Novel Therapies Boosting T Cell Immunity in Epstein Barr Virus-Associated Nasopharyngeal Carcinoma




Abstract
:1. Nasopharyngeal Carcinoma
2. NPC Epidemiology
3. NPC Aetiology
3.1. Genetic Factors
3.2. Environmental Factors
3.3. Viral Factors: Epstein-Barr Virus
4. NPC Classification
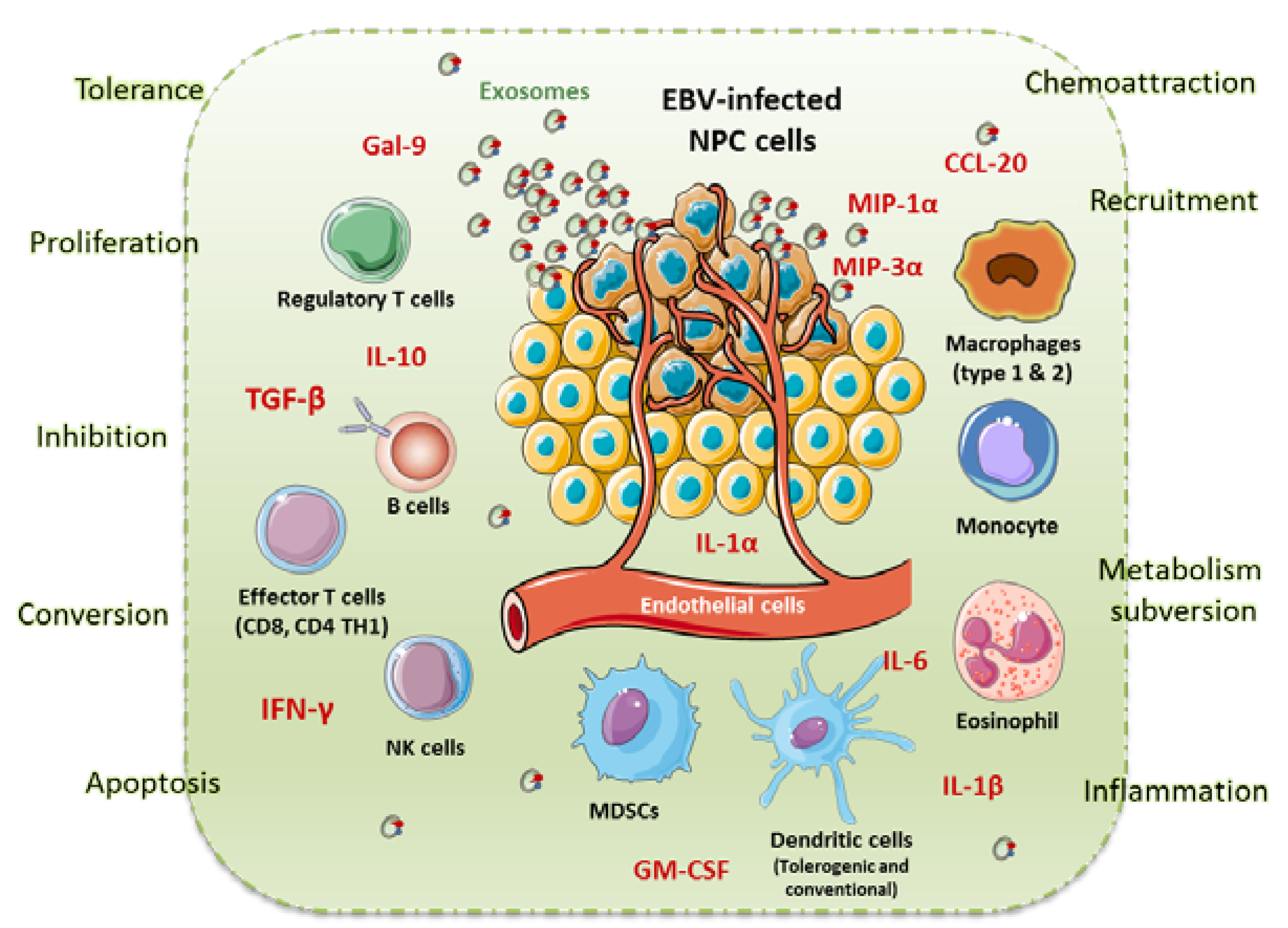
5. NPC Microenvironment
6. NPC Conventional Treatments
7. NPC Novel Therapies
7.1. Targeted Therapies
7.2. Immunotherapies
7.3. EBV-Based Strategies
7.4. Cell Therapy
7.5. Immunotherapy Targeting Checkpoint Inhibitors
7.6. Photodynamic Therapy
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jain, A.; Chia, W.K.; Toh, H.C. Immunotherapy for nasopharyngeal cancer-a review. Chin. Clin. Oncol. 2016, 5, 22. [Google Scholar] [CrossRef]
- Chang, E.T.; Adami, H.-O. The enigmatic epidemiology of nasopharyngeal carcinoma. Cancer Epidemiol. Prev. Biomark. 2006, 15, 1765–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AK, E.-N.; JKC, C.; JR, G.; Takata, T.; Slootweg, P. WHO Classification of Head and Neck Tumours—IARC. Available online: https://www.iarc.fr/fr/news-events/who-classification-of-head-and-neck-tumours-2/ (accessed on 13 April 2020).
