ASGR1 and Its Enigmatic Relative, CLEC10A

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. ASGR1
3. Targeting Liver Cells via ASGR1
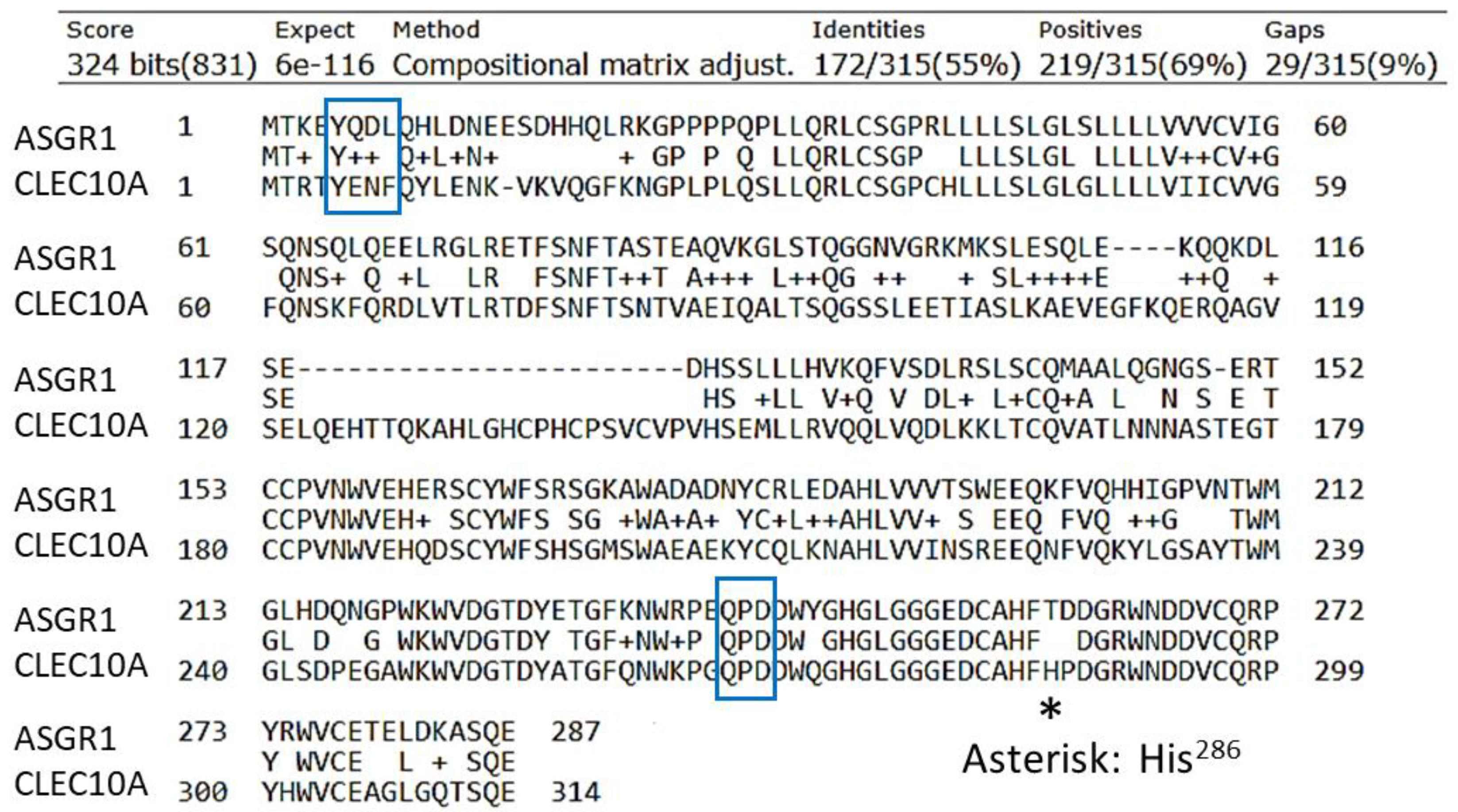
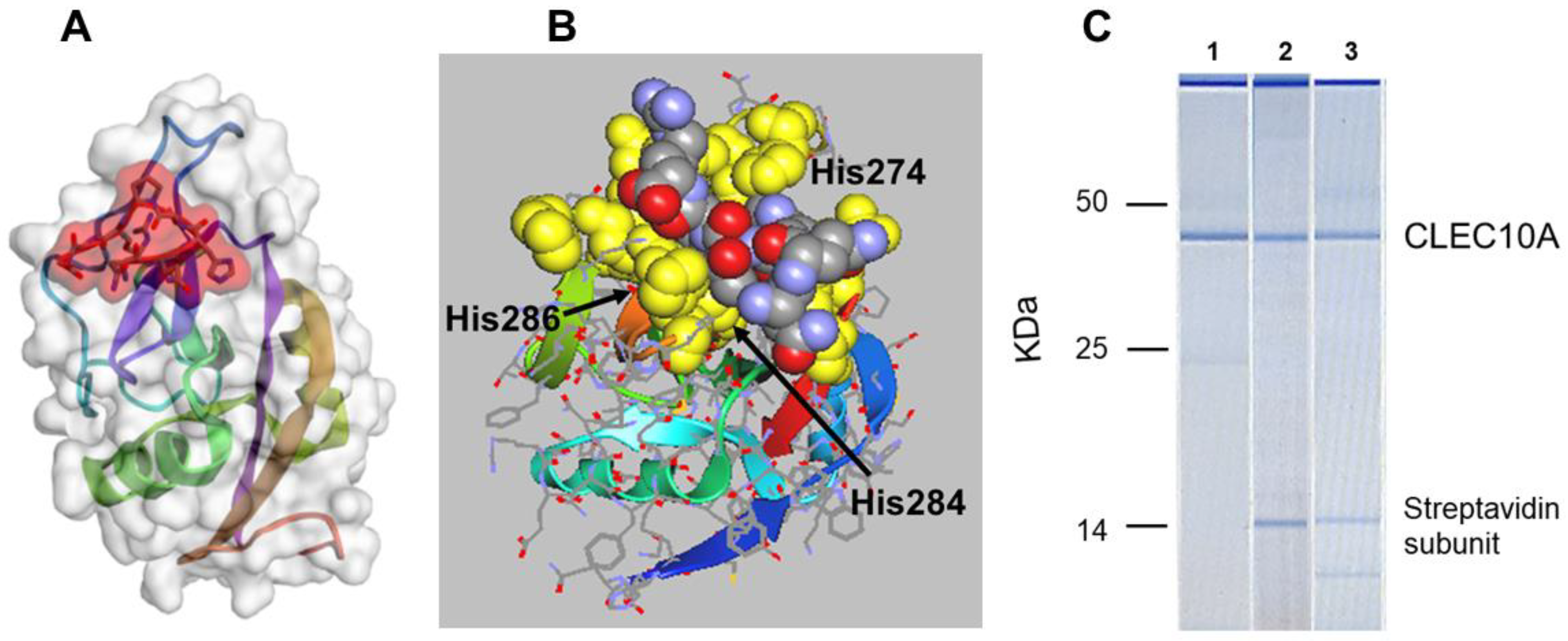
4. CLEC10A
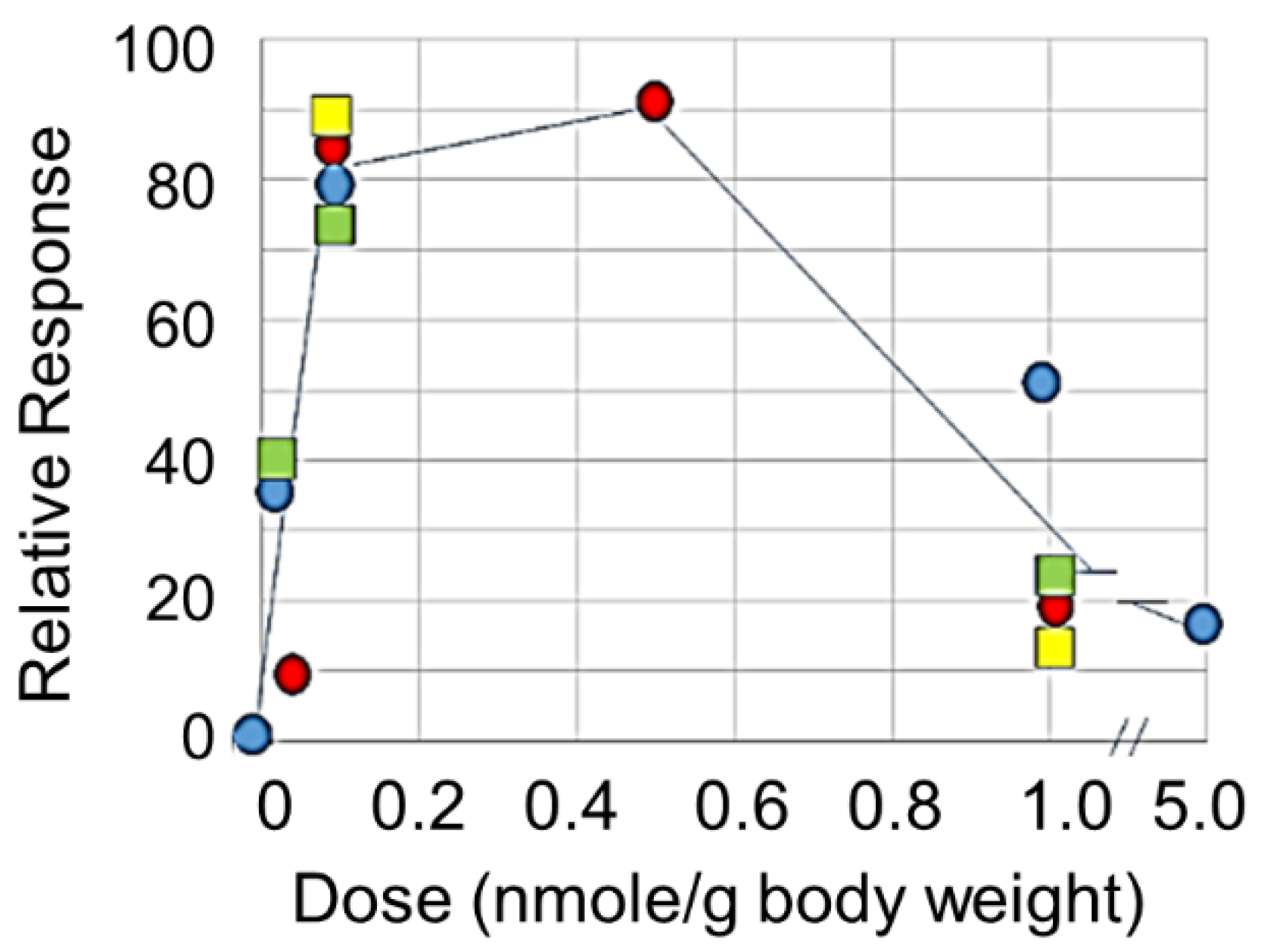
5. A Ligand Mimetic
6. Roles for CLEC10A in Health and Disease
7. Activation of T Cells
8. The Ca2+ Connection
9. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Pillai, S.; Netravali, I.A.; Cariappa, A.; Mattoo, H. Siglecs and Immune Regulation. Annu. Rev. Immunol. 2012, 30, 357–392. [Google Scholar] [CrossRef] [Green Version]
- Macauley, M.S.; Crocker, P.R.; Paulson, J.C. Siglec-Mediated Regulation of Immune Cell Function in Disease. Nat. Rev. Immunol. 2014, 14, 653–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, S.; Paulson, J.C. Siglecs as Immune Cell Checkpoints in Disease. Annu. Rev. Immunol. 2020, 38, 365–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabinovich, G.A.; Toscano, M.A. Turning ‘Sweet’ on Immunity: Galectin-Glycan Interactions in Immune Tolerance and Inflammation. Nat. Rev. Immunol. 2009, 9, 338–352. [Google Scholar] [CrossRef]
- Di Lella, S.; Sundblad, V.; Cerliani, J.P.; Guardia, C.M.; Estrin, D.A.; Vasta, G.R.; Rabinovich, G.A. When Galectins Recognize Glycans: From Biochemistry to Physiology and Back Again. Biochemistry 2011, 50, 7842–7857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasta, G.R.; Amzel, L.M.; Bianchet, M.A.; Cammarata, M.; Feng, C.; Saito, K. F-Type Lectins: A Highly Diversified Family of Fucose-Binding Proteins with a Unique Sequence Motif and Structural Fold, Involved in Self/Non-Self-Recognition. Front. Immunol. 2017, 8, 1648. [Google Scholar] [CrossRef]
