A New Selective PPARγ Modulator Inhibits Triglycerides Accumulation during Murine Adipocytes’ and Human Adipose-Derived Mesenchymal Stem Cells Differentiation

 ,
,  ,
, 
 ,
, 
Abstract
:1. Introduction
2. Results
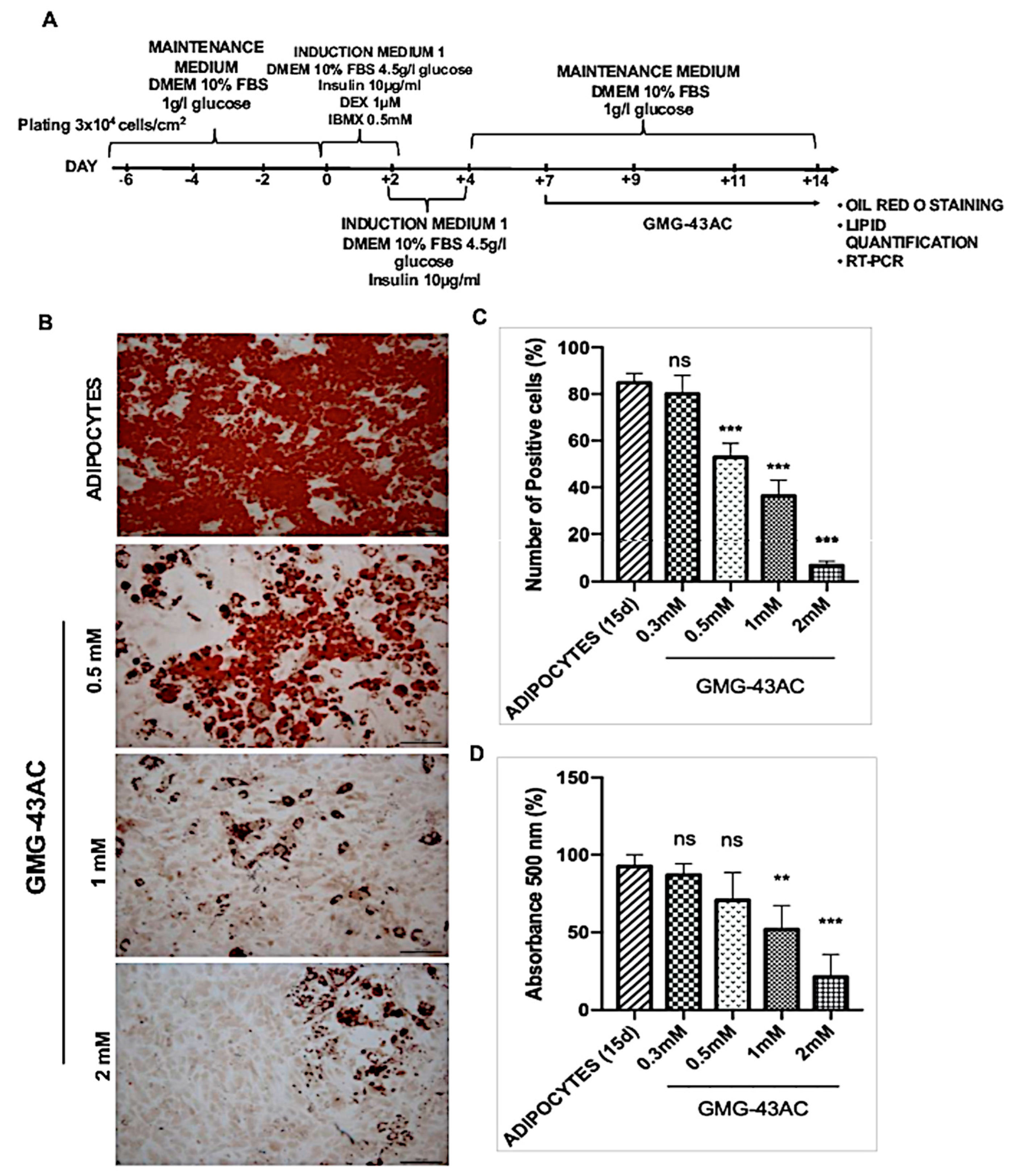
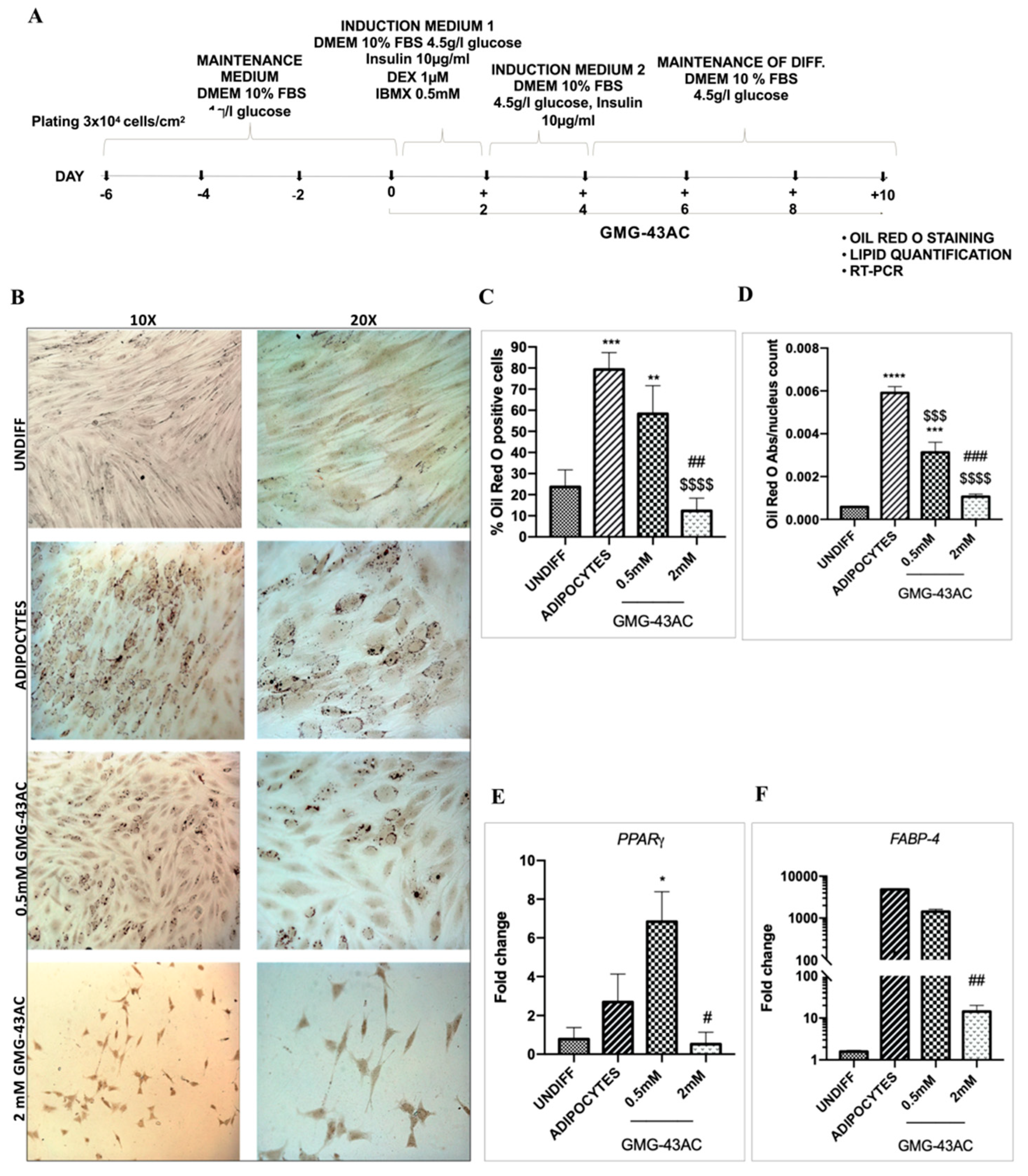
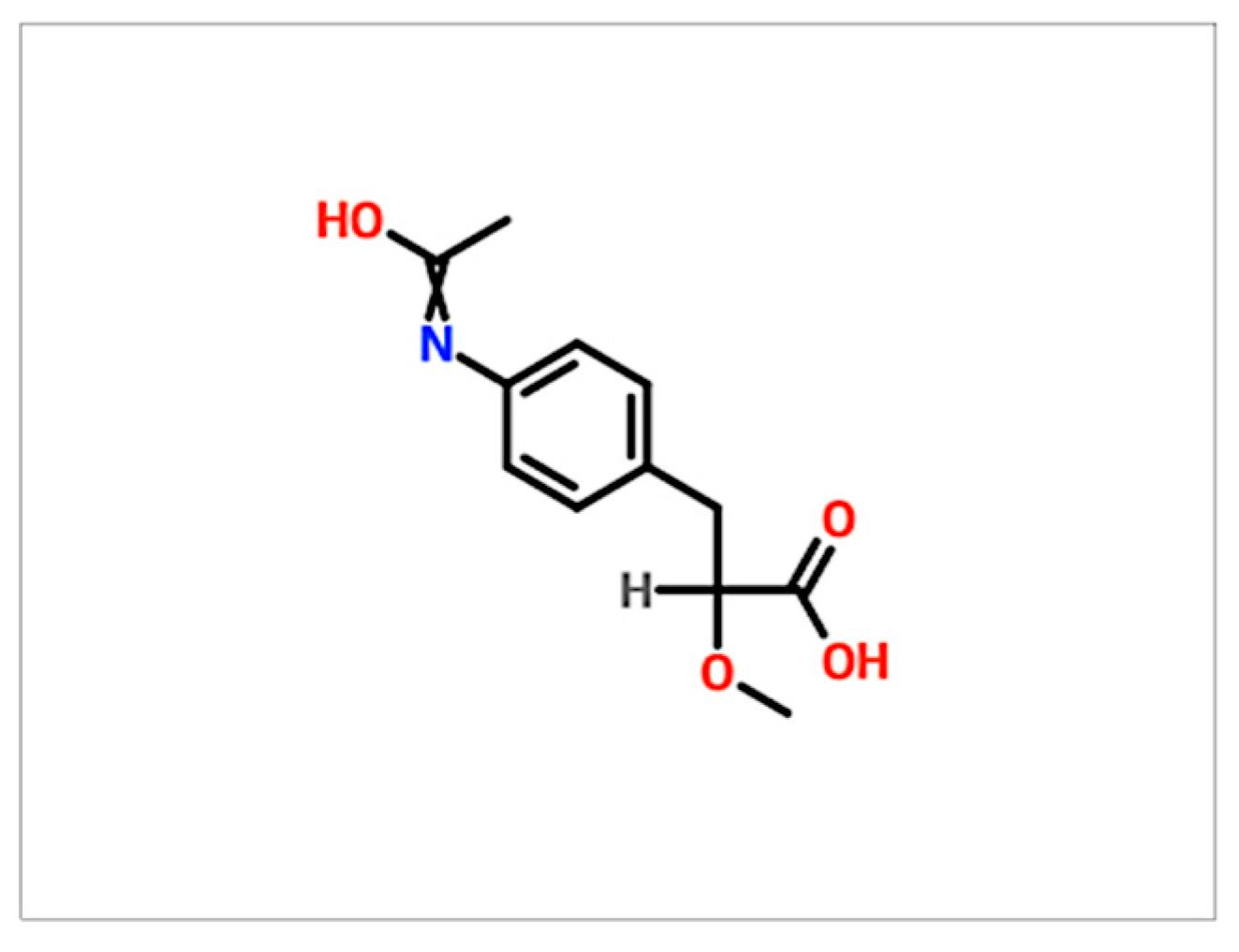
2.1. GMG-43AC Inhibits Triglycerides Accumulation in Murine 3T3-L1 Cells
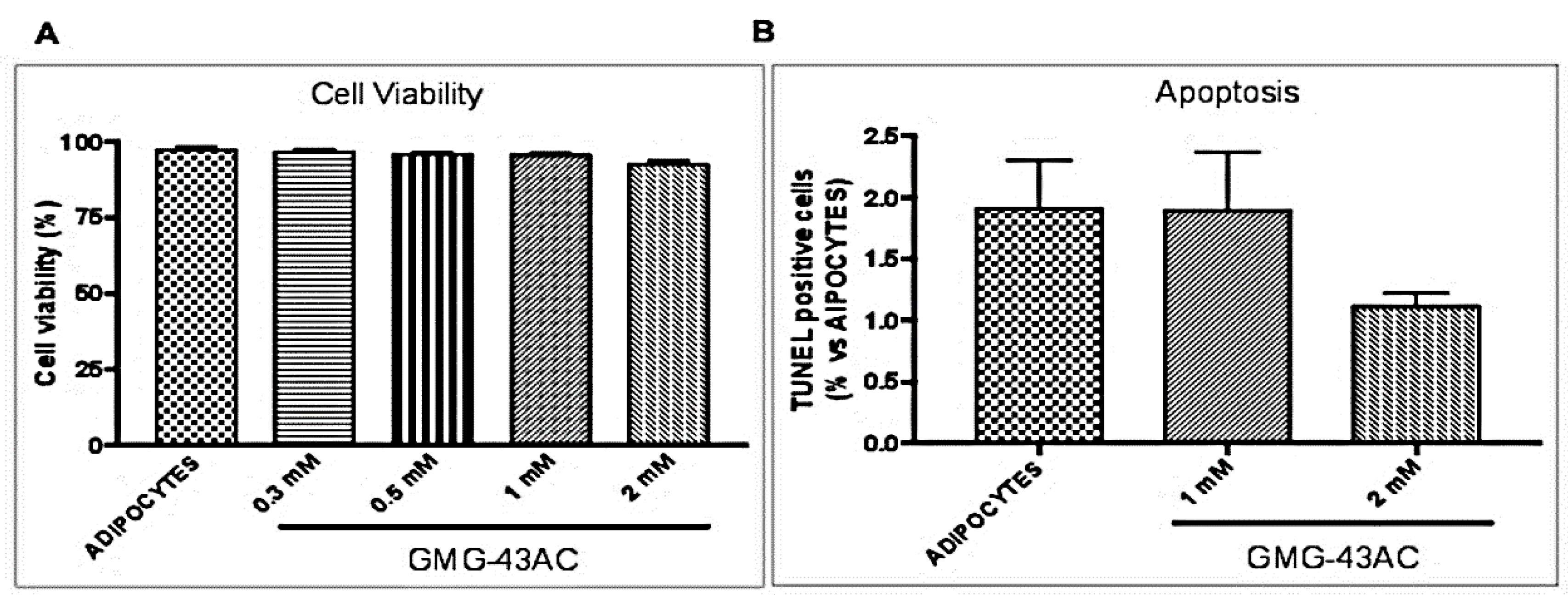
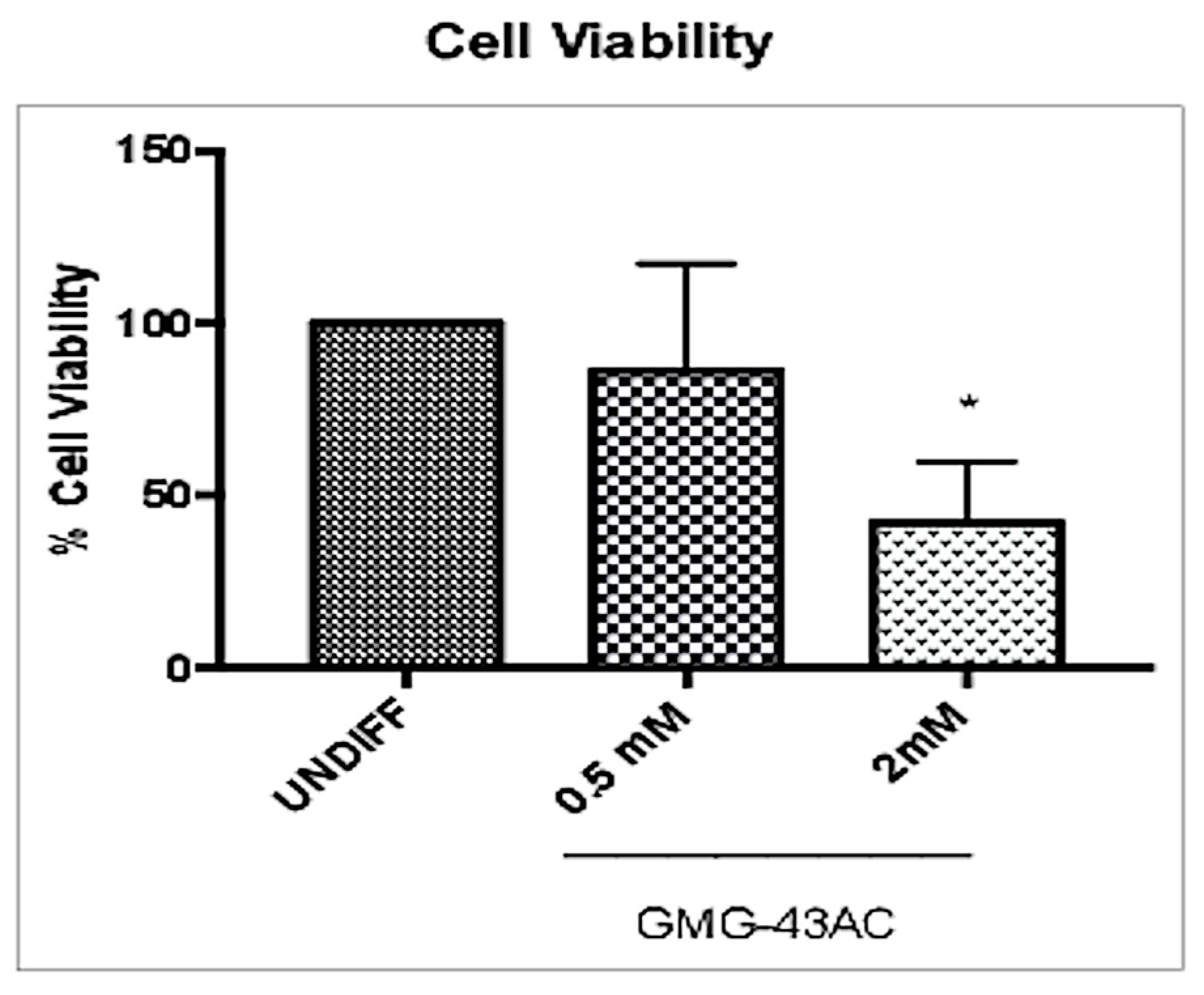
2.2. GMG-43AC Does Not Influence Viability of 3T3-L1 Cells
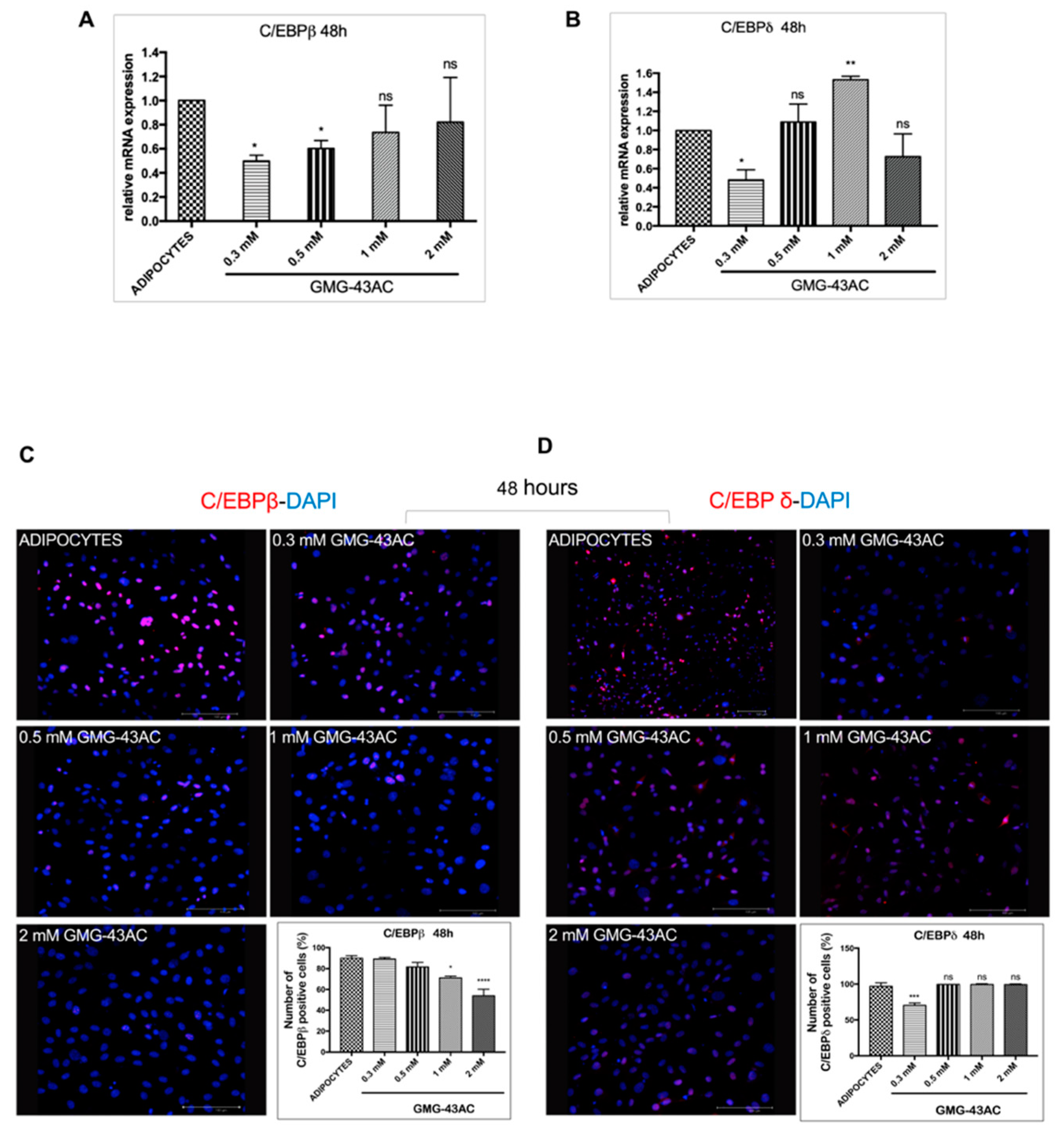
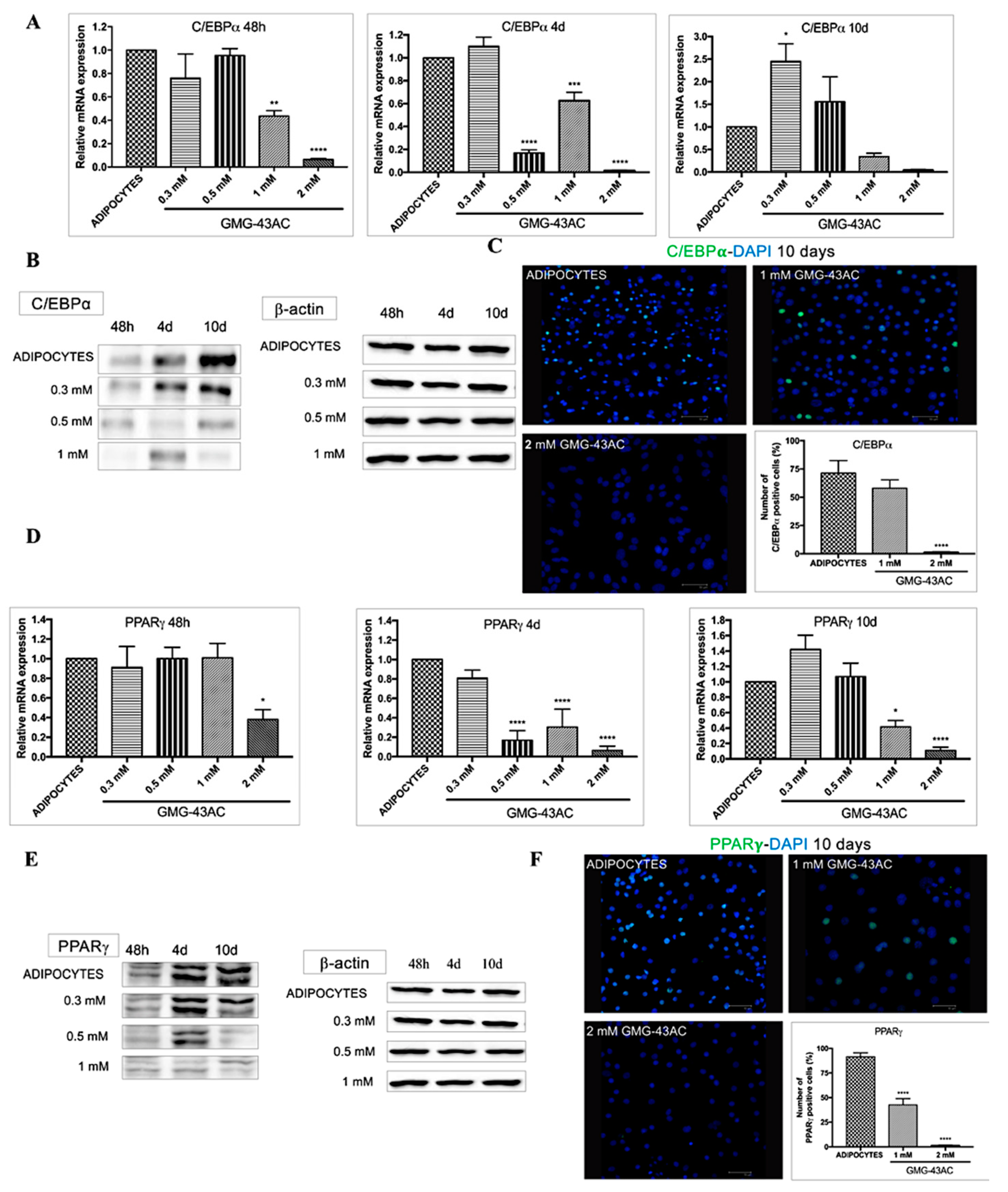
2.3. GMG-43AC Influences the Expression of Early Adipocytes Markers
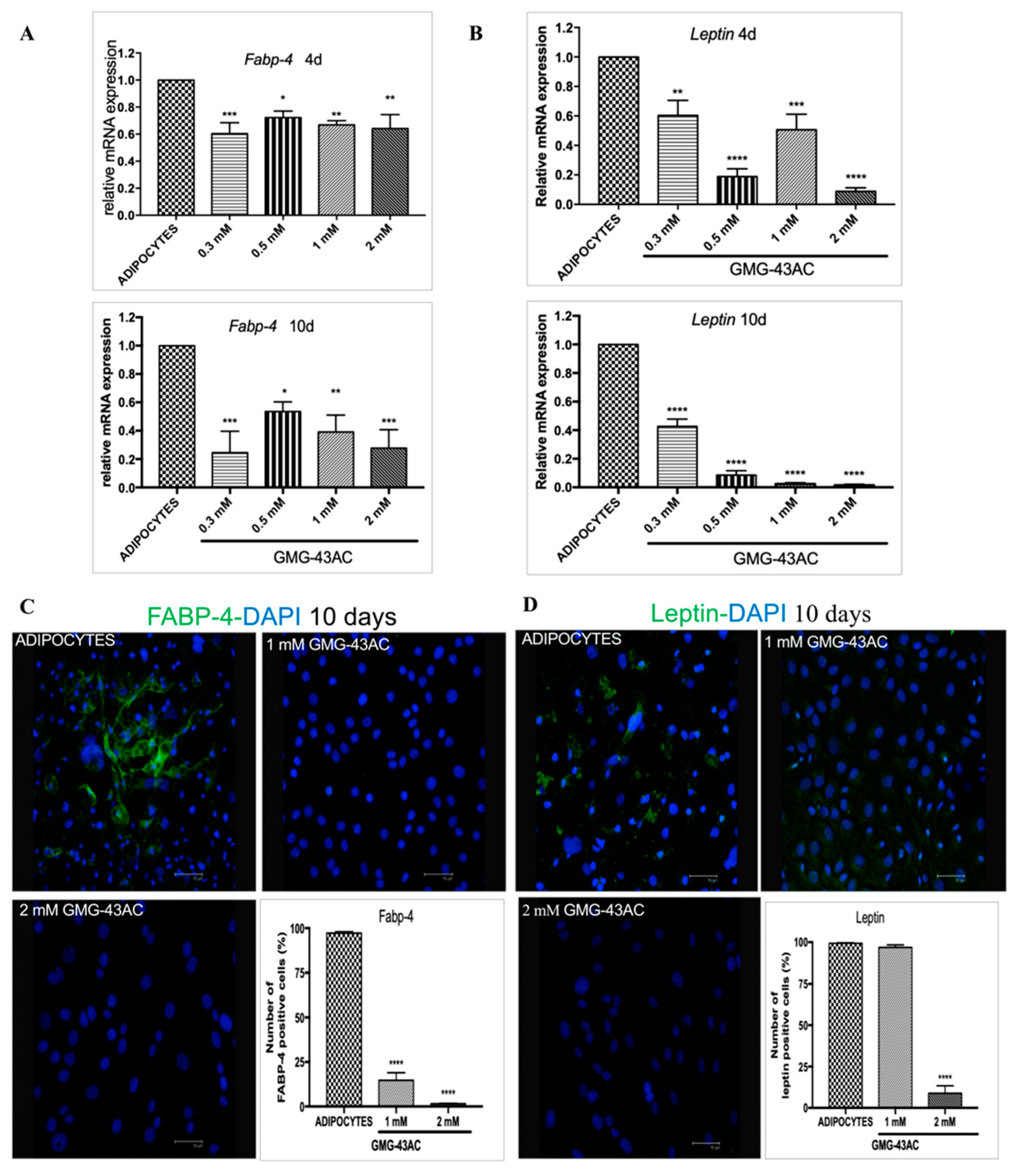
2.4. GMG-43AC Down-Regulates the Expression of Late Markers of Adipose Differentiation
2.5. GMG-43AC Inhibits the Expression and Modifies the Localization of Adipocyte-Specific Markers
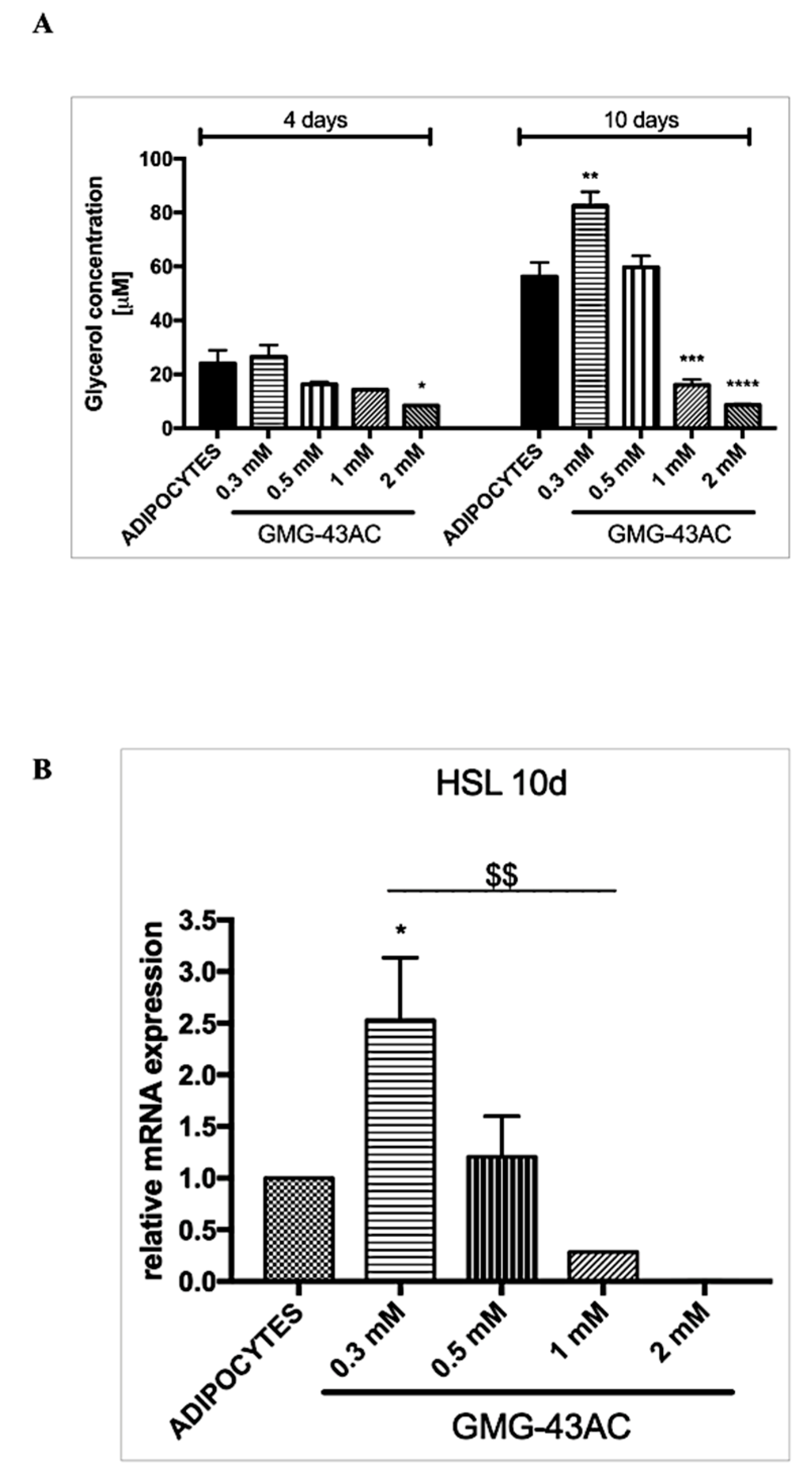
2.6. GMG-43AC Induces Lipolysis and Influences the Hsl Gene
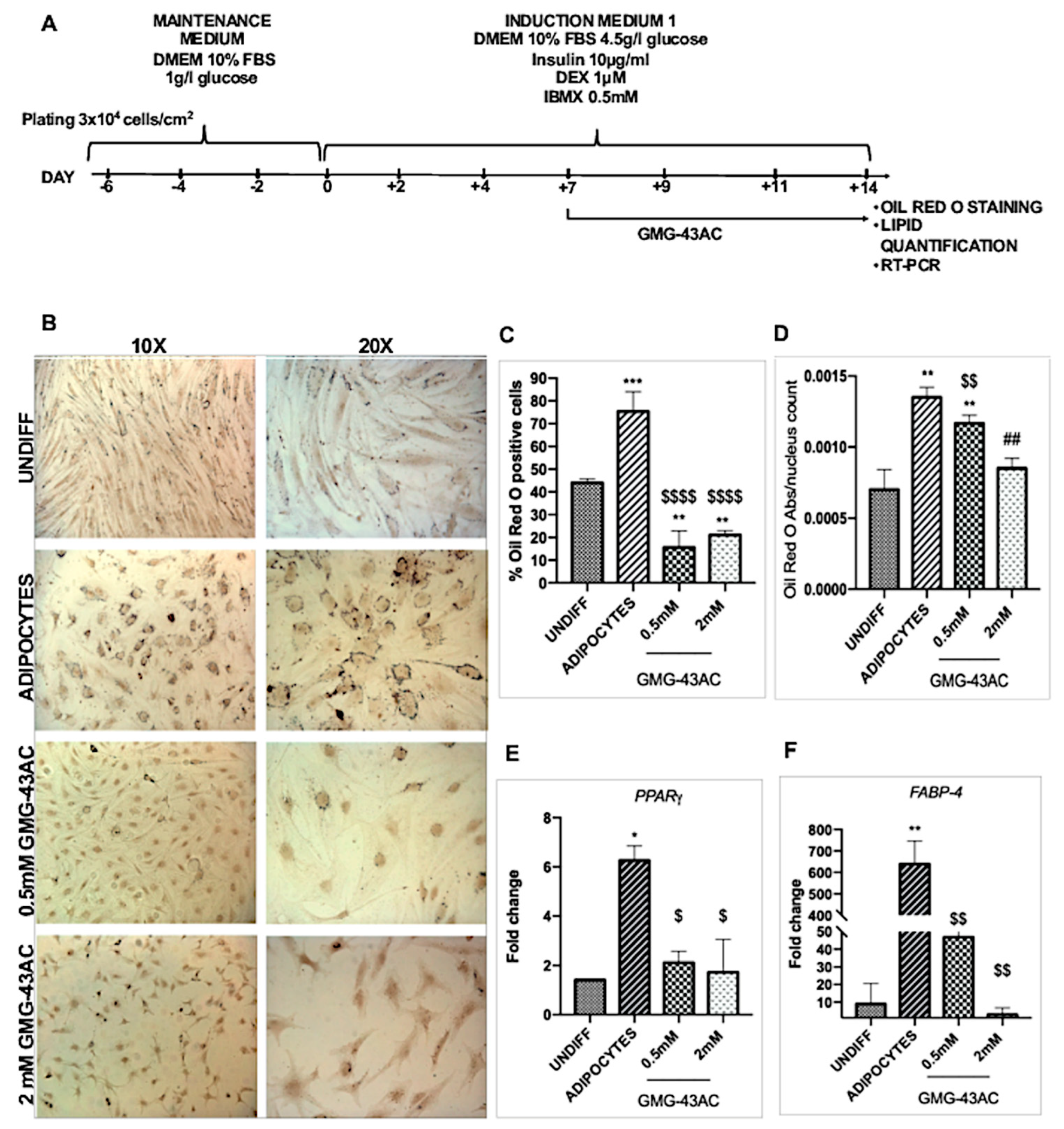
2.7. Reversion of Adipogenesis Process by GMG-43AC
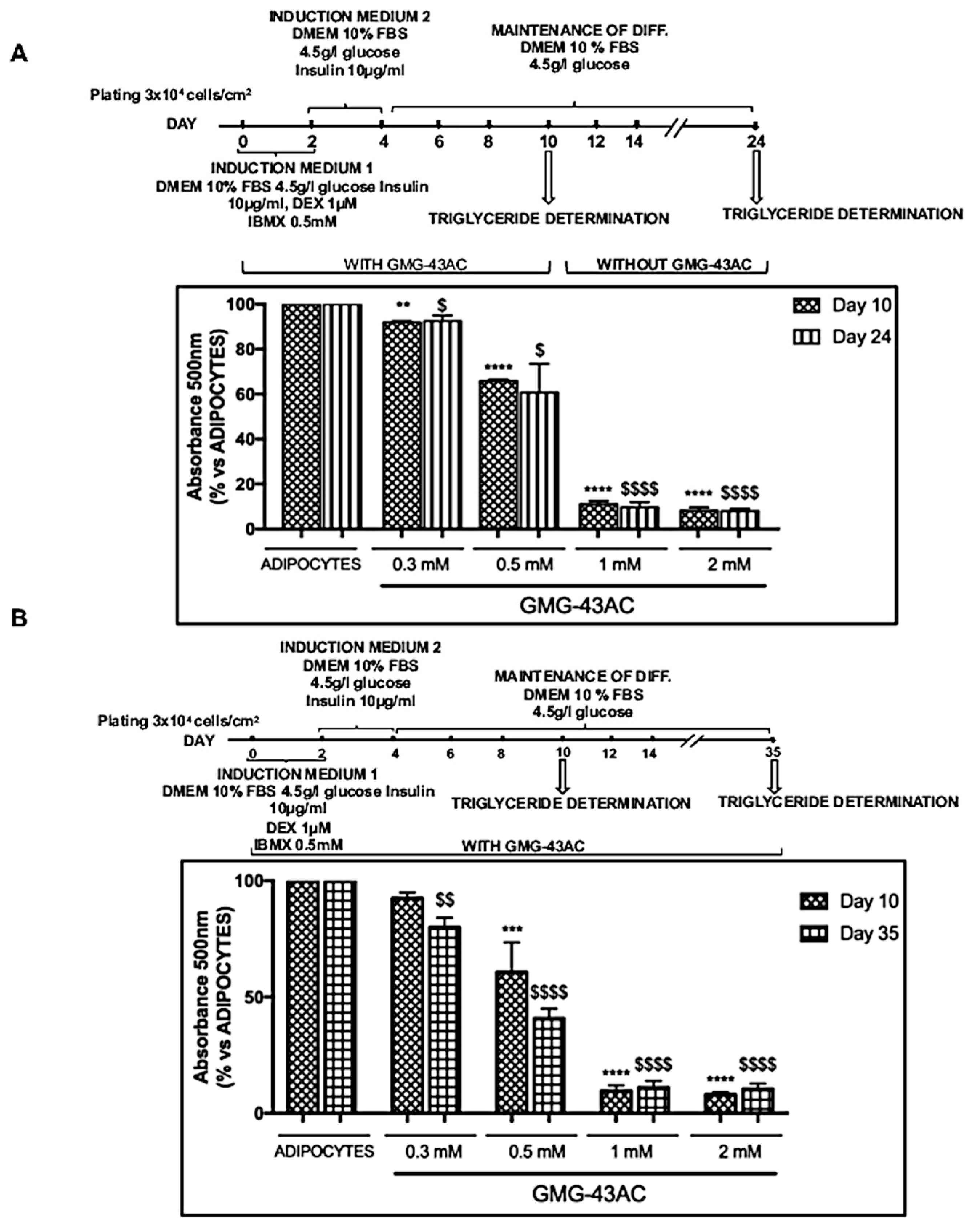
2.8. The Inhibition of Adipogenesis by GMG-43AC Is Not Reversible and Can Persist in a Long-Term Observational Period
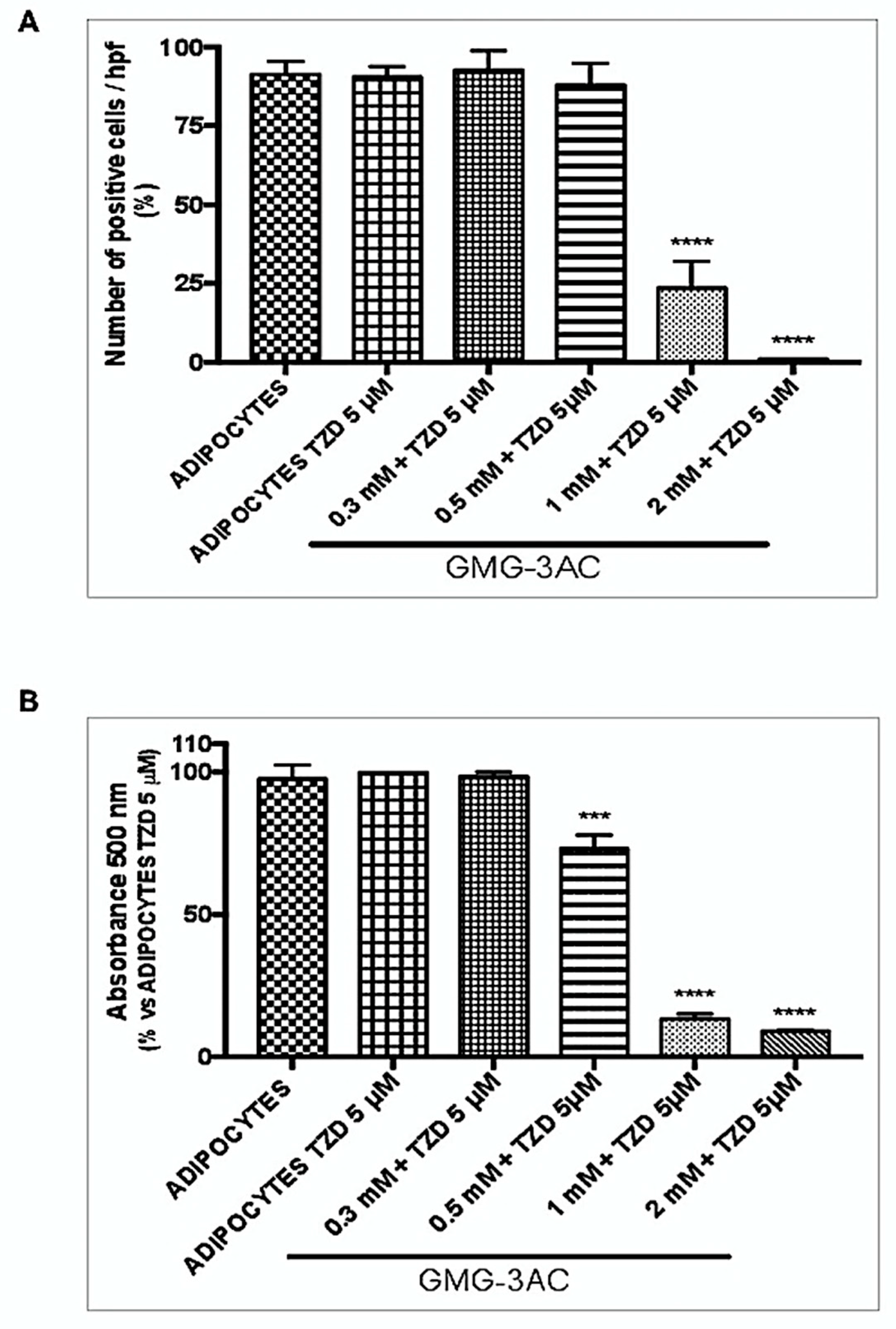
2.9. GMG-43AC Inhibits Lipids Accumulation and Leads to a Loss of Accumulated Triglycerides in the Differentiation Induced by Troglitazone
2.10. GMG-43AC Inhibits Triglycerides Accumulation and Has an Effect on the Expression of Adipocyte-Specific Genes in Human Adipose Derived Stem Cells (hADSCs)