- Vokes, E.E.; Liebowitz, D.N.; Weichselbaum, R.R. Nasopharyngeal carcinoma. Lancet Lond. Engl. 1997, 350, 1087–1091. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, J.-O.; Huang, Q.-H.; Li, Y.-H. Trends in the survival of patients with nasopharyngeal carcinoma between 1976 and 2005 in Sihui, China: A population-based study. Chin. J. Cancer 2013, 32, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.-M.; Simons, M.J.; Qian, C.-N. The prevalence and prevention of nasopharyngeal carcinoma in China. Chin. J. Cancer 2011, 30, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkin, D.-M.; Whelan, S.-L.; Ferlay, J.; Raymond, L.; Young, J. Cancer incidence in five continents. Volume VII. IARC Sci. Publ 1997, i–xxxiv, 1–1240. [Google Scholar]
- Yu, M.C.; Yuan, J.-M. Epidemiology of nasopharyngeal carcinoma. Semin. Cancer Biol. 2002, 12, 421–429. [Google Scholar] [CrossRef]
- Zeng, Y.-X.; Jia, W.-H. Familial nasopharyngeal carcinoma. Semin. Cancer Biol. 2002, 12, 443–450. [Google Scholar] [CrossRef]
- Lu, S.J.; Day, N.E.; Degos, L.; Lepage, V.; Wang, P.C.; Chan, S.H.; Simons, M.; McKnight, B.; Easton, D.; Zeng, Y. Linkage of a nasopharyngeal carcinoma susceptibility locus to the HLA region. Nature 1990, 346, 470–471. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.P.; Day, N.E.; Li, D.R.; Luben, R.N.; Cai, K.L.; Ou-Yang, T.; Li, B.; Lu, X.Z.; Ponder, B. Further evidence for an HLA-related recessive mutation in nasopharyngeal carcinoma among the Chinese. Br. J. Cancer 2005, 92, 967–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, W.; Zeng, Z.Y.; Xia, J.H.; Xia, K.; Shen, S.R.; Li, X.L.; Hu, D.X.; Tan, C.; Xiang, J.J.; Zhou, J.; et al. A susceptibility locus at chromosome 3p21 linked to familial nasopharyngeal carcinoma. Cancer Res. 2004, 64, 1972–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, B.-J.; Huang, W.; Shugart, Y.Y.; Lee, M.K.; Zhang, F.; Xia, J.-C.; Wang, H.-Y.; Huang, T.-B.; Jian, S.-W.; Huang, P.; et al. Genome-wide scan for familial nasopharyngeal carcinoma reveals evidence of linkage to chromosome 4. Nat. Genet. 2002, 31, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Zhang, H.; Zhai, Y.; Wang, Z.; Li, P.; Yu, L.; Xia, X.; Zhang, Y.; Zeng, Y.; et al. Common variations in TERT-CLPTM1L locus are reproducibly associated with the risk of nasopharyngeal carcinoma in Chinese populations. Oncotarget 2016, 7, 759–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Z.; Xiao, J.-Y.; Guo, M.-H. [Telomere shortening in the pathogenesis of nasopharyngeal carcinoma]. Acad. J. First Med. Coll. PLA 2002, 22, 329–330. [Google Scholar]
- Du, C.W.; Wen, B.G.; Li, D.R.; Lin, Y.C.; Zheng, Y.W.; Chen, L.; Chen, J.Y.; Lin, W.; Wu, M.Y. Latent membrane protein-1 of Epstein - Barr virus increases sensitivity to arsenic trioxide-induced apoptosis in nasopharyngeal carcinoma cell. Exp. Oncol. 2005, 27, 267–272. [Google Scholar] [PubMed]
- James, M.A.; Vikis, H.G.; Tate, E.; Rymaszewski, A.L.; You, M. CRR9/CLPTM1L regulates cell survival signaling and is required for Ras transformation and lung tumorigenesis. Cancer Res. 2014, 74, 1116–1127. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.-R.; Chen, J.-C.; Li, M.-D.; Li, T.; Tan, Y.; Zhang, M. A meta-analysis of glutathione S-transferase M1 and T1 genetic polymorphism in relation to susceptibility to nasopharyngeal carcinoma. Int. J. Clin. Exp. Med. 2015, 8, 10626–10632. [Google Scholar]
- Cheng, D.; Hao, Y.; Zhou, W. IL-1α -889 C/T polymorphism and cancer susceptibility: A meta-analysis. OncoTargets Ther. 2014, 7, 2067–2074. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Peng, Q.; Lao, X.; Chen, Z.; Lu, Y.; Lao, X.; Mo, C.; Sui, J.; Wu, J.; Zhai, L.; et al. The association of interleukin-16 gene polymorphisms with IL-16 serum levels and risk of nasopharyngeal carcinoma in a Chinese population. Tumor Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 1917–1924. [Google Scholar] [CrossRef]
- Guo, X.-G.; Xia, Y. The Interleukin-18 promoter -607C>A polymorphism contributes to nasopharyngeal carcinoma risk: Evidence from a meta-analysis including 1,886 subjects. Asian Pac. J. Cancer Prev. APJCP 2013, 14, 7577–7581. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ren, D.; Li, Y.; Xu, J.; Liu, C.; Zhao, Y. Increased cancer risk associated with the -607C/A polymorphism in interleukin-18 gene promoter: An updated meta-analysis including 12,502 subjects. J. Balk. Union Oncol. 2015, 20, 902–917. [Google Scholar]
- Carrel, L.; Willard, H.F. X-inactivation profile reveals extensive variability in X-linked gene expression in females. Nature 2005, 434, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.H. Nasopharyngeal carcinoma (NPC). Adv. Cancer Res. 1972, 15, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.P.; Ho, J.H.; Saw, D.; Teoh, T.B. Carcinoma of the nasal and paranasal regions in rats fed Cantonese salted marine fish. IARC Sci. Publ. 1978, 20, 315–328. [Google Scholar]
- Yu, M.C.; Nichols, P.W.; Zou, X.N.; Estes, J.; Henderson, B.E. Induction of malignant nasal cavity tumours in Wistar rats fed Chinese salted fish. Br. J. Cancer 1989, 60, 198–201. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Luo, Y.; Christensson, B.; Drettner, B. Induction of nasal and nasopharyngeal tumours in Sprague-Dawley rats fed with Chinese salted fish. Acta Otolaryngol. (Stockh.) 1994, 114, 98–104. [Google Scholar] [CrossRef]