- Cummings, R.D.; McEver, R.P. Chapter 31: R-Type Lectins. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017; pp. 2015–2017. [Google Scholar]
- Drickamer, K.; Taylor, M.E. Recent Insights into Structures and Functions of C-Type Lectins in the Immune System. Curr. Opin. Struct. Biol. 2015, 34, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.E.; Drickamer, K. Mammalian Sugar-Binding Receptors: Known Functions and Unexplored Roles. FEBS J. 2019, 286, 1800–1814. [Google Scholar] [CrossRef]
- Sancho, D.; Reis e Sousa, C. Signaling by Myeloid C-Type Lectin Receptors in Immunity and Homeostasis. Ann. Rev. Immunol. 2012, 30, 491–529. [Google Scholar] [CrossRef] [Green Version]
- Drickamer, K. Two Distinct Classes of Carbohydrate-recognition Domains in Animal Lectins. J. Biol. Chem. 1988, 263, 9557–9560. [Google Scholar]
- Weis, W.I.; Taylor, M.E.; Drickamer, K. The C-Type Lectin Superfamily in the Immune System. Immunol. Rev. 1998, 163, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Kanemaru, K.; Noguchi, E.; Tahara-Hanaoka, S.; Mizuno, S.; Tateno, H.; Denda-Nagai, K.; Irimura, T.; Matsuda, H.; Sugiyama, F.; Takahashi, S.; et al. Clec10a Regulates Mite-Induced Dermatitis. Sci. Immunol. 2019, 4, eaax6908. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.L.; van den Berg, C.W.; Bowen, D.J. ASGR1 and ASGR2, the Genes that Encode the Asialoglycoprotein Receptor (Ashwell Receptor), Are Expressed in Peripheral Blood Monocytes and Show Interindividual Differences in Transcript Profile. Mol. Biol. int. 2012, 2012, 283974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UniProt.org. UniProtKB-Q8IUN9(CLC10_HUMAN). Available online: www.uniprot.org (accessed on 25 May 2020).