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Induction of Differentiation in 3T3-L1 Cells
4.2. Cell Cultures and Induction of Differentiation in hADSCs Cells
4.3. Oil Red-O Staining and Quantification of Lipid Accumulation in Adipocytes
4.4. Immunofluorescence in 3T3-L1 Cells
4.5. Western Blot Analysis
4.6. Real-Time PCR Analysis
4.7. Lipolysis Measurement
4.8. Cell Viability and Terminal deoxynucleotidyl transferase dUTP Nick end Labeling Assay
4.9. MTT Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| hADSCs | human Adipose-Derived Stem Cells |
| PA | Propanoic Acid |
| BMI | Body Mass Index |
| C/EBP | CCAAT/enhancer-binding protein α, β, δ |
| PPAR γ | Peroxisome proliferator-activated receptor γ |
| FABP-4 | Fatty acid binding protein |
| HSL | Hormone-sensitive lipase gene |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
| TZD | Troglitazone |
| DEXA | Dexamethasone |
| DMSO | Dimethyl Sulfoxide; IBMX isobutil-1-metylxantine |
| DAPI | 4′,6′-diamidino-2-phenyl-indole |
| FBS | Fetal bovine serum |
| NHK | normal human keratinocytes |
| ECM | Extracellular Matrix |
| NP-40 | Nonidet P-40 |
| MSC | Mesenchymal stem cells |
| HCL | Hydrogen Chloride |
| DMEM | Dulbecco’s modified Eagle’s medium |
| PBS | Phosphate Buffer Solution |
| BSA | Bovine Serum Albumin |
References