- Huang, D.P.; Ho, J.H.; Webb, K.S.; Wood, B.J.; Gough, T.A. Volatile nitrosamines in salt-preserved fish before and after cooking. Food Cosmet. Toxicol. 1981, 19, 167–171. [Google Scholar] [CrossRef]
- Zou, X.N.; Lu, S.H.; Liu, B. Volatile N-nitrosamines and their precursors in Chinese salted fish—A possible etological factor for NPC in china. Int. J. Cancer 1994, 59, 155–158. [Google Scholar] [CrossRef]
- Shao, Y.M.; Poirier, S.; Ohshima, H.; Malaveille, C.; Zeng, Y.; de Thé, G.; Bartsch, H. Epstein-Barr virus activation in Raji cells by extracts of preserved food from high risk areas for nasopharyngeal carcinoma. Carcinogenesis 1988, 9, 1455–1457. [Google Scholar] [CrossRef]
- Poirier, S.; Bouvier, G.; Malaveille, C.; Ohshima, H.; Shao, Y.M.; Hubert, A.; Zeng, Y.; de Thé, G.; Bartsch, H. Volatile nitrosamine levels and genotoxicity of food samples from high-risk areas for nasopharyngeal carcinoma before and after nitrosation. Int. J. Cancer 1989, 44, 1088–1094. [Google Scholar] [CrossRef]
- Bouvier, G.; Hergenhahn, M.; Polack, A.; Bornkamm, G.W.; de Thé, G.; Bartsch, H. Characterization of macromolecular lignins as Epstein-Barr virus inducer in foodstuff associated with nasopharyngeal carcinoma risk. Carcinogenesis 1995, 16, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Jeannel, D.; Hubert, A.; de Vathaire, F.; Ellouz, R.; Camoun, M.; Ben Salem, M.; Sancho-Garnier, H.; de-Thé, G. Diet, living conditions and nasopharyngeal carcinoma in Tunisia—A case-control study. Int. J. Cancer 1990, 46, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control (CDC). The Surgeon General’s 1989 Report on Reducing the Health Consequences of Smoking: 25 Years of Progress. MMWR Suppl. 1989, 38, 1–32. [Google Scholar]
- Cheng, Y.J.; Hildesheim, A.; Hsu, M.M.; Chen, I.H.; Brinton, L.A.; Levine, P.H.; Chen, C.J.; Yang, C.S. Cigarette smoking, alcohol consumption and risk of nasopharyngeal carcinoma in Taiwan. Cancer Causes Control CCC 1999, 10, 201–207. [Google Scholar] [CrossRef]
- Lin, J.-H.; Jiang, C.-Q.; Ho, S.-Y.; Zhang, W.-S.; Mai, Z.-M.; Xu, L.; Lo, C.-M.; Lam, T.-H. Smoking and nasopharyngeal carcinoma mortality: A cohort study of 101,823 adults in Guangzhou, China. BMC Cancer 2015, 15, 906. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.C.; Garabrant, D.H.; Huang, T.B.; Henderson, B.E. Occupational and other non-dietary risk factors for nasopharyngeal carcinoma in Guangzhou, China. Int. J. Cancer 1990, 45, 1033–1039. [Google Scholar] [CrossRef]
- Yuan, J.M.; Wang, X.L.; Xiang, Y.B.; Gao, Y.T.; Ross, R.K.; Yu, M.C. Non-dietary risk factors for nasopharyngeal carcinoma in Shanghai, China. Int. J. Cancer 2000, 85, 364–369. [Google Scholar] [CrossRef]
- Armstrong, R.W.; Armstrong, M.J.; Yu, M.C.; Henderson, B.E. Salted fish and inhalants as risk factors for nasopharyngeal carcinoma in Malaysian Chinese. Cancer Res. 1983, 43, 2967–2970. [Google Scholar]
- Demers, P.A.; Boffetta, P.; Kogevinas, M.; Blair, A.; Miller, B.A.; Robinson, C.F.; Roscoe, R.J.; Winter, P.D.; Colin, D.; Matos, E. Pooled reanalysis of cancer mortality among five cohorts of workers in wood-related industries. Scand. J. Work. Environ. Health 1995, 21, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.E.; Louie, E.; SooHoo Jing, J.; Buell, P.; Gardner, M.B. Risk factors associated with nasopharyngeal carcinoma. N. Engl. J. Med. 1976, 295, 1101–1106. [Google Scholar] [CrossRef]
- Hildesheim, A.; Dosemeci, M.; Chan, C.C.; Chen, C.J.; Cheng, Y.J.; Hsu, M.M.; Chen, I.H.; Mittl, B.F.; Sun, B.; Levine, P.H.; et al. Occupational exposure to wood, formaldehyde, and solvents and risk of nasopharyngeal carcinoma. Cancer Epidemiol. Prev. Biomark. 2001, 10, 1145–1153. [Google Scholar]
- Nam, J.M.; McLaughlin, J.K.; Blot, W.J. Cigarette smoking, alcohol, and nasopharyngeal carcinoma: A case-control study among U.S. whites. J. Natl. Cancer Inst. 1992, 84, 619–622. [Google Scholar] [CrossRef]
- Vaughan, T.L.; Shapiro, J.A.; Burt, R.D.; Swanson, G.M.; Berwick, M.; Lynch, C.F.; Lyon, J.L. Nasopharyngeal cancer in a low-risk population: Defining risk factors by histological type. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 1996, 5, 587–593. [Google Scholar]
- Khanim, F.; Yao, Q.Y.; Niedobitek, G.; Sihota, S.; Rickinson, A.B.; Young, L.S. Analysis of Epstein-Barr virus gene polymorphisms in normal donors and in virus-associated tumors from different geographic locations. Blood 1996, 88, 3491–3501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzellos, S.; Farrell, P.J. Epstein-barr virus sequence variation-biology and disease. Pathog. Basel Switz. 2012, 1, 156–174. [Google Scholar] [CrossRef] [Green Version]
- Feng, F.-T.; Cui, Q.; Liu, W.-S.; Guo, Y.-M.; Feng, Q.-S.; Chen, L.-Z.; Xu, M.; Luo, B.; Li, D.-J.; Hu, L.-F.; et al. A single nucleotide polymorphism in the Epstein-Barr virus genome is strongly associated with a high risk of nasopharyngeal carcinoma. Chin. J. Cancer 2015, 34, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Rey, J.; Xerri, L.; Bouabdallah, R.; Keuppens, M.; Brousset, P.; Meggetto, F. Detection of different clonal EBV strains in Hodgkin lymphoma and nasopharyngeal carcinoma tissues from the same patient. Br. J. Haematol. 2008, 142, 79–81. [Google Scholar] [CrossRef]