- Vlismas, A.; Bletsa, R.; Mavrogianni, D.; Mamali, G.; Pergamali, M.; Dinopoulo, V.; Partsinevelos, G.; Drakakis, P.; Loutradis, D.; Kiessling, A.A. Microarray Analyses Reveal Marked Differences in Growth Factor and Receptor Expression Between 8-Cell Human Embryos and Pluripotent Stem Cells. Stem Cells Dev. 2016, 25, 160–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimmeck, D.; Hansson, J.; Raffel, S.; Vakhrushev, S.Y.; Trumpp, A.; Krijgsveld, J. Proteomic Cornerstones of Hematopoietic Stem Cell Differentiation: Distinct Signatures of Multipotent Progenitors and Myeloid Committed Cells. Mol. Cell. Proteom. 2012, 11, 286–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Kooyk, Y.; Ilarregui, J.M.; van Vliet, S.J. Novel Insights into the Immunomodulatory Role of the Dendritic Cell and Macrophage-expressed C-type Lectin MGL. Immunobiology 2015, 220, 185–192. [Google Scholar] [CrossRef]
- Napoletano, C.; Rughetti, A.; Tarp, M.P.A.; Coleman, J.; Bennett, E.P.; Picco, G.; Sale, P.; Denda-Nagai, K.; Irimura, T.; Mandel, U.; et al. Tumor-Associated Tn-MUC1 Glycform is Internalized through the Macrophage Galactose-type C-type Lectin and Delivered to the HLA Class I and II Compartments in Dendritic Cells. Cancer Res. 2007, 67, 8358–8367. [Google Scholar] [CrossRef] [Green Version]
- Tang-Huau, T.L.; Gueguen, P.; Goudot, C.; Durand, M.; Bohec, M.; Baulande, S.; Pasquier, B.; Amigorena, S.; Sequra, E. Human in vivo-Generated Monocyte-derived Dendritic Cells and Macrophages Cross-present Antigens through a Vacuolar Pathway. Nat. Commun. 2018, 9, 2570. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, W.; Xiong, S. Macrophage Differentiation and Polarization via Phosphatidylinositol 3-Kinase/Akt-ERK Signaling Pathway Conferred by Serum Amyloid P Component. J. Immunol. 2011, 187, 1764–1777. [Google Scholar] [CrossRef] [Green Version]
- Röszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.L.; Tai, J.J.-Y.; Wong, W.-C.; Han, H.; Sem, X.; Yeap, W.-H.; Kourilsky, P.; Wong, S.-C. Gene Expression Profiling Reveals the Defining Features of the Classical, Intermediate, and Nonclassical Human Monocyte Subsets. Blood 2011, 118, e16–e31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapoport, E.M.; Moiseeva, E.V.; Aronov, D.A.; Khaidukov, S.V.; Pazynina, G.V.; Tsygankova, S.V.; Ryzhov, I.M.; Belyanchikov, I.M.; Tyrtysh, T.V.; McCullough, K.C.; et al. Glycan-Binding Profile of DC-like Cells. Glycoconj. J. 2020, 37, 129–138. [Google Scholar] [CrossRef]
- Napoletano, C.; Zizzari, I.G.; Rughetti, A.; Rahimi, H.; Irimura, T.; Clausen, H.; Wandall, H.H.; Belleudi, F.; Bellati, F.; Pierelli, L.; et al. Targeting of Macrophage Galactose-type C-type Lectin (MGL) Induces DC Signaling and Activation. Eur. J. Immunol. 2012, 42, 936–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zizzari, I.G.; Napoletano, C.; Battisti, F.; Rahimi, H.; Caponnetto, S.; Pierelli, L.; Nuti, M.; Rughetti, A. MGL Receptor and Immunity: When the Ligand can Make the Difference. J. Immunol. Res. 2015, 2015, 450695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morell, A.G.; Irvine, R.A.; Sternlieb, I.; Scheinberg, I.H.; Ashwell, G. Physical and Chemical Studies on Ceruloplasmin. V. Metabolic Studies on Sialic Acid-Free Ceruloplasmin In Vivo. J. Biol. Chem. 1968, 243, 155–159. [Google Scholar]
- Morell, A.G.; Gregoriadis, G.; Scheinberg, I.H.; Hickman, J.; Ashwell, G. The Role of Sialic Acid in Determining the Survival of Glycoproteins in the Circulation. J. Biol. Chem. 1971, 246, 1461–1467. [Google Scholar] [PubMed]
- Pricer, W.E., Jr.; Ashwell, G. The Binding of Desialylated Glycoproteins by Plasma Membranes of Rat Liver. J. Biol. Chem. 1971, 246, 4825–4833. [Google Scholar] [PubMed]
- Sarkar, M.; Liao, J.; Kabat, E.A.; Tanabe, T.; Ashwell, G. The Binding Site of Rabbit Hepatic Lectin. J. Biol. Chem. 1979, 254, 3170–3174. [Google Scholar]
- Steer, C.J.; Ashwell, G. Studies on a Mammalian Hepatic Binding Protein Specific for Asialoglycoproteins. Evidence for Receptor Recycling in Isolated Hepatocytes. J. Biol. Chem. 1980, 255, 3008–3013. [Google Scholar]
- Schönfeld, D.L.; Ravelli, R.B.G.; Mueller, U.; Skerra, A. The 1.8-Å Crystal Structure of α1-Acid Glycoprotein (Orosomucoid) Solved by UV RIP Reveals the Broad Drug-binding Activity of This Human Plasma Lipocalin. J. Mol. Biol. 2008, 384, 393–405. [Google Scholar] [CrossRef]
- Nishi, K.; Ono, T.; Nakamura, T.; Fukunaga, N.; Izumi, M.; Watanabe, H.; Senaga, A.; Maruyama, T.; Yamagata, Y.; Curry, S.; et al. Structural Insights into Differences in Drug-binding Selectivity between Two Forms of Human α1-Acid Glycoprotein Genetic Variants, the A and F1*S Forms. J. Biol. Chem. 2011, 286, 14427–14434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, H.; Torgersen, D.; Drickamer, K.; Weis, W.I. Mechanism of pH-Dependent N-Acetylgalactosamine Binding by a Functional Mimic of the Hepatic Asialoglycoprotein Receptor. J. Biol. Chem. 2000, 275, 35176–35184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, N.I.; Drikamer, K. Differential Ligand Binding by Two Subunits of the Rat Liver Asialoglycoprotein Receptor. Glycobiology 1996, 6, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Henis, Y.I.; Katzir, Z.; Shia, M.A.; Lodish, H.F. Oligomeric Structure of the Human Asialoglycoprotein Receptor: Nature and Stoichiometry of Mutual Complexes Containing H1 and H2 Polypeptides Assessed by Fluorescence Photobleaching Recovery. J. Cell Biol. 1990, 111, 1409–1418. [Google Scholar] [CrossRef]
- Bider, M.D.; Wahlberg, J.M.; Kammerer, R.A.; Spiess, M. The Oligomerization Domain of the Asialoglycoprotein Receptor Preferentially Forms 2:2 Heterotetramers In Vitro. J. Biol. Chem. 1996, 271, 31996–32001. [Google Scholar] [CrossRef] [Green Version]
- Hardy, M.R.; Townsend, R.R.; Parkhurst, S.M.; Lee, Y.C. Different Modes of Ligand Binding to the Hepatic Galactose/N-Acetylgalactosamine Lectin on the Surface of Rabbit Hepatocytes. Biochemistry 1985, 24, 22–28. [Google Scholar] [CrossRef]
- Schwartz, A.L.; Rup, D.; Lodish, H.F. Difficulties in the Quantitation of Asialoglycoprotein Receptors on the Rat Hepatocyte. J. Biol. Chem. 1980, 255, 9033–9036. [Google Scholar]
- Schwartz, A.L.; Marshak-Rothstein, A.; Rup, D.; Lodish, H.F. Identification and Quantitation of the Rat Hepatocyte Asialoglycoprotein Receptor. Proc. Natl. Acad. Sci. USA 1981, 78, 3348–3352. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, A.L.; Fridovich, S.E.; Lodish, H.F. Kinetics of Internalization and Recycling of the Asialoglycoprotein Receptor in a Hepatoma Cell Line. J. Biol. Chem. 1982, 257, 4230–4237. [Google Scholar]
- Geuze, H.J.; Slot, J.W.; Strous, G.J.A.; Lodish, H.F.; Schwartz, A.L. Intracellular Site of Asialoglycoprotein Receptor-Ligand Uncoupling: Double-Label Immunoelectron Microscopy during Receptor-Mediated Endocytosis. Cell 1983, 32, 277–287. [Google Scholar] [CrossRef]
