.C.; Rombo, D.M.; De Liberto, L.K.; Fedele, D.E.; Alharfoush, E.; Tomé, Â.R.; Cunha, R.A.; Sebastião, A.M.; Boison, D. Role of adenosine in epilepsy and seizures. J. Caffeine Adenosine Res. 2020, 00, 1–16. [Google Scholar] [CrossRef]
- Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use Oflaboratory Animals, 8th ed.; Garber, J.C., Ed.; The National Academies Press: Washington, DC, USA, 2011; pp. 1–208. [Google Scholar]
- Murphy, N.P.; Maidment, N.T. Orphanin FQ/Nociceptin modulation of mesolimbic dopamine transmission determined by microdialysis. J. Neurochem. 2002, 73, 179–186. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: San Diego, CA, USA, 2001; pp. 1–264. [Google Scholar]
- Ferry, B.; Gervasoni, D.; Vogt, C. Stereotaxic Neurosurgery in Laboratory Rodent—Handbook on Best Practices, 1st ed.; Springer: Paris, France, 2014; pp. 1–198. [Google Scholar]
- Cavalheiro, E.A.; Santos, N.F.; Priel, M.R. The pilocarpine model of epilepsy in mice. Epilepsia 1996, 37, 1015–1019. [Google Scholar] [CrossRef]
- Dix, S.L.; Aggleton, J.P. Extending the spontaneous preference test of recognition: Evidence of object-location and object-context recognition. Behav. Brain Res. 1999, 99, 191–200. [Google Scholar] [CrossRef]
- Sarter, M.; Bodewitz, G.; Stephens, D.N. Attenuation of scopolamine-induced impairment of spontaneous alternation behaviour by antagonist but not inverse agonist and agonist β-carbolines. Psychopharmacology (Berl.) 1988, 94, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Scorza, F.A.; Arida, R.M.; Cysneiros, R.M.; Scorza, C.A.; Albuquerque, M.; de Cavalheiro, E.A. Estudo qualitativo da formação hipocampal de animais hipertensos com epilepsia. Arq. Neuropsiquiatr. 2005, 63, 283–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A.; Martin, M.J.; O’Donovan, C.; Magrane, M.; Alpi, E.; Antunes, R.; Bely, B.; Bingley, M.; Bonilla, C.; Britto, R.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Pagadala, N.S.; Syed, K.; Tuszynski, J. Software for molecular docking: A review. Biophys. Rev. 2017, 9, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-M.; Raine, L.; Fanger, H. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: A comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem. 1981, 29, 577–580. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Uniprot ID | Target Class | Probability | Known Actives (3D/2D) |
|---|---|---|---|---|
| GABA transporter 1 | P30531 | Electrochemical transporter | 0.03 | 5/4 |
| GABA A receptor lpha-3/beta-2/gamma-2 | P34903 P47870 P18507 | Ligand-gated ion channel | 0.03 | 3/0 |
| GABA A receptor alpha-2/beta-2/gamma-2 | P47869 P47870 P18507 | Ligand-gated ion channel | 0.03 | 4/0 |
| Renin | P00797 | Protease | 0.03 | 0/13 |
| Egl nine homolog 1 | Q9GZT9 | Oxidoreductase | 0.02 | 1/0 |
| Adenosine A3 receptor | P0DMS8 | Family A G protein-coupled receptor | 0.02 | 2/3 |
| GABA-B receptor | O75899 Q9UBS5 | Family C G protein-coupled receptor | 0 | 8/0 |
| Thrombin | P00734 | Protease | 0 | 0/8 |
| Leukotriene A4 hydrolase | P09960 | Protease | 0 | 0/4 |
| GABA receptor rho-1 subunit | P24046 | Ligand-gated ion channel | 0 | 4/0 |
| Phospholipase A2 group IIA | P14555 | Enzyme | 0 | 0/6 |
| Phospholipase A2 group V | P39877 | Enzyme | 0 | 0/3 |
| Integrin alpha-IIb/beta-3 | P08514 P05106 | Membrane receptor | 0 | 0/11 |
| Leucine aminopeptidase | P28838 | Protease | 0 | 0/5 |
| Bile acid receptor FXR | Q96RI1 | Nuclear receptor | 0 | 0/1 |
| Neprilysin | P08473 | Protease | 0 | 0/23 |
| Voltage-gated calcium channel alpha2/delta subunit 1 (by homology) | P54289 | Calcium channel auxiliary subunit alpha2delta family | 0 | 10/0 |
| Metastin receptor | Q969F8 | Family A G protein-coupled receptor | 0 | 0/2 |
| 11-beta-hydroxysteroid dehydrogenase 1 | P28845 | Enzyme | 0 | 0/9 |
| Nitric-oxide synthase, brain | P29475 | Enzyme | 0 | 0/9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Aquino, P.E.A.; Rabelo Bezerra, J.; de Souza Nascimento, T.; Tavares, J.; Rosal Lustosa, Í.; Chaves Filho, A.J.M.; Mottin, M.; Macêdo Gaspar, D.; Andrade, G.M.d.; Tavares Neves, K.R.; et al. A Proline Derivative-Enriched Fraction from Sideroxylon obtusifolium Protects the Hippocampus from Intracerebroventricular Pilocarpine-Induced Injury Associated with Status Epilepticus in Mice. Int. J. Mol. Sci. 2020, 21, 4188. https://doi.org/10.3390/ijms21114188
de Aquino PEA, Rabelo Bezerra J, de Souza Nascimento T, Tavares J, Rosal Lustosa Í, Chaves Filho AJM, Mottin M, Macêdo Gaspar D, Andrade GMd, Tavares Neves KR, et al. A Proline Derivative-Enriched Fraction from Sideroxylon obtusifolium Protects the Hippocampus from Intracerebroventricular Pilocarpine-Induced Injury Associated with Status Epilepticus in Mice. International Journal of Molecular Sciences. 2020; 21(11):4188. https://doi.org/10.3390/ijms21114188
Chicago/Turabian Stylede Aquino, Pedro Everson Alexandre, Jéssica Rabelo Bezerra, Tyciane de Souza Nascimento, Juliete Tavares, Ítalo Rosal Lustosa, Adriano José Maia Chaves Filho, Melina Mottin, Danielle Macêdo Gaspar, Geanne Matos de Andrade, Kelly Rose Tavares Neves, and et al. 2020. "A Proline Derivative-Enriched Fraction from Sideroxylon obtusifolium Protects the Hippocampus from Intracerebroventricular Pilocarpine-Induced Injury Associated with Status Epilepticus in Mice" International Journal of Molecular Sciences 21, no. 11: 4188. https://doi.org/10.3390/ijms21114188
APA Stylede Aquino, P. E. A., Rabelo Bezerra, J., de Souza Nascimento, T., Tavares, J., Rosal Lustosa, Í., Chaves Filho, A. J. M., Mottin, M., Macêdo Gaspar, D., Andrade, G. M. d., Tavares Neves, K. R., Biagini, G., Silveira, E. R., & de Barros Viana, G. S. (2020). A Proline Derivative-Enriched Fraction from Sideroxylon obtusifolium Protects the Hippocampus from Intracerebroventricular Pilocarpine-Induced Injury Associated with Status Epilepticus in Mice. International Journal of Molecular Sciences, 21(11), 4188. https://doi.org/10.3390/ijms21114188