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Lee, J.; Hong, V.S.; Choi, J.S.; Lee, T.Y.; Jang, B.C. Identification of KMU-3, a novel derivative of gallic acid, as an inhibitor of adipogenesis. PLoS ONE 2014, 9, e109344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.J.; Song, J.H.; Shon, M.S.; Kim, H.O.; Kwon, O.J.; Roh, S.S.; Kim, C.Y.; Kim, G.N. Anti-Adipogenic Effects of Ethanol Extracts Prepared from Selected Medicinal Herbs in 3T3-L1 Cells. Prev. Nutr. Food Sci. 2016, 21, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Sarbatly, R.; Krishnaiah, D.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.B.; Petersen, R.K.; Larsen, B.M.; Bartkova, J.; Alsner, J.; Kristiansen, K. Activation of peroxisome proliferator-activated receptor gamma bypasses the function of the retinoblastoma protein in adipocyte differentiation. J. Biol. Chem. 1999, 274, 2386–2393. [Google Scholar] [CrossRef] [Green Version]
- Prusty, D.; Park, B.H.; Davis, K.E.; Farmer, S.R. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor gamma (PPARgamma) and C/EBPalpha gene expression during the differentiation of 3T3-L1 preadipocytes. J. Biol. Chem. 2002, 277, 46226–46232. [Google Scholar] [CrossRef] [Green Version]
- Fève, B. Adipogenesis: Cellular and molecular aspects. Best Pract. Res. Clin. Endocrinol. Metab. 2005, 19, 483–499. [Google Scholar] [CrossRef]
- Kuri-Harcuch, W.; Velez-delValle, C.; Vazquez-Sandoval, A.; Hernández-Mosqueira, C.; Fernandez-Sanchez, V. A cellular perspective of adipogenesis transcriptional regulation. J. Cell. Physiol. 2019, 234, 1111–1129. [Google Scholar] [CrossRef]