- Ciechanover, A.; Schwartz, A.L.; Lodish, H.F. Sorting and Recycling of Cell Surface Receptors and Endocytosed Ligands: The Asialoglycoprotein and Transferrin Receptors. J. Cell. Biochem. 1983, 23, 107–130. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.L.; Bolognesi, A.; Fridovich, S.E. Recycling of the Asialoglycoprotein Receptor and the Effect of Lysosomotropic Amines in Hepatoma Cells. J. Cell Biol. 1984, 98, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Mettlen, M.; Chen, P.-H.; Srinivasan, S.; Danuser, G.; Schmid, S.L. Regulation of Clathrin-Mediated Endocytosis. Annu. Rev. Biochem. 2018, 87, 871–896. [Google Scholar] [CrossRef] [PubMed]
- Meier, M.; Bider, M.D.; Malashkevich, V.N.; Spiess, M.; Burkhard, P. Crystal Structure of the Carbohydrate Recognition Domain of the H1 Subunit of the Asialoglycoprotein Receptor. J. Mol. Biol. 2000, 300, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Gerasimenko, J.V.; Tepikin, A.V.; Petersen, O.H.; Gerasimenko, O.V. Calcium uptake via endocytosis with rapid release from acidifying endosomes. Curr. Biol. 1998, 8, 1335–1338. [Google Scholar] [CrossRef] [Green Version]
- Onizuka, T.; Shimizu, H.; Moriwaki, Y.; Nakano, T.; Kanai, S.; Shimada, I.; Takahashi, H. NMR Study of Ligand Release from Asialoglycoprotein Receptor Under Solution Conditions in Early Endosomes. FEBS J. 2012, 279, 2645–2656. [Google Scholar] [CrossRef]
- Maxson, M.E.; Grinstein, S. The Vacuolar-type H+-ATPase at a Glance—More than a Proton Pump. J. Cell Sci. 2014, 127, 4987–4993. [Google Scholar] [CrossRef] [Green Version]
- Pamarthy, S.; Kulshrestha, A.; Katara, G.K.; Beaman, K.D. The Curious Case of Vacuolar ATPase: Regulation of Signaling Pathways. Mol. Cancer 2018, 17, 41. [Google Scholar] [CrossRef]
- Morgan, A.J.; Platt, F.M.; Lloyd-Evans, E.; Galione, A. Molecular Mechanisms of Endolysosomal Ca2+ Signalling in Health and Disease. Biochem. J. 2011, 439, 349–374. [Google Scholar] [CrossRef] [Green Version]
- Shumilina, E.; Huber, S.M.; Lang, F. Ca2+ Signaling in the Regulation of Dendritic Cell Functions. Am. J. Physiol. Cell. Physiol. 2011, 300, C1205–C1214. [Google Scholar] [CrossRef] [Green Version]
- Plattner, H.; Verkhratsky, A. Inseparable Tanden: Evolution Chooses ATP and Ca2+ to Control life, Death and Cellular Signlling. Philos. Trans. R. Soc. B 2016, 371, 20150419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.T.; Lee, Y.C. Facile Synthesis of a High-Affinity Ligand for Mammalian Hepatic Lectin Containing Three Terminal N-Acetylgalactosamine Residues. Bioconj. Chem. 1997, 8, 762–765. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, J.; Gleinich, A.S.; Zhang, Q.; Whitfeld, R.; Kempe, K.; Haddleton, D.M.; Davis, T.P.; Perrier, S.; Mitchell, D.A.; Wilson, P. Specific and Differential Binding of N-Acetylgalactosamine Glycopolymers to the Human Macrophage Galactose Lectin and Asialoglycoprotein Receptor. Biomolecules 2017, 18, 1624–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerlind, U.; Westman, J.; Törnquist, E.; Smith, C.I.E.; Oscarson, S.; Lahmann, M.; Norberg, T. Ligands of the Asialoglycoprotein Receptor for Targeted Gene Delivery, Part 1: Synthesis of and Binding Studies with Biotinylated Cluster Glycosides Containing N-Acetylgalactosamine. Glycoconj. J. 2004, 21, 227–241. [Google Scholar] [CrossRef]
- Rensen, P.C.N.; van Leeuwen, S.H.; Sliedregt, L.A.J.M.; van Berkel, T.J.C.; Biessen, E.A.L. Design and Synthesis of Novel N-Acetylgalactosamine-Terminated Glycolipids for Targeting of Lipoproteins to the Hepatic Asialoglyoprotein Receptor. J. Med. Chem. 2004, 47, 5798–5808. [Google Scholar] [CrossRef]
- Lepenies, B.; Lee, J.; Sonkaria, S. Targeting C-Type Lectin Receptors with Multivalent Carbohydrate Ligands. Adv. Drug Del. Rev. 2013, 65, 1271–1281. [Google Scholar] [CrossRef]
- Khorev, O.; Stokmaier, D.; Schwardt, O.; Cutting, B.; Ernst, B. Trivalent, Gal/GalNAc-Containing Ligands Designed for the Asialoglycoprotein Receptor. Bioorg. Med. Chem. 2008, 16, 5216–5231. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, X.; Liu, Y. Asialoglycoprotein Receptor-Targeted Radiopharmaceuticals for Measurement of Liver Function. Curr. Med. Chem. 2014, 21, 4–23. [Google Scholar] [CrossRef]
- Abe, M.; Lai, J.; Kortylewicz, Z.P.; Nagata, H.; Fox, I.J.; Enke, C.A.; Baranowska-Kortylewicz, J. Radiolabeled Constructs for Evaluation of the Asialoglycoprotein Receptor Status and Hepatic Functional Reserves. Bioconj. Chem. 2003, 14, 997–1006. [Google Scholar] [CrossRef]
- Prakash, T.P.; Yu, J.; Migawa, M.T.; Kinberger, G.A.; Wan, W.B.; Østergaard, M.E.; Vasquez, G.; Low, A.; Chappell, A.; Schmidt, K. Comprehensive Structure-Activity Relationship of Triantennary N-Acetylgalactosamine Conjugated Antisense Oligonucleotides for Targeted Delivery to Hepatocytes. J. Med. Chem. 2016, 59, 2718–2733. [Google Scholar] [CrossRef]
- Yamamoto, T.; Sawamura, M.; Wada, F.; Harada-Shiba, M.; Obika, S. Serial Incorporation of a Monovalent GalNAc Phosphoramidite Unit into Hepatocyte-Targeting Antisense Oligonucleotides. Bioorg. Med. Chem. 2016, 24, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Nair, J.K.; Willoughby, J.L.S.; Chan, A.; Charisse, K.; Alam, M.R.; Wang, Q.; Hoekstra, M.; Kandasamy, P.; Kel’in, A.V.; Milstein, S.; et al. Multivalent N-Acetylgalactosamine-Conjugated siRNA Localizes in Hepatocytes and Elicits Robust RNAi-Mediated Gene Silencing. J. Am. Chem. Soc. 2014, 136, 16958–16961. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y. Preclinical and Clinical Advances of GalNAc-Decorated Nucleic Acid Therapeutics. Mol. Ther. Nucleic Acids 2017, 6, 116–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinberger, G.A.; Prakash, T.P.; Yu, J.; Vasquez, G.; Low, A.; Chappell, A.; Schmidt, K.; Murray, H.M.; Gaus, H.; Swayze, E.E.; et al. Conjugation of Mono- and Di-GalNAc Sugars Enhances the Potency of Antisense Oligonucleotides via ASGR Mediated Delivery to Hepatocytes. Bioorg. Med. Chem. Lett. 2016, 26, 3690–3693. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, R.Z.; Henry, S.; Geary, R.S. Pharmacokinetics and Clinical Pharmacology Considerations of GalNAc3-Conjugated Antisense Oligonucleotides. Exp. Opin. Drug Metab. Toxicol. 2019, 15, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.T.; Baker, B.F.; Zia, S.; Yu, R.Z.; Viney, N.J.; Wang, Y.; Tsimikas, S.; Geary, R.S. Integrated Assessment of the Clinical Performance of GalNAc3-Conjugated 2′-O-Methoxyethyl Chimeric Antisense Oligonucleotides. 1. Human Volunteer Experience. Nucleic Acid Ther. 2019, 29, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Guan, H.; McGuire, M.J.; Li, S.; Brown, K.C. Peptide-Targeted Polyglutamic Acid Doxorubicin Conjugates for the Treatment of αvβ6-Positive Cancers. Bioconjug. Chem. 2008, 19, 1813–1821. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Wang, Y.; Ling, X.; Chaurasiya, B.; Yang, C.; Du, Y.; Tu, J.; Xiong, Y.; Sun, C. Efficient Delivery of Paclitaxel into ASGPR Over-expressed Cancer Cells Using Reversibly Stabilized Multifunctional Pullulan Nanoparticles. Carbohydr. Polym. 2017, 159, 178–187. [Google Scholar] [CrossRef]