- Sitki-Green, D.; Covington, M.; Raab-Traub, N. Compartmentalization and transmission of multiple epstein-barr virus strains in asymptomatic carriers. J. Virol. 2003, 77, 1840–1847. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.J.; Brennan, R.; Miles, J.J.; Moss, D.J.; Burrows, J.M.; Burrows, S.R. Widespread sequence variation in Epstein-Barr virus nuclear antigen 1 influences the antiviral T cell response. J. Infect. Dis. 2008, 197, 1594–1597. [Google Scholar] [CrossRef] [Green Version]
- Young, L.S.; Dawson, C.W.; Clark, D.; Rupani, H.; Busson, P.; Tursz, T.; Johnson, A.; Rickinson, A.B. Epstein-Barr virus gene expression in nasopharyngeal carcinoma. J. Gen. Virol. 1988, 69 Pt 5, 1051–1065. [Google Scholar] [CrossRef]
- Brooks, L.; Yao, Q.Y.; Rickinson, A.B.; Young, L.S. Epstein-Barr virus latent gene transcription in nasopharyngeal carcinoma cells: Coexpression of EBNA1, LMP1, and LMP2 transcripts. J. Virol. 1992, 66, 2689–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sam, C.K.; Brooks, L.A.; Niedobitek, G.; Young, L.S.; Prasad, U.; Rickinson, A.B. Analysis of Epstein-Barr virus infection in nasopharyngeal biopsies from a group at high risk of nasopharyngeal carcinoma. Int. J. Cancer 1993, 53, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Gilligan, K.J.; Rajadurai, P.; Lin, J.C.; Busson, P.; Abdel-Hamid, M.; Prasad, U.; Tursz, T.; Raab-Traub, N. Expression of the Epstein-Barr virus BamHI A fragment in nasopharyngeal carcinoma: Evidence for a viral protein expressed in vivo. J. Virol. 1991, 65, 6252–6259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaussin, G.; Sbih-Lammali, F.; de Turenne-Tessier, M.; Bouguermouh, A.; Ooka, T. Expression of BARF1 gene encoded by Epstein-Barr virus in nasopharyngeal carcinoma biopsies. Cancer Res. 2000, 60, 5584–5588. [Google Scholar] [PubMed]
- Seto, E.; Yang, L.; Middeldorp, J.; Sheen, T.-S.; Chen, J.-Y.; Fukayama, M.; Eizuru, Y.; Ooka, T.; Takada, K. Epstein-Barr virus (EBV)-encoded BARF1 gene is expressed in nasopharyngeal carcinoma and EBV-associated gastric carcinoma tissues in the absence of lytic gene expression. J. Med. Virol. 2005, 76, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P. Nasopharyngeal carcinoma and the EBV-specific T cell response: Prospects for immunotherapy. Semin. Cancer Biol. 2002, 12, 463–471. [Google Scholar] [CrossRef]
- Hislop, A.D.; Taylor, G.S.; Sauce, D.; Rickinson, A.B. Cellular responses to viral infection in humans: Lessons from Epstein-Barr virus. Annu. Rev. Immunol. 2007, 25, 587–617. [Google Scholar] [CrossRef]
- Khanna, R.; Tellam, J.; Duraiswamy, J.; Cooper, L. Immunotherapeutic strategies for EBV-associated malignancies. Trends Mol. Med. 2001, 7, 270–276. [Google Scholar] [CrossRef]
- Smith, C.; Beagley, L.; Khanna, R. Acquisition of polyfunctionality by Epstein-Barr virus-specific CD8+ T cells correlates with increased resistance to galectin-1-mediated suppression. J. Virol. 2009, 83, 6192–6198. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.; Wakisaka, N.; Crough, T.; Peet, J.; Yoshizaki, T.; Beagley, L.; Khanna, R. Discerning regulation of cis- and trans-presentation of CD8+ T-cell epitopes by EBV-encoded oncogene LMP-1 through self-aggregation. Blood 2009, 113, 6148–6152. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.L.; Lu, J.H.; Cao, L.; Wu, M.H.; Peng, S.P.; Zhou, H.D.; Huang, C.; Yang, Y.X.; Zhou, Y.H.; Chen, Q.; et al. Genetic variations of EBV-LMP1 from nasopharyngeal carcinoma biopsies: Potential loss of T cell epitopes. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Medicas E Biol. 2008, 41, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Horst, D.; Burrows, S.R.; Gatherer, D.; van Wilgenburg, B.; Bell, M.J.; Boer, I.G.J.; Ressing, M.E.; Wiertz, E.J.H.J. Epstein-Barr virus isolates retain their capacity to evade T cell immunity through BNLF2a despite extensive sequence variation. J. Virol. 2012, 86, 572–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.-H.; Raykova, A.; Klinke, O.; Bernhardt, K.; Gärtner, K.; Leung, C.S.; Geletneky, K.; Sertel, S.; Münz, C.; Feederle, R.; et al. Spontaneous lytic replication and epitheliotropism define an Epstein-Barr virus strain found in carcinomas. Cell Rep. 2013, 5, 458–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gourzones, C.; Barjon, C.; Busson, P. Host-tumor interactions in nasopharyngeal carcinomas. Semin. Cancer Biol. 2012, 22, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Tang, K.F.; Chua, Y.N.; Lu, J.; Feng, P.; Chew, C.T.; Chan, S.H. Expression of interleukin-18 by nasopharyngeal carcinoma cells: A factor that possibly initiates the massive leukocyte infiltration. Hum. Pathol. 2004, 35, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Mrizak, D.; Martin, N.; Barjon, C.; Jimenez-Pailhes, A.-S.; Mustapha, R.; Niki, T.; Guigay, J.; Pancré, V.; de Launoit, Y.; Busson, P.; et al. Effect of nasopharyngeal carcinoma-derived exosomes on human regulatory T cells. J. Natl. Cancer Inst. 2015, 107, 363. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.F.; Tan, S.Y.; Chan, S.H.; Chong, S.M.; Loh, K.S.; Tan, L.K.; Hu, H. A distinct expression of CC chemokines by macrophages in nasopharyngeal carcinoma: Implication for the intense tumor infiltration by T lymphocytes and macrophages. Hum. Pathol. 2001, 32, 42–49. [Google Scholar] [CrossRef]