- Lowe, C.E.; O’Rahilly, S.; Rochford, J.J. Adipogenesis at a glance. J. Cell Sci. 2011, 124 Pt 16, 2681–2686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salma, N.; Xiao, H.; Imbalzano, A.N. Temporal recruitment of CCAAT/enhancer-binding proteins to early and late adipogenic promoters in vivo. J. Mol. Endocrinol. 2006, 36, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.; Naderi-Manesh, H.; Vali, H.; Eslaminejad, M.B.; Sayahpour, F.A.; Sheibani, S.; Faghihi, S. Contribution of osteocalcin-mimetic peptide enhances osteogenic activity and extracellular matrix mineralization of human osteoblast-like cells. Colloids Surf. B Biointerfaces 2019, 173, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, R.E.B.; Mazurek, M.S.; Soos, A.; Simmons, C.A. Mesenchymal Stromal/Stem Cells in Regenerative Medicine and Tissue Engineering. Stem Cells Int. 2018, 2018, 8031718. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Rey, F.; Lesma, E.; Massihnia, D.; Ciusani, E.; Nava, S.; Vasco, C.; al Haj, G.; Ghilardi, G.; Opocher, E.; Gorio, A.; et al. Adipose-Derived Stem Cells from Fat Tissue of Breast Cancer Microenvironment Present Altered Adipogenic Differentiation Capabilities. Stem Cells Int. 2019, 2019, 1480314. [Google Scholar] [CrossRef] [Green Version]
- Al-Lahham, S.H.; Peppelenbosch, M.P.; Roelofsen, H.; Vonk, R.J.; Venema, K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 2010, 1801, 1175–1183. [Google Scholar] [CrossRef]
- Byrne, C.S.; Chambers, E.S.; Morrison, D.J.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [Green Version]