- Pranatharthiharan, S.; Patel, M.D.; Malshe, V.C.; Pujari, V.; Gorakshakar, A.; Madkaikar, M.; Ghosh, K.; Devarajan, P.V. Asialoglycoprotein Receptor Targeted Delivery of Doxorubicin Nanoparticles for Hepatocellular Carcinoma. Drug Deliv. 2017, 24, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Gibadullin, R.; Farnsworth, D.W.; Barchi, J.J., Jr.; Gildersleeve, J.C. GalNAc-Tyrosine is a Ligand of Plant Lectins, Antibodies, and Human and Murine Macrophage Galactose-Type Lectins. ACS Chem. Biol. 2017, 12, 2172–2182. [Google Scholar] [CrossRef]
- Rensen, P.C.N.; Sliedregt, L.A.J.M.; Ferns, M.; Kieviet, E.; van Rossenberg, S.M.W.; van Leeuwen, S.H.; van Berkel, T.J.C.; Biessen, E.A.L. Determination of the Upper Size Limit for Uptake and Processing of Ligands by the Asialoglycoprotein Receptor on Hepatocytes In Vitro and In Vivo. J. Biol. Chem. 2001, 276, 37577–37584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ii, M.; Kurata, H.; Itoh, N.; Yamashina, I.; Kawasaki, T. Molecular Cloning and Sequence Analysis of cDNA Encoding the Macrophage Lectin Specific for Galactose and N-Acetylgalactosamine. J. Biol. Chem. 1990, 265, 11295–11298. [Google Scholar] [PubMed]
- Sato, M.; Kawakami, K.; Osawa, T.; Toyoshima, S. Molecular Cloning and Expression of cDNA Encoding a Galactose/N-Acetylgalactosamine-Specific Lectin on Mouse Tumoricidal Macrophages. J. Biochem. 1992, 111, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Tsuiji, M.; Fujimori, M.; Ohashi, Y.; Higashi, N.; Onami, T.M.; Hedrick, S.M.; Irimura, T. Molecular Cloning and Characterization of a Novel Mouse Macrophage C-type Lectin, mMGL2, Which Has a Distinct Carbohydrate Specificity from mMGL1. J. Biol. Chem. 2002, 277, 28892–28901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denda-Nagai, K.; Aida, A.; Saba, K.; Suzuki, K.; Moriyama, S.; Oo-puthinan, S.; Tsuiji, M.; Morikawa, A.; Kumamoto, Y.; Sugiura, D.; et al. Distribution and Function of Macrophage Galactose-type C-type Lectin 2 (MGL2/CD301b). J. Biol. Chem. 2010, 285, 19193–19204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Yamamoto, K.; Toyoshima, S.; Osawa, T.; Irimura, T. Molecular Cloning and Expression of cDNA Encoding Human Macrophage C-Type Lectin. Its Unique Carbohydrate Binding Specificity for the Tn Antigen. J. Immunol. 1996, 156, 128–135. [Google Scholar]
- Higashi, N.; Fujioka, K.; Denda-Nagai, K.; Hashimoto, S.-I.; Nagai, S.; Sato, T.; Fujita, Y.; Morikawa, A.; Tsuiji, M.; Miyata-Takeuchi, M.; et al. The Macrophage C-type Lectin Specific for Galactose/N-Acetylgalactosamine Is an Endocytic Receptor Expressed on Monocyte-derived Immature Dendritic Cells. J. Biol. Chem. 2002, 277, 20686–20693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artigas, G.; Monteiro, J.T.; Hinou, H.; Nishimura, S.I.; Lepenies, B.; Garcia-Martin, F. Glycopeptides as Targets for Dendritic Cells: Exploring MUC1 Glycopeotides Binding Profile toward Macrophage Galactose-Type Lectin (MGL) Orthologs. J. Med. Chem. 2017, 60, 9012–9021. [Google Scholar] [CrossRef]
- Valladeau, J.; Duvert-Frances, V.; Pin, J.-J.; Kleijmeer, M.J.; Ait-Yahia, S.; Ravel, O.; Vincent, C.; Vega, F., Jr.; Helms, A.; Gorman, D.; et al. Immature Human Dendritic Cells Express Asialoglycoprotein Receptor Isoforms for Efficient Receptor-Mediated Endocytosis. J. Immunol. 2001, 167, 5767–5774. [Google Scholar] [CrossRef] [Green Version]
- Drickamer, K. Engineering Galactose-binding Activity into a C-type Mannose-binding Protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef]
- Iobst, S.T.; Drickamer, K. Selective Sugar Binding to the Carbohydrate Recognition Domain of the Rat Hepatic and Macrophage Asialoglycoprotein Receptors. J. Biol. Chem. 1996, 271, 6686–6693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Vliet, S.J.; van Liempt, E.; Saeland, E.; Aarnoudse, C.A.; Appelmelk, B.; Irimura, T.; Geijtenbeek, T.B.H.; Blixt, O.; Alvarez, R.; van Die, I.; et al. Carbohydrate Profiling Reveals a Distinctive Role for the C-Type Lectin MGL in the Recognition of Helminth Parasites and Tumor Antigens by Dendritic Cells. Int. Immunol. 2005, 17, 661–669. [Google Scholar] [CrossRef]
- Iida, S.-I.; Yamamoto, K.; Irimura, T. Interaction of Human Macrophage C-type Lectin with O-linked N-Acetylgalactosamine Residues on Mucin Glycopeptides. J. Biol. Chem. 1999, 274, 10697–10705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortezai, N.; Behnken, H.N.; Kurze, A.-K.; Ludewig, P.; Buck, F.; Meyer, B.; Wagener, C. Tumor-Associated Neu5Ac-Tn and Neu5Gc-Tn Antigens bind to C-Type Lectin CLEC10A (CD301, MGL). Glycobiology 2013, 23, 844–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heger, L.; Balk, S.; Lühr, J.J.; Heidkamp, G.F.; Lehmann, C.H.K.; Hatscher, L.; Purbojo, A.; Hartmann, A.; Garcia-Martin, F.; Nishimura, S.-I.; et al. CLEC10A Is a Specific Marker for Human CD1c+ Dendritic Cells and Enhances Their Toll-Like Receptor 7/8-Induced Cytokine Secretion. Front. Immunol. 2018, 9, 744. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, S.J.; Aarnoudse, C.A.; Broks-van den Berg, V.C.M.; Boks, M.; Geijtenbeek, T.B.H.; van Kooyk, Y. MGL-Mediated Internalization and Antigen Presentation by Dendritic Cells: A Role for Tyrosine-5. Eur. J. Immunol. 2007, 37, 2075–2081. [Google Scholar] [CrossRef]
- Gu, C.; Wang, L.; Zurawski, S.; Oh, S.K. Signaling Cascade through DC-ASGPR Induces Transcriptionally Active CREB for IL-10 Induction and Immune Regulation. J. Immunol. 2019, 203, 389–399. [Google Scholar] [CrossRef]
- Wang, L.-X.; Zhang, S.-X.; Wu, H.-J.; Rong, X.-L.; Guo, J. M2b Macrophage Polarization and Its Roles in Diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Forrester, M.A.; Wassall, H.J.; Hall, L.S.; Cao, H.; Wilson, H.M.; Barker, R.N.; Vickers, M.A. Similarities and Differences in Surface Receptor Expression by THP-1 Monocytes and Differentiated Macrophages Polarized Using Seven Different Conditioning Regimens. Cell. Immunol. 2018, 332, 58–76. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, S.J.; Saeland, E.; van Kooyk, E. Sweet Preferences of MGL: Carbohydrate Specificity and Function. Trends Immunol. 2008, 29, 83–90. [Google Scholar] [CrossRef]
- Cote, R.; Eggink, L.L.; Hoober, J.K. CLEC Receptors, Endocytosis, and Calcium Signaling. AIMS Aller Immunol. 2017, 1, 207–231. [Google Scholar] [CrossRef]
- Villani, A.-C.; Satija, R.; Reynolds, G.; Sarkizova, S.; Shekkar, K.; Fletcher, J.; Griesbeck, M.; Butler, A.; Zheng, S.; Lazo, S.; et al. Single-Cell RNA-seq Reveals New Types of Human Blood Dendritic Cells, Monocytes, and Progenitors. Science 2017, 356, eaah4573. [Google Scholar] [CrossRef] [Green Version]
- Minoda, Y.; Virshup, I.; Rojas, I.L.; Haigh, O.; Wong, Y.; Miles, J.J.; Wells, C.A.; Radford, K.J. Human CD141+ Dendritic Cell and CD1c+ Dendritic Cell Undergo Concordant Early Genetic Programming after Activation in Humanized Mice In Vivo. Front. Immunol. 2017, 8, 1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.G.; Gudjonson, H.; Pritykin, Y.; Deep, D.; Lavalléd, V.-P.; Mendoza, A.; Fromme, R.; Mazutis, L.; Ariyan, C.; Leslie, C.; et al. Transcriptional Basis of Mouse and Human Dendritic Cell Heterogeneity. Cell 2019, 179, 846–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang-Huau, T.-L.; Segura, E. Human In Vivo-Differentiated Monocyte-Derived Dendritic Cells. Sem. Cell Dev. Biol. 2018, 86, 44–49. [Google Scholar] [CrossRef]