- Busson, P.; Braham, K.; Ganem, G.; Thomas, F.; Grausz, D.; Lipinski, M.; Wakasugi, H.; Tursz, T. Epstein-Barr virus-containing epithelial cells from nasopharyngeal carcinoma produce interleukin 1 alpha. Proc. Natl. Acad. Sci. USA 1987, 84, 6262–6266. [Google Scholar] [CrossRef] [Green Version]
- Duffield, A.S.; Ascierto, M.L.; Anders, R.A.; Taube, J.M.; McMiller, T.L.; Engle, E.L.; Meeker, A.K.; Berger, A.E.; Pardoll, D.M.; Ambinder, R.F.; et al. Abstract 4750: The immunosuppressive tumor microenvironment (TME) in nasopharyngeal carcinoma: Implications for immunotherapy. Cancer Res. 2018, 78, 4750. [Google Scholar] [CrossRef]
- Zhou, Y.; Xia, L.; Lin, J.; Wang, H.; Oyang, L.; Tan, S.; Tian, Y.; Su, M.; Wang, H.; Cao, D.; et al. Exosomes in Nasopharyngeal Carcinoma. J. Cancer 2018, 9, 767–777. [Google Scholar] [CrossRef]
- Ai, P.; Li, Z.; Jiang, Y.; Song, C.; Zhang, L.; Hu, H.; Wang, T. Tumor microenvironment contributes to Epstein-Barr virus anti-nuclear antigen-1 antibody production in nasopharyngeal carcinoma. Oncol. Lett. 2017, 14, 2458–2462. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, T.; Kondo, S.; Endo, K.; Nakanishi, Y.; Aga, M.; Kobayashi, E.; Hirai, N.; Sugimoto, H.; Hatano, M.; Ueno, T.; et al. Modulation of the tumor microenvironment by Epstein-Barr virus latent membrane protein 1 in nasopharyngeal carcinoma. Cancer Sci. 2018, 109, 272–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, W.K.J.; Chan, J.Y.K. Recent advances in the management of nasopharyngeal carcinoma. F1000Research 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, H.; Sun, X.; Wang, D.; Gu, Y.; Liu, Q.; Wang, H.; Han, W.; Fry, A. Salvage endoscopic nasopharyngectomy for local recurrent or residual nasopharyngeal carcinoma: A 10-year experience. Int. J. Clin. Oncol. 2017, 22, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.H.C.; Liew, Y.T.; Abu Bakar, M.Z.; Lim, E.Y.L.; Prepageran, N. A preliminary report on the role of endoscopic endonasal nasopharyngectomy in recurrent rT3 and rT4 nasopharyngeal carcinoma. Eur. Arch. Oto-Rhino-Laryngol. 2017, 274, 275–281. [Google Scholar] [CrossRef]
- You, R.; Zou, X.; Hua, Y.-J.; Han, F.; Li, L.; Zhao, C.; Hong, M.-H.; Chen, M.-Y. Salvage endoscopic nasopharyngectomy is superior to intensity-modulated radiation therapy for local recurrence of selected T1-T3 nasopharyngeal carcinoma—A case-matched comparison. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2015, 115, 399–406. [Google Scholar] [CrossRef]
- Tsang, R.K.; To, V.S.; Ho, A.C.; Ho, W.-K.; Chan, J.Y.; Wei, W.I. Early results of robotic assisted nasopharyngectomy for recurrent nasopharyngeal carcinoma. Head Neck 2015, 37, 788–793. [Google Scholar] [CrossRef]
- Richmon, J.D. Transoral palate-sparing nasopharyngectomy with the Flex® System: Preclinical study. Laryngoscope 2015, 125, 318–322. [Google Scholar] [CrossRef]
- Chan, J.Y.W.; Wong, S.T.S.; Chan, R.C.L.; Wei, W.I. Extracranial/intracranial vascular bypass and craniofacial resection: New hope for patients with locally advanced recurrent nasopharyngeal carcinoma. Head Neck 2016, 38 (Suppl. 1), E1404–E1412. [Google Scholar] [CrossRef]
- Lee, A.W.M.; Sze, W.M.; Au, J.S.K.; Leung, S.F.; Leung, T.W.; Chua, D.T.T.; Zee, B.C.Y.; Law, S.C.K.; Teo, P.M.L.; Tung, S.Y.; et al. Treatment results for nasopharyngeal carcinoma in the modern era: The Hong Kong experience. Int. J. Radiat. Oncol. Biol. Phys. 2005, 61, 1107–1116. [Google Scholar] [CrossRef]
- Al-Sarraf, M.; LeBlanc, M.; Giri, P.G.; Fu, K.K.; Cooper, J.; Vuong, T.; Forastiere, A.A.; Adams, G.; Sakr, W.A.; Schuller, D.E.; et al. Chemoradiotherapy versus radiotherapy in patients with advanced nasopharyngeal cancer: Phase III randomized Intergroup study 0099. J. Clin. Oncol. 1998, 16, 1310–1317. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.-T.; Ng, Q.-S.; Ivy, P.; Leong, S.-S.; Singh, O.; Chowbay, B.; Gao, F.; Thng, C.H.; Goh, B.-C.; Tan, D.S.-W.; et al. A Phase II study of pazopanib in Asian patients with recurrent/metastatic nasopharyngeal carcinoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 5481–5489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, E.P.; Ma, B.B.Y.; King, A.D.; Mo, F.; Chan, S.L.; Kam, M.K.M.; Loong, H.H.; Ahuja, A.T.; Zee, B.C.Y.; Chan, A.T.C. Hemorrhagic complications in a phase II study of sunitinib in patients of nasopharyngeal carcinoma who has previously received high-dose radiation. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2011, 22, 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.T.; Sheen, T.S.; Chen, C.L.; Lu, J.; Chang, Y.; Chen, J.Y.; Tsai, C.H. Profile of cytokine expression in nasopharyngeal carcinomas: A distinct expression of interleukin 1 in tumor and CD4+ T cells. Cancer Res. 1999, 59, 1599–1605. [Google Scholar] [PubMed]
- Lau, K.-M.; Cheng, S.H.; Lo, K.W.; Lee, S.A.K.W.; Woo, J.K.S.; van Hasselt, C.A.; Lee, S.P.; Rickinson, A.B.; Ng, M.H.L. Increase in circulating Foxp3+CD4+CD25(high) regulatory T cells in nasopharyngeal carcinoma patients. Br. J. Cancer 2007, 96, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.-T.; Ye, S.-B.; Liu, Y.-N.; He, J.; Chen, Q.-Y.; Mai, H.-Q.; Zhang, C.-X.; Cui, J.; Zhang, X.-S.; Busson, P.; et al. LMP1-mediated glycolysis induces myeloid-derived suppressor cell expansion in nasopharyngeal carcinoma. PLoS Pathog. 2017, 13, e1006503. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.M.; Lo, K.W.; Nicholls, J.M.; Huang, S.C.M.; Tsao, S.W. Chapter 3—Pathogenesis of Nasopharyngeal Carcinoma: Histogenesis, Epstein–Barr Virus Infection, and Tumor Microenvironment. In Nasopharyngeal Carcinoma; Lee, A.W.M., Lung, M.L., Ng, W.T., Eds.; Academic Press: Waltham, MA, USA, 2019; pp. 45–64. ISBN 978-0-12-814936-2. [Google Scholar]