- Heimann, E.; Nyman, M.; Degerman, E. Propionic acid and butyric acid inhibit lipolysis and de novo lipogenesis and increase insulin-stimulated glucose uptake in primary rat adipocytes. Adipocyte 2015, 4, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, N.R.; Cong, Z.; Nelson, A.M.; Albert, A.J.; Rosamilia, L.L.; Sivarajah, S.; Gilliland, K.L.; Liu, W.; Mauger, D.T.; Gabbay, R.A.; et al. Peroxisome proliferator-activated receptors increase human sebum production. J. Investig. Dermatol. 2006, 126, 2002–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramot, Y.; Mastrofrancesco, A.; Herczeg-Lisztes, E.; Bíró, T.; Picardo, M.; Kloepper, J.E.; Paus, R. Advanced Inhibition of Undesired Human Hair Growth by PPARγ Modulation? J. Investig. Dermatol. 2014, 134, 1128–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrofrancesco, A.; Ottaviani, M.; Cardinali, G.; Flori, E.; Briganti, S.; Ludovici, M.; Zouboulis, C.C.; Lora, V.; Camera, E.; Picardo, M. Pharmacological PPARγ modulation regulates sebogenesis and inflammation in SZ95 human sebocytes. Biochem. Pharmacol. 2017, 138, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Rosenfield, R.L.; Kentsis, A.; Deplewski, D.; Ciletti, N. Rat preputial sebocyte differentiation involves peroxisome proliferator-activated receptors. J. Investig. Dermatol. 1999, 112, 226–232. [Google Scholar] [CrossRef]
- Kawaji, A.; Ohnaka, Y.; Osada, S.; Nishizuka, M.; Imagawa, M. Gelsolin, an actin regulatory protein, is required for differentiation of mouse 3T3-L1 cells into adipocytes. Biol. Pharm. Bull. 2010, 33, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, A.S.; Shen, W.J.; Muliro, K.; Patel, S.; Souza, S.C.; Roth, R.A.; Kraemer, F.B. Stimulation of lipolysis and hormone-sensitive lipase via the extracellular signal-regulated kinase pathway. J. Biol. Chem. 2001, 276, 45456–45461. [Google Scholar] [CrossRef] [Green Version]