- Freire, T.; Zhang, X.; Dériaud, E.; Ganneau, C.; Vichier-Guerre, S.; Azria, E.; Launay, O.; Lo-Man, R.; Bay, S.; Leclerc, C. Glycosidic Tn-based Vaccines Targeting Dermal Dendritic Cells Favor Germinal Center B-Cell Development and Potent Antibody Response in the Absence of Adjuvant. Blood 2010, 116, 3526–3536. [Google Scholar] [CrossRef] [Green Version]
- Murakami, R.; Denda-Nagai, K.; Hashimoto, S.-I.; Nagai, S.; Hattori, M.; Irimura, T. A Unique Dermal Dendritic Cell Subset that Skews the Immune Response Toward Th2. PLoS ONE 2013, 8, e73270. [Google Scholar] [CrossRef]
- Garg, A.; Oran, A.; Wajchman, J.; Sasaki, S.; Maris, C.H.; Kapp, J.A.; Jacob, J. Genetic Tagging Shows Increased Frequency and Longevity of Antigen-presenting, Skin-derived Dendritic Cells In Vivo. Nat. Immunol. 2003, 4, 907–912. [Google Scholar] [CrossRef]
- Tomura, M.; Hata, A.; Matsuoka, S.; Shand, F.H.W.; Nakahishi, Y.; Ikebuchi, R.; Ueha, S.; Tsutsui, H.; Inaba, K.; Matsushima, K.; et al. Tracking and Quantification of Dendritic Cell Migration and Antigen Trafficking Between the Skin and Lymph Nodes. Sci. Rep. 2014, 4, 6030. [Google Scholar] [CrossRef]
- Kitano, M.; Yamazaki, C.; Takumi, A.; Ikeno, T.; Hemmi, H.; Takahashi, N.; Shimizu, K.; Fraser, S.E.; Hoshino, K.; Kaisho, T.; et al. Imaging of the Cross-Presenting Dendritic Cell Subsets in the Skin-Draining Lymph Node. Proc. Natl. Acad. Sci. USA 2016, 113, 1044–1049. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, S.J.; Gringhuis, S.I.; Geijtenbeek, T.B.H.; van Kooyk, Y. Regulation of Effector T Cells by Antigen-Presenting Cells via Interaction of the C-Type Lectin MGL with CD45. Nat. Immunol. 2006, 7, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Mnich, M.E.; van Dalen, R.; Gerlach, D.; Hendriks, A.; Xia, G.; Peschel, A.; van Strijp, J.A.G.; van Sorge, N.M. The C-Type Lectin Receptor MGL Senses N-Acetylgalactosamine on the Unique Staphylococcus aureus ST395 Wall Teichoic Acid. Cell. Microbiol. 2019, e13072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raes, G.; Brys, L.; Dahal, B.K.; Brandt, J.; Grooten, J.; Brombacher, F.; Vanham, G.; Noel, W.; Bogaert, P.; Boonefaes, T.; et al. Macrophage Galactose-Type C-Type Lectins as Novel Markers for Alternatively Activated Macrophages Elicited by Parasitic Infections and Allergic Airway Inflammation. J. Leukoc. Biol. 2005, 77, 321–327. [Google Scholar] [CrossRef]
- Everts, B.; Smits, H.H.; Hokke, C.H.; Yazdanbakhsh, M. Helminths and Dendritic Cells: Sensing and Regulating via Pattern Recognition Receptors, Th2 and Treg Responses. Eur. J. Immunol. 2010, 40, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Fohn, M.; Hakomori, S.-I. Induction of α-N-Acetylgalactosamine-O-Serine/Threonine (Tn) Antigen-mediated Cellular Immune Response for Active Immunotherapy in Mice. Cancer Res. 1991, 51, 1406–1411. [Google Scholar] [PubMed]
- Springer, G.F. Immunoreactive T and Tn Epitopes in Cancer Diagnosis, Prognosis, and Immunotherapy. J. Mol. Med. 1997, 75, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Dusoswa, A.; Verhoeff, J.; Abels, E.; Méndez-Huergo, S.P.; Croci, D.O.; Kuijper, L.H.; de Miguel, E.; Wouters, V.M.C.J.; Best, M.G.; Rodriguez, E.; et al. Glioblastomas Exploit Truncated O-Linked Glycans for Local and Distant Immune Modulation via the Macrophage Galactose-Type Lectin. Proc. Natl. Acad. Sci. USA 2020, 117, 3693–3703. [Google Scholar] [CrossRef]
- Marcelo, F.; Garcia-Martin, F.; Matsushita, T.; Sardinha, J.; Coelho, H.; Oude-Vrielink, A.; Koller, C.; André, S.; Cabrita, E.J.; Gabium, H.-J.; et al. Delineating Binding Modes of Gal/GalNAc and Structural Elements of the Molecular Recognition of Tumor-Associated Mucin Glycopeptides by the Human Macrophage Galactose-Type Lectin. Chem. Eur. J. 2014, 20, 16147–16155. [Google Scholar] [CrossRef]
- Diniz, A.; Coelho, H.; Dias, J.S.; van Vliet, S.J.; Jiménez-Barbero, J.; Corzana, F.; Cabrita, E.J.; Marcelo, F. The Plasticity of Carbohydrate Recognition Domain Dictates the Exquisite Mechanism of Binding of Human Macrophage Galactose-Type Lectin. Chemistry 2019, 25, 13945–13955. [Google Scholar] [CrossRef]
- Marcelo, F.; Supekar, N.; Corzana, F.; van der Horst, J.C.; Vuist, I.M.; Live, D.; Boons, G.-J.P.H.; Smith, D.F.; van Vliet, S.J. Identification of a Secondary Binding Site in Human Macrophage Galactose-Type Lectin by Microarray Studies: Implications for the Molecular Recognition of its Ligands. J. Biol. Chem. 2019, 294, 1300–1311. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Xiao, H.; Wu, R. Specific Identification of Glycoproteins Bearing the Tn Antigen in Human Cells. Angew. Chem. Int. Ed. 2017, 56, 7107–7111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermiston, M.L.; Xu, Z.; Weiss, A. CD45: A Critical Regulator of Signaling Thresholds in Immune Cells. Annu. Rev. Immunol. 2003, 21, 107–137. [Google Scholar] [CrossRef] [PubMed]
- Tong, A.; Nguyen, J.; Lynch, K.W. Differential Expression of CD45 Isoforms is Controlled by the Combined Activity of Basal and Inducible Splicing-Regulatory Elements in Each of the Variable Exons. J. Biol. Chem. 2005, 280, 38297–38304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeill, L.; Cassady, R.L.; Sarkardei, S.; Cooper, J.C.; Morgan, G.; Alexander, D.R. CD45 Isoforms in T Cell Signaling and Development. Immunol. Lett. 2004, 92, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Freire, T.; Lo-Man, R.; Bay, S.; Leclerc, C. Tn Glycosylation of the MUC6 Protein Modulates Its Immunogenicity and Promotes the Induction of Th17-Biased T Cell Responses. J. Biol. Chem. 2011, 286, 7797–7811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo-Man, R.; Vichier-Guerre, S.; Bay, S.; Dériaud, E.; Cantacuzène, D.; Leclerc, C. Anti-Tumor Immunity Provided by a Synthetic Multiple Antiogenic Glycopeptide Displaying a Tri-Tn Glycotope. J. Immunol. 2001, 166, 2849–2854. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, S.J.; Bay, S.; Vuist, I.M.; Kalay, H.; Garcia-Vallejo, J.J.; Leclere, C.; van Kooyk, Y. MGL Signaling Augments TLR2-Mediated Responses for Enhanced IL-10 and TNF-α Secretion. J. Leukoc. Biol. 2013, 94, 315–323. [Google Scholar] [CrossRef]
- Zaal, A.; Li, R.J.E.; Lübbers, J.; Bruijns, S.C.M.; Kalay, H.; van Kooyk, Y.; van Vliet, S.J. Activation of the C-Type Lectin MGL by Terminal GalNAc Ligands Reduces the Glycolytic Activity of Human Dendritic Cells. Front. Immunol. 2020, 11, 305. [Google Scholar] [CrossRef]
- Eggink, L.L.; Roby, K.F.; Cote, R.; Hoober, J.K. An Innovative Immunotherapeutic Strategy for Ovarian Cancer: CLEC10A and Glycomimetic Peptides. J. Immunother. Cancer 2018, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Kolathar, A.R.; Leung, A.K.; Isecke, R.; Brossmer, R.; Drickamer, K.; Weis, W.I. Mechanism of N-Acetylgalactosamine Binding to a C-type Animal Lectin Carbohydrate-recognition Domain. J. Biol. Chem. 1998, 273, 19502–19508. [Google Scholar] [CrossRef] [Green Version]
