EPA and DHA Fatty Acids Induce a Remodeling of Tumor Vasculature and Potentiate Docetaxel Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