- Su, T.Z.; Wang, M.; Oxender, D.L.; Saltiel, A.R. Troglitazone increases system a amino acid transport in 3T3-L1 cells. Endocrinology 1998, 139, 832–837. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, O.H.; Lee, B.Y. Fucoidan, a sulfated polysaccharide, inhibits adipogenesis through the mitogen-activated protein kinase pathway in 3T3-L1 preadipocytes. Life Sci. 2010, 86, 791–797. [Google Scholar] [CrossRef]
- Tafuri, S.R. Troglitazone enhances differentiation, basal glucose uptake, and Glut1 protein levels in 3T3-L1 adipocytes. Endocrinology 1996, 137, 4706–4712. [Google Scholar] [CrossRef] [Green Version]
- Carelli, S.; Messaggio, F.; Canazza, A.; Hebda, D.M.; Caremoli, F.; Latorre, E.; Grimoldi, M.G.; Colli, M.; Bulfamante, G.; Tremolada, C.; et al. Characteristics and Properties of Mesenchymal Stem Cells Derived From Microfragmented Adipose Tissue. Cell Transplant 2015, 24, 1233–1252. [Google Scholar] [CrossRef] [Green Version]
- Farmer, S.R. Regulation of PPARgamma activity during adipogenesis. Int. J. Obes. 2005, 29 (Suppl. 1), S13–S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sears, A.H.D.D.; Ofrecio, J.M.; Chapman, J.; He, W.; Olefsky, J.M. selective modulation of promoter recruitment and transcriptional activity of PPARgamma. Biochem Biophys Res Commun. 2007, 364, 515–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, L.S.; Depaoli, A.M. Selective peroxisome proliferator-activated receptor gamma (PPARgamma) modulation as a strategy for safer therapeutic PPARgamma activation. Am. J. Clin. Nutr. 2010, 91, 267S–272S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirat, C.; Farce, A.; Lebègue, N.; Renault, N.; Furman, C.; Millet, R.; Yous, S.; Speca, S.; Berthelot, P.; Desreumaux, P.; et al. Targeting peroxisome proliferator-activated receptors (PPARs): Development of modulators. J. Med. Chem. 2012, 55, 4027–4061. [Google Scholar] [CrossRef]
- Downie, M.M.; Guy, R.; Kealey, T. Advances in sebaceous gland research: Potential new approaches to acne management. Int. J. Cosmet. Sci. 2004, 26, 291–311. [Google Scholar] [CrossRef]
- Coder, D.M. Assessment of cell viability. Curr Protoc. Cytom. 2001, 15, 9.2.1–9.2.14. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| C/EBPβ | CGCCTACCTGGGCTACCA | GACAGCTGCTCCACCTTCTTC |
| C/EBPδ | ATACCTCAGACCCCGACAGC | ATGCTTTCCCGTGTTCCTTC |
| C/EBPα | GAAGGTGCTGGAGTTGACCA | AGGAAGCAGGAATCCTCCAA |
| PPARγ | GTGGGGATGTCTCACAATGC | TGATCTCTTGCACGGCTTCT |
| FABP-4 | ACGGCCCTGCAGAACTATCT | AAGGTTCACAAACGCGACAG |
| leptin | TGTGCACCTGAGGGTAGAGG | CCCTGGACAACCTTGGAGAT |
| Hsl | GCTTCTCCCTCTCGTCTGCT | CAGACACACTCCTGCGCATA |
| 18S rRNA | ACCGCGGTTCTATTTTGTTG | GACAAATCGCTCCACCAACT |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| PPARγ | CAAGAGTACCAAAGTGCAATCAAAGTGGAG | GTTCTCCGGAAGAAACCCTTGCATCCTTCA |
| FABP-4 | CTGGGCCAGGAATTTGACGA | ACCAGGACACCCCATCTAA |
| GAPDH | CTTTTGCGTCGCCAG | TTGATGGCAACAATATCCAC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Haj, G.; Rey, F.; Giallongo, T.; Colli, M.; Marzani, B.; Giuliani, G.; Gorio, A.; Zuccotti, G.V.; Di Giulio, A.M.; Carelli, S. A New Selective PPARγ Modulator Inhibits Triglycerides Accumulation during Murine Adipocytes’ and Human Adipose-Derived Mesenchymal Stem Cells Differentiation. Int. J. Mol. Sci. 2020, 21, 4415. https://doi.org/10.3390/ijms21124415
Al Haj G, Rey F, Giallongo T, Colli M, Marzani B, Giuliani G, Gorio A, Zuccotti GV, Di Giulio AM, Carelli S. A New Selective PPARγ Modulator Inhibits Triglycerides Accumulation during Murine Adipocytes’ and Human Adipose-Derived Mesenchymal Stem Cells Differentiation. International Journal of Molecular Sciences. 2020; 21(12):4415. https://doi.org/10.3390/ijms21124415
Chicago/Turabian StyleAl Haj, Ghina, Federica Rey, Toniella Giallongo, Mattia Colli, Barbara Marzani, Giammaria Giuliani, Alfredo Gorio, Gian Vicenzo Zuccotti, Anna Maria Di Giulio, and Stephana Carelli. 2020. "A New Selective PPARγ Modulator Inhibits Triglycerides Accumulation during Murine Adipocytes’ and Human Adipose-Derived Mesenchymal Stem Cells Differentiation" International Journal of Molecular Sciences 21, no. 12: 4415. https://doi.org/10.3390/ijms21124415
APA StyleAl Haj, G., Rey, F., Giallongo, T., Colli, M., Marzani, B., Giuliani, G., Gorio, A., Zuccotti, G. V., Di Giulio, A. M., & Carelli, S. (2020). A New Selective PPARγ Modulator Inhibits Triglycerides Accumulation during Murine Adipocytes’ and Human Adipose-Derived Mesenchymal Stem Cells Differentiation. International Journal of Molecular Sciences, 21(12), 4415. https://doi.org/10.3390/ijms21124415