- London, N.; Movshovitz-Attias, D.; Shueler-Furman, O. The Structural Basis of Peptide-Protein Binding Strategies. Structure 2010, 18, 188–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaszczyk, M.; Kurcinski, M.; Kouza, M.; Wieteska, L.; Debinski, A.; Kolinski, A.; Kmiecik, S. Modeling of protein-peptide interactions using the CABS-dock web server for binding site search and flexible docking. Methods 2016, 93, 72–83. [Google Scholar] [CrossRef]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.H.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.F.; Adema, G.J.; van Kooyk, Y.; Figdor, C.G. Identification of DC-SIGN, a Novel Dendritic Cell-Specific ICAM-3 Receptor that Supports Primary Immune Responses. Cell 2000, 100, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Engering, A.; Geijtenbeek, T.B.H.; van Vliet, S.J.; Wijers, M.; van Liempt, E.; Demaurex, N.; Lanzavecchia, A.; Fransen, J.; Figdor, C.G.; Piguet, V.; et al. The Dendritic Cell-specific Adhesion Receptor DC-SIGN Internalizes Antigen for Presentation to T Cells. J. Immunol. 2002, 168, 2118–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, E.; Touzot, M.; Bohineust, A.; Cappuccio, A.; Chiocchia, G.; Hosmalin, A.; Dalod, M.; Soumelis, V.; Amigorena, S. Human Inflammatory Dendritic Cells Induce Th17 Differentiation. Immunity 2013, 38, 336–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umemoto, T.; Hashimoto, M.; Matsumura, T.; Nakamura-Ishizu, A.; Suda, T. Ca2+-Mitochondria Axis Drives Cell Division in Hematopoietic Stem Cells. J. Exp. Med. 2018, 215, 2097–2113. [Google Scholar] [CrossRef]
- Kurze, A.-K.; Buhs, S.; Eggert, D.; Oliveira-Ferrer, L.; Müller, V.; Niendorf, A.; Wagener, C.; Nollau, P. Immature O-Glycans Recognized by the Macrophage Glycoreceptor CLEC10A (MGL) are Induced by 4-Hydroxy-tamoxifen, oxidative stress and DNA-Damage in Breast Cancer Cells. Cell Commun. Signal. 2019, 17, 107. [Google Scholar] [CrossRef] [Green Version]
- Jondle, C.N.; Sharma, A.; Simonson, T.J.; Larson, B.; Mishra, B.B.; Sharma, J. Macrophage Galactose Lectin-1 (MGL-1) Deficiency Is Associated with Increased Neutrophilia and Hyper Inflammation in Gram-Negative Pneumonia. J. Immunol. 2016, 196, 3088–3096. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, S.J.; van Liempt, E.; Geijtenbeek, T.B.H.; van Kooyk, Y. Differential Regulation of C-Type Lectin Expression on Tolerogenic Dendritic Cell Subsets. Immunobiology 2006, 211, 577–585. [Google Scholar] [CrossRef]
- van Vliet, S.J.; Vuist, I.M.; Lenos, K.; Tefsen, B.; Kalay, H.; Garcia-Vallejo, J.J.; van Kooyk, Y. Human T Cell Activation Results in Extracellular Signal-regulated Kinase (ERK)-Calcineurin-Dependent Exposure of Tn Antigen on the Cell Surface and Binding of the Macrophage Galactose-type Lectin (MGL). J. Biol. Chem. 2013, 288, 27519–27532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sckisel, G.D.; Bouchlaka, M.N.; Monjazeb, A.M.; Crittenden, M.; Curti, B.; Wilkins, D.E.C.; Alderson, K.A.; Sungur, C.M.; Ames, E.; Mirsoian, A.; et al. Out-of-Sequence Signal 3 Paralyzes Primary CD4+ T-Cell-Dependent Immunity. Immunity 2015, 43, 240–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzinger, P. An Innate Sense of Danger. Semin. Immunol. 1998, 10, 399–415. [Google Scholar] [CrossRef] [PubMed]
- Jakubzick, C.V.; Randolph, G.; Henson, P.M. Monocyte Differentiation and Antigen-Presenting Functions. Nat. Rev. Immunol. 2017, 17, 349–362. [Google Scholar] [CrossRef]
- Hervas-Stubbs, S.; Riezu-Boj, J.-I.; Gonzalez, I.; Mancheño, U.; Dubrot, J.; Azpilicueta, A.; Gabari, I.; Palazon, A.; Aranguren, A.; Ruiz, J.; et al. Effects of IFN-α as a Signal-3 Cytokine on Human Naïve and Antigen-Experienced CD8+ T Cells. Eur. J. Immunol. 2010, 40, 3389–3402. [Google Scholar] [CrossRef]
- Keppler, S.J.; Rosenits, K.; Koegl, T.; Vucikuja, S.; Aichele, P. Signal 3 Cytokines as Modulators of Primary Immune Responses during Infections: The Interplay of Type I IFN and IL-12 in CD8 T Cell Responses. PLoS ONE 2012, 7, e40865. [Google Scholar] [CrossRef] [Green Version]
- Diamond, M.S.; Kinder, M.; Matsushita, H.; Mashayekhi, M.; Dunn, G.P.; Archambault, J.M.; Lee, H.; Arthur, C.D.; White, J.M.; Kalinke, U.; et al. Type I Interferon is Selectively Required by Dendritic Cells for Immune Rejection of Tumors. J. Exp. Med. 2011, 208, 1989–2001. [Google Scholar] [CrossRef]
- Raker, V.K.; Domogalla, M.P.; Steinbrink, K. Tolerogenic Dendritic Cells for Regulatory T Cell Induction in Man. Front. Immunol. 2015, 6, 569. [Google Scholar] [CrossRef] [Green Version]
- Domogalla, M.P.; Rostan, P.V.; Raker, V.K.; Steinbrink, K. Tolerance through Education: How Tolerogenic Dendritic Cells Shape Immunity. Front. Immunol. 2017, 8, 1764. [Google Scholar] [CrossRef] [Green Version]
- Heissmeyer, V.; Macián, F.; Im, S.-H.; Varma, R.; Feske, S.; Venuprasad, K.; Gu, H.; Liu, Y.-C.; Dustin, M.L.; Rao, A. Calcineurin Imposes T Cell Unresponsiveness Through Targeted Proteolysis of Signaling Proteins. Nat. Immunol. 2004, 5, 255–265. [Google Scholar] [CrossRef]
- Razzadq, T.M.; Ozegbe, P.; Jury, E.C.; Sembi, P.; Blackwell, N.M.; Kabouridis, P.S. Regulation of T-Cell Receptor Signaling by Membrane Microdomains. Immunology 2004, 113, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.R.; Rao, A. NFAT, Immunity and Cancer: A Transcription Factor Comes of Age. Nat. Rev. Immunol. 2010, 10, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, M.; Jagannathan, S.; Vallabh, S.; Kartashov, A.V.; Chen, X.; Weirauch, M.T.; Barski, A. AP-1 Activity Induced by Co-stimulation is Required for Chromatin Opening During T Cell Activation. J. Exp. Med. 2020, 217, e20182009. [Google Scholar] [CrossRef]
- Powell, J.D. The Induction and Maintenance of T Cell Anergy. Clin. Immunol. 2006, 120, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Hogan, P.G. Calcium-NFAT Transcriptional Signaling in T Cell Activation and T Cell Exhaustion. Cell Calcium 2017, 63, 66–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, R.M.; Turley, S.; Mellman, I.; Inaba, K. The Induction of Tolerance by Dendritic Cells That Have Captured Apoptotic Cells. J. Exp. Med. 2000, 191, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Wherry, E.J.; Kurachi, M. Molecular and Cellular Insights into T Cell Exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Saeidi, A.; Zandi, K.; Cheok, Y.Y.; Saeidi, H.; Wong, W.F.; Lee, C.Y.Q.; Cheong, H.C.; Yong, Y.K.; Larsson, M.; Shankar, E.M. T-Cell Exhaustion in Chronic Infections: Reversing the State of Exhaustion and Reinvigorating Optimal Protective Immune Responses. Front. Immunol. 2018, 9, 2569. [Google Scholar] [CrossRef]
- Nossal, G.J.V. Cellular Mechanisms of Immunologic Tolerance. Annu. Rev. Immunol. 1983, 1, 33–62. [Google Scholar] [CrossRef]