- Cao, Y. EBV based cancer prevention and therapy in nasopharyngeal carcinoma. NPJ Precis. Oncol. 2017, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Teow, S.-Y.; Liew, K.; Khoo, A.S.-B.; Peh, S.-C. Pathogenic Role of Exosomes in Epstein-Barr Virus (EBV)-Associated Cancers. Int. J. Biol. Sci. 2017, 13, 1276–1286. [Google Scholar] [CrossRef] [Green Version]
- Hui, E.P.; Taylor, G.S.; Jia, H.; Ma, B.B.Y.; Chan, S.L.; Ho, R.; Wong, W.-L.; Wilson, S.; Johnson, B.F.; Edwards, C.; et al. Phase I trial of recombinant modified vaccinia ankara encoding Epstein-Barr viral tumor antigens in nasopharyngeal carcinoma patients. Cancer Res. 2013, 73, 1676–1688. [Google Scholar] [CrossRef] [Green Version]
- Chia, W.K.; Wang, W.-W.; Teo, M.; Tai, W.M.; Lim, W.T.; Tan, E.H.; Leong, S.S.; Sun, L.; Chen, J.J.; Gottschalk, S.; et al. A phase II study evaluating the safety and efficacy of an adenovirus-ΔLMP1-LMP2 transduced dendritic cell vaccine in patients with advanced metastatic nasopharyngeal carcinoma. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2012, 23, 997–1005. [Google Scholar] [CrossRef]
- Moralès, O.; Depil, S.; Mrizak, D.; Martin, N.; Ndour, P.A.; Dufosse, F.; Miroux, C.; Coll, J.; de Launoit, Y.; Auriault, C.; et al. EBV Latency II-derived peptides induce a specific CD4+ cytotoxic T-cell activity and not a CD4+ regulatory T-cell response. J. Immunother. 2012, 35, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Chia, W.-K.; Teo, M.; Wang, W.-W.; Lee, B.; Ang, S.-F.; Tai, W.-M.; Chee, C.-L.; Ng, J.; Kan, R.; Lim, W.-T.; et al. Adoptive T-cell transfer and chemotherapy in the first-line treatment of metastatic and/or locally recurrent nasopharyngeal carcinoma. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comoli, P.; Pedrazzoli, P.; Maccario, R.; Basso, S.; Carminati, O.; Labirio, M.; Schiavo, R.; Secondino, S.; Frasson, C.; Perotti, C.; et al. Cell therapy of stage IV nasopharyngeal carcinoma with autologous Epstein-Barr virus-targeted cytotoxic T lymphocytes. J. Clin. Oncol. 2005, 23, 8942–8949. [Google Scholar] [CrossRef] [PubMed]
- Secondino, S.; Zecca, M.; Licitra, L.; Gurrado, A.; Schiavetto, I.; Bossi, P.; Locati, L.; Schiavo, R.; Basso, S.; Baldanti, F.; et al. T-cell therapy for EBV-associated nasopharyngeal carcinoma: Preparative lymphodepleting chemotherapy does not improve clinical results. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2012, 23, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [Green Version]
- Walunas, T.L.; Lenschow, D.J.; Bakker, C.Y.; Linsley, P.S.; Freeman, G.J.; Green, J.M.; Thompson, C.B.; Bluestone, J.A. CTLA-4 can function as a negative regulator of T cell activation. Immunity 1994, 1, 405–413. [Google Scholar] [CrossRef]
- Ahmadzadeh, M.; Johnson, L.A.; Heemskerk, B.; Wunderlich, J.R.; Dudley, M.E.; White, D.E.; Rosenberg, S.A. Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood 2009, 114, 1537–1544. [Google Scholar] [CrossRef]
- Chen, B.J.; Chapuy, B.; Ouyang, J.; Sun, H.H.; Roemer, M.G.M.; Xu, M.L.; Yu, H.; Fletcher, C.D.M.; Freeman, G.J.; Shipp, M.A.; et al. PD-L1 expression is characteristic of a subset of aggressive B-cell lymphomas and virus-associated malignancies. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 3462–3473. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.-C.; Hsiao, J.-R.; Chang, K.-C.; Wu, Y.-H.; Su, I.-J.; Jin, Y.-T.; Chang, Y. Increase of programmed death-1-expressing intratumoral CD8 T cells predicts a poor prognosis for nasopharyngeal carcinoma. Mod. Pathol. 2010, 23, 1393–1403. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.-Y.; Guo, S.-S.; Zhang, Y.; Lu, J.-B.; Chen, Q.-Y.; Tang, L.-Q.; Zhang, L.; Liu, L.-T.; Zhang, L.; Mai, H.-Q. Tumor CTLA-4 overexpression predicts poor survival in patients with nasopharyngeal carcinoma. Oncotarget 2016, 7, 13060–13068. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Fogg, M.; Wirth, L.J.; Daley, H.; Ritz, J.; Posner, M.R.; Wang, F.C.; Lorch, J.H. Epstein-Barr virus-specific adoptive immunotherapy for recurrent, metastatic nasopharyngeal carcinoma. Cancer 2017, 123, 2642–2650. [Google Scholar] [CrossRef] [PubMed]
- Ning, N.; Pan, Q.; Zheng, F.; Teitz-Tennenbaum, S.; Egenti, M.; Yet, J.; Li, M.; Ginestier, C.; Wicha, M.S.; Moyer, J.S.; et al. Cancer stem cell vaccination confers significant antitumor immunity. Cancer Res. 2012, 72, 1853–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.P.; Othus, M.; Chae, Y.K.; Giles, F.J.; Hansel, D.E.; Singh, P.P.; Fontaine, A.; Shah, M.H.; Kasi, A.; Baghdadi, T.A.; et al. A Phase II Basket Trial of Dual Anti-CTLA-4 and Anti-PD-1 Blockade in Rare Tumors (DART SWOG 1609) in Patients with Nonpancreatic Neuroendocrine Tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 2290–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.-Y.; Bay, B.-H.; Olivo, M. Biodistribution and photodynamic therapy with hypericin in a human NPC murine tumor model. Int. J. Oncol. 2003, 22, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Lofgren, L.A.; Hallgren, S.; Nilsson, E.; Westerborn, A.; Nilsson, C.; Reizenstein, J. Photodynamic therapy for recurrent nasopharyngeal cancer. Arch. Otolaryngol. Head Neck Surg. 1995, 121, 997–1002. [Google Scholar] [CrossRef]
- Tong, M.C.; van Hasselt, C.A.; Woo, J.K. Preliminary results of photodynamic therapy for recurrent nasopharyngeal carcinoma. Eur. Arch. Oto-Rhino-Laryngol. 1996, 253, 189–192. [Google Scholar] [CrossRef]
- Li, L.-B.; Luo, R.-C.; Liao, W.-J.; Zhang, M.-J.; Luo, Y.-L.; Miao, J.-X. Clinical study of Photofrin photodynamic therapy for the treatment of relapse nasopharyngeal carcinoma. Photodiagn. Photodyn. Ther. 2006, 3, 266–271. [Google Scholar] [CrossRef]
- Indrasari, S.R.; Timmermans, A.J.; Wildeman, M.A.; Karakullukcu, M.B.; Herdini, C.; Hariwiyanto, B.; Tan, I.B. Remarkable response to photodynamic therapy in residual T4N0M0 nasopharyngeal carcinoma: A case report. Photodiagn. Photodyn. Ther. 2012, 9, 319–320. [Google Scholar] [CrossRef]
- Stoker, S.D.; Indrasari, S.R.; Herdini, C.; Hariwiyanto, B.; Karakullukcu, B.; Dhamiyati, W.; Widayati, K.; Romdhoni, A.C.; Fles, R.; Haryana, S.M.; et al. Photodynamic therapy as salvage therapy for patients with nasopharyngeal carcinoma experiencing local failures following definitive radiotherapy. Photodiagn. Photodyn. Ther. 2015, 12, 519–525. [Google Scholar] [CrossRef]
- Nyst, H.J.; Wildeman, M.A.; Indrasari, S.R.; Karakullukcu, B.; van Veen, R.L.P.; Adham, M.; Stewart, F.A.; Levendag, P.C.; Sterenborg, H.J.C.M.; Tan, I.B. Temoporfin mediated photodynamic therapy in patients with local persistent and recurrent nasopharyngeal carcinoma after curative radiotherapy: A feasibility study. Photodiagn. Photodyn. Ther. 2012, 9, 274–281. [Google Scholar] [CrossRef]
- Succo, G.; Rosso, S.; Fadda, G.L.; Fantini, M.; Crosetti, E. Salvage photodynamic therapy for recurrent nasopharyngeal carcinoma. Photodiagn. Photodyn. Ther. 2014, 11, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.W.K.; Chu, E.S.M.; Yow, C.M.N.; Chen, J.Y. Photodynamic effects on nasopharyngeal carcinoma (NPC) cells with 5-aminolevulinic acid or its hexyl ester. Cancer Lett. 2006, 242, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Castano, A.P.; Mroz, P.; Hamblin, M.R. Photodynamic therapy and anti-tumour immunity. Nat. Rev. Cancer 2006, 6, 535–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mroz, P.; Hashmi, J.T.; Huang, Y.-Y.; Lange, N.; Hamblin, M.R. Stimulation of anti-tumor immunity by photodynamic therapy. Expert Rev. Clin. Immunol. 2011, 7, 75–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turubanova, V.D.; Balalaeva, I.V.; Mishchenko, T.A.; Catanzaro, E.; Alzeibak, R.; Peskova, N.N.; Efimova, I.; Bachert, C.; Mitroshina, E.V.; Krysko, O.; et al. Immunogenic cell death induced by a new photodynamic therapy based on photosens and photodithazine. J. Immunother. Cancer 2019, 7, 350. [Google Scholar] [CrossRef]
- Quilbe, A.; Moralès, O.; Baydoun, M.; Kumar, A.; Mustapha, R.; Murakami, T.; Leroux, B.; de Schutter, C.; Thecua, E.; Ziane, L.; et al. An Efficient Photodynamic Therapy Treatment for Human Pancreatic Adenocarcinoma. J. Clin. Med. 2020, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Reginato, E.; Wolf, P.; Hamblin, M.R. Immune response after photodynamic therapy increases anti-cancer and anti-bacterial effects. World J. Immunol. 2014, 4, 1–11. [Google Scholar] [CrossRef]
- Koon, H.-K.; Lo, K.-W.; Leung, K.-N.; Lung, M.L.; Chang, C.C.-K.; Wong, R.N.-S.; Leung, W.-N.; Mak, N.-K. Photodynamic therapy-mediated modulation of inflammatory cytokine production by Epstein-Barr virus-infected nasopharyngeal carcinoma cells. Cell. Mol. Immunol. 2010, 7, 323–326. [Google Scholar] [CrossRef] [Green Version]
- Indrasari, S.R.; Sastrowijoto, S.; Astuti, I.; Hariwiyanto, B.; Tan T, B. Clinical Study on Photodynamic Therapy (PDT) In Patients with Nasopharyngeal Carcinoma in YogyakartaReview of Life Expectancy and Cytokine Level. J. Otolaryngol.-ENT Res. 2016, 4. [Google Scholar] [CrossRef]
- Lobo, A.C.S.; Gomes-da-Silva, L.C.; Rodrigues-Santos, P.; Cabrita, A.; Santos-Rosa, M.; Arnaut, L.G. Immune Responses after Vascular Photodynamic Therapy with Redaporfin. J. Clin. Med. 2019, 9, 104. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Characteristics | ||
|---|---|---|
| T Stage | T1 | The tumour is within the nasopharynx, or it has grown into the oropharynx and/or nasal cavity, but no extension into the parapharyngeal space (soft tissue space behind and to the side of the pharynx). |
| T2 | The tumour extends into the parapharyngeal space. | |
| T3 | The tumour has grown into the bone of the skull base and/or the sinuses. | |
| T4 | The tumour has grown into the skull and/or involves the cranial nerves, hypopharynx and eye socket. Alternatively, it has extended to the infratemporal fossa or masticator space. | |
| N Stage | N0 | No evidence of cancer spread to LNs 1 in the neck or retropharyngeal space. |
| N1 | Presence of cancer in the LNs on one side of the neck (6 cm or less in size) and above the clavicle (supraclavicular fossa). The LNs at this stage should be found in the retropharyngeal space (6 cm or less in size, one side or both). | |
| N2 | Presence of cancer in the LNs on both sides of the neck (biggest LN is 6 cm or less) and above the supraclavicular fossa. | |
| N3a | Presence of a LN with cancer bigger than 6 cm. | |
| N3b | Presence of a LN of any size that is far down the neck, just above the clavicle. | |
| M Stage | M0 | No evidence of distant spread outside the head and neck. |
| M1 | Evidence of spread outside the head and neck. |
| Phase | Status | Treatment Tested | Patient Details | Aim of the Study | Reference |
|---|---|---|---|---|---|
| I | Completed | EBV-specific adoptive T cell immunotherapy | 28 relapsed or metastatic NPC patients | To determine the safety of EBV-based adoptive transfer immunotherapy in NPC | NCT00431210 [103] |
| I | Active, not recruiting | EBV-specific T cells (2 antigens) that have an extra T cell receptor named DNT ± chemo lymphodepletion beforehand (Cyclophosphamide and fludarabine) | 14 participants with advanced NPC | To examine efficacy of EBV-specific T cells in NPC patients and determine if lymphodepleting chemotherapy before T cell infusion increases treatment efficacy | NCT02065362 |
| I | Recruiting | CAR-T cells (recognise EpCAM) | 30 NPC and breast cancer patients | Determine if treatment is well tolerated, the dosage and the adverse effects | NCT02915445 |
| I | Completed | Using two variants of LMP2 peptide vaccine | 99 patients with a high-risk of NPC recurrence | Evaluate the immunologic effectiveness of peptide immunisation in adjuvant settings in NPC | NCT00078494 |
| I/II | Recruiting | LMP1-CAR-T cells | 20 patients with EBV associated malignant tumours (nasopharyngeal neoplasms) | Evaluate safety and efficacy of designed LMP1-CAR-T cells in the treatment of EBV associated malignant tumours. | NCT02980315 |
| I/II | Recruiting | High-activity NKs | 20 NPC patients with small metastases | Assessment of the safety of high activity NKs on NPC patients | NCT03007836 |
| I/II | Completed | Cancer stem cell (CSC) vaccine | 40 metastatic NPC patients | To demonstrate that cytotoxic T cells generated after CSC vaccination are capable of specific killing of CSCs and conferring anti-tumour immunity | NCT02115958 [104] |
| II | Active, not recruiting | EBV-specific adoptive T cell immunotherapy | 20 relapsed or metastatic NPC patients | To determine effectiveness and safety of EBV-based adoptive transfer immunotherapy in NPC | NCT00834093 [103] |
| II | Recruiting | Combinations of Dendritic cells and Cytokine-induced Killer Cells (DC-CIK) treatment in solid tumours | 200 patients with treatment-refractory solid tumours: Colorectal cancer Renal cell Carcinoma Nasopharyngeal carcinoma Lung cancer | Aim is to investigate the efficacy of concurrent chemotherapy with DC-CIK and CIK treatment in patients with treatment-refractory solid tumours | NCT03047525 |
| II | Recruiting | Cisplatin and CRT ± TILs | 116 patients with only locoregionally advanced high-risk NPC | The Phase I results showed that TILs following CRT resulted in sustained anti-tumour activity and anti-EBV immune responses with good tolerance | NCT02421640 |
| II | Recruiting | (cisplatin) CRT ± nivolumab | 40 NPC patients ranging from low stage II to high stage IVB | Establish how well nivolumab and chemotherapy work to treat advanced NPC | NCT03267498 |
| II | Not yet recruiting | Pembrolizumab | 63 patients with detectable levels of EBV DNA in plasma after CRT. No residual disease and/or metastases | Examine efficacy and safety of pembrolizumab on NPC patients | NCT03544099 |
| II | Recruiting | Ipilimumab and nivolumab | 35 patients with advanced NPC | Test a combination of ipilimumab and nivolumab in EBV+ NPC | NCT03097939 |
| III | Recruiting | Chemotherapy (Gemcitabine and IV carboplatin) + autologous EBV-specific cytotoxic T cells | 330 participants with advanced NPC | Assess the efficacy of CTL following first line chemotherapy in prolonging overall survival of NPC patients | NCT02578641 Phase II complete trial [94] |
| III | Recruiting | Camrelizumab (PD-1 Antibody) after chemoradiotherapy | 400 patients with stage III-IVA non-metastatic NPC | Investigate whether adjuvant PD-1 antibody treatment could improve survival | NCT03427827 |
| II | Recruiting | Nivolumab and ipilimumab | Patients with rare tumours including NPC | Evaluate the efficacy of a combination of nivolumab and ipilimumab on hindering tumour cell growth | NCT02834013 [105] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renaud, S.; Lefebvre, A.; Mordon, S.; Moralès, O.; Delhem, N. Novel Therapies Boosting T Cell Immunity in Epstein Barr Virus-Associated Nasopharyngeal Carcinoma. Int. J. Mol. Sci. 2020, 21, 4292. https://doi.org/10.3390/ijms21124292
Renaud S, Lefebvre A, Mordon S, Moralès O, Delhem N. Novel Therapies Boosting T Cell Immunity in Epstein Barr Virus-Associated Nasopharyngeal Carcinoma. International Journal of Molecular Sciences. 2020; 21(12):4292. https://doi.org/10.3390/ijms21124292
Chicago/Turabian StyleRenaud, Sarah, Anthony Lefebvre, Serge Mordon, Olivier Moralès, and Nadira Delhem. 2020. "Novel Therapies Boosting T Cell Immunity in Epstein Barr Virus-Associated Nasopharyngeal Carcinoma" International Journal of Molecular Sciences 21, no. 12: 4292. https://doi.org/10.3390/ijms21124292
APA StyleRenaud, S., Lefebvre, A., Mordon, S., Moralès, O., & Delhem, N. (2020). Novel Therapies Boosting T Cell Immunity in Epstein Barr Virus-Associated Nasopharyngeal Carcinoma. International Journal of Molecular Sciences, 21(12), 4292. https://doi.org/10.3390/ijms21124292