- Heath, W.R.; Carbone, F.R. Cross-Presentation, Dendritic Cells, Tolerance, and Immunity. Annu. Rev. Immunol. 2001, 19, 47–64. [Google Scholar] [CrossRef]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and Therapeutic Potential of Interleukin-10. J. Exp. Med. 2019, 217, e20190418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium Signaling: Dynamics, Homeostatis and Remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carafoli, E.; Krebs, J. Why Calcium? How Calcium Became the Best Communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef] [Green Version]
- Marois, L.; Paré, G.; Vaillancourt, M.; Rollet-Labelle, E.; Naccache, P.H. FcγRIIIb Triggers Raft-dependent Calcium Influx in IgG-mediated Responses in Human Neutrophils. J. Biol. Chem. 2011, 286, 3509–3519. [Google Scholar] [CrossRef] [Green Version]
- Nunes, P.; Demaurex, N. The Role of Calcium Signaling in Phagocytosis. J. Leukoc. Biol. 2010, 88, 57–68. [Google Scholar] [CrossRef]
- Mellado-Sánchez, G.; Vivanco-Cid, H.; Sumoza-Toledo, A. Regulation of Murine Dendritic Cell Functions by Calcium Channels. SOJ Immunol. 2014, 2, 1–6. [Google Scholar]
- Feske, S. Calcium signaling in Lymphocyte Activation and Disease. Nat. Rev. Immunol. 2007, 7, 690–702. [Google Scholar] [CrossRef]
- Izquierdo, J.H.; Bonella-Abadia, F.; Cañas, C.A.; Tobón, G.J. Calcium Channels, Intracellular Signaling and Autoimmunity. Reumatol. Clin. 2014, 10, 43–47. [Google Scholar] [CrossRef]
- Li, D.; Romain, G.; Flamar, A.-L.; Duluc, D.; Dullaers, M.; Li, X.-H.; Zurawski, S.; Bosquet, N.; Palucka, A.K.; Le Grand, R.; et al. Targeting Self- and Foreign Antigens to Dendritic Cells via DC-ASPGR Generates IL-10-Producing Suppressive CD4+ T Cells. J. Exp. Med. 2012, 209, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.-Q.; Kao, K.J. Suppression of Interleukin-12 Production through Endogenously Secreted Interleukin-10 in Activated Dendritic Cells: Involvement of Activation of Extracellular Signal-Regulated Protein Kinase. Scand. J. Immunol. 2003, 58, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Yan, W.; Zheng, H.; Du, Q.; Zhang, L.; Ban, Y.; Li, N.; Fang Wei, F. Regulation of IL-10 and IL-12 Production and Function in Macrophages and Dendritic Cells [version 1; peer review: 3 approved]. F1000Research 2015, 4, 1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, M.H.; Berg, H. Interleukin-10 Expression is Induced by Increase of Intracellular Calcium Levels in the Monocytic Cell Line U937. Pflügers Archiv 1998, 435, 868–870. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.K.; Wang, L.; Ivashkiv, L.B. Calcium-Activated Pathways and Oxidative Burst Mediate Zymosan-Induced Signaling and IL-10 Production in Human Macrophages. J. Immunol. 2010, 184, 5545–5552. [Google Scholar] [CrossRef]
- Rafiq, K.; Charitidou, L.; Bullens, D.M.A.; Kasran, A.; Lorré, K.; Ceuppens, J.L.; van Gool, S.W. Regulation of the IL-10 Production by Human T Cells. Scand. J. Immunol. 2001, 53, 139–147. [Google Scholar] [CrossRef]
- Boubali, S.; Liopeta, K.; Virgillo, L.; Thyphronitis, G.; Mavrothallassitis, G.; Dimitracopoulos, G.; Paliogianni, F. Calcium/Calmodulin-Dependent Protein Kinase II Regulates IL-10 Production by Human T Lymphocytes: A Distinct Target in the Calcium Dependent Pathway. Mol. Immunol. 2012, 52, 51–60. [Google Scholar] [CrossRef]
- Faries, M.B.; Bedrosian, I.; Xu, S.; Koski, G.; Roros, J.G.; Moise, M.A.; Nguyen, H.Q.; Engels, F.H.C.; Cohen, P.A.; Czemiecki, B.J. Calcium Signaling Inhibits Interleukin-12 Production and Activates CD83+ Dendritic Cells that Induce Th2 Cell Development. Blood 2001, 98, 2489–2497. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, N.; Zhu, Y.; Yang, Y.; Chen, X.; Fan, S.; Chen, Q.; Zhou, H.; Zheng, J. Inhibition of Extracellular Calcium Influx Results in Enhanced IL-12 Production in LPS-Treated Murine Macrophages by Downregulation of the CaMKKβ-AMPK-SIRT1 Signaling Pathway. Med. Inflamm. 2016, 2016, 6152713. [Google Scholar] [CrossRef] [Green Version]
- Bauer, B.; Steinle, A. HemITAM: A Single Tyrosine Motif that Packs a Punch. Sci. Signal. 2017, 10, eaan3676. [Google Scholar] [CrossRef] [Green Version]
- Schülke, S. Induction of interleukin-10 producing dendritic cells as a tool to suppress allergen-specific T helper 2 responses. Front. Immunol. 2018, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Sheng, M.; Thompson, M.A.; Greenberg, M.E. CREB: A Ca2+-regulated transcription factor phosphorylated by calmodulin-dependent kinases. Science 1991, 252, 1427–1430. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, L.; Thompson, R.; Chan, C. CREB Modulates Calcium Signaling in cAMP-Induced Bone Marrow Stromal Cells (BMSCs). Cell Calcium 2014, 56, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Kushchayev, S.V.; Sankar, T.; Eggink, L.L.; Kushchayeva, Y.S.; Wiener, P.C.; Hoober, J.K.; Eschbacher, J.; Liu, R.; Shi, F.D.; Abdelwahab, M.G.; et al. Monocyte Galactose/N-Acetylgalactosamine-Specific C-Type Lectin Receptor Stimulant Immunotherapy of an Experimental Glioma. Part II: Combination with External Radiation Improves Survival. Cancer Manag. Res. 2012, 4, 325–334. [Google Scholar] [PubMed] [Green Version]
- Mayes, P.A.; Hance, K.W.; Hoos, A. The Promise and Challenges of Immune Agonist Antibody Development in Cancer. Nat. Rev. Drug Discov. 2018, 17, 509–527. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, J.M.; Barrera, F.N. Membrane Receptor Activation Mechanisms and Transmembrane Peptide Tools to Elucidate Them. J. Biol. Chem. 2020, 295, 1792–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martyanov, A.A.; Balabin, F.A.; Dunster, J.L.; Panteleev, M.A.; Gibbins, J.M.; Sveshnikova, A.N. Control of Platelet CLEC-2-Mediated Activation by Receptor Clustering and Tyrosine Kinase Signaling. Biophys. J. 2020, 118, 1–15. [Google Scholar] [CrossRef]
- Jégouzo, S.A.F.; Quintero-Martinez, A.; Ouyang, X.; dos Santos, Á.; Taylor, M.E.; Drickamer, K. Organization of the Extracellular Portion of the Macrophage Galactose Receptor: A Trimeric Cluster of Simple Binding Sites for N-Acetylgalactosamine. Glycobiology 2013, 23, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Finch, E.A.; Turner, T.J.; Goldin, S.M. Calcium as a Coagonist of Inositol 1,4,5-Triphosphate-Induced Calcium Release. Science 1991, 252, 443–446. [Google Scholar] [CrossRef]
- Bezprozvanny, I.; Watras, J.; Ehrlich, B.E. Bell-Shaped Calcium-Response Curves of Ins(1,4,5)P3 and Calcium-Gated Channels from Endoplasmic Reticulum of Cerebellum. Nature 1991, 351, 751–754. [Google Scholar] [CrossRef]



© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoober, J.K. ASGR1 and Its Enigmatic Relative, CLEC10A. Int. J. Mol. Sci. 2020, 21, 4818. https://doi.org/10.3390/ijms21144818
Hoober JK. ASGR1 and Its Enigmatic Relative, CLEC10A. International Journal of Molecular Sciences. 2020; 21(14):4818. https://doi.org/10.3390/ijms21144818
Chicago/Turabian StyleHoober, J. Kenneth. 2020. "ASGR1 and Its Enigmatic Relative, CLEC10A" International Journal of Molecular Sciences 21, no. 14: 4818. https://doi.org/10.3390/ijms21144818
APA StyleHoober, J. K. (2020). ASGR1 and Its Enigmatic Relative, CLEC10A. International Journal of Molecular Sciences, 21(14), 4818. https://doi.org/10.3390/ijms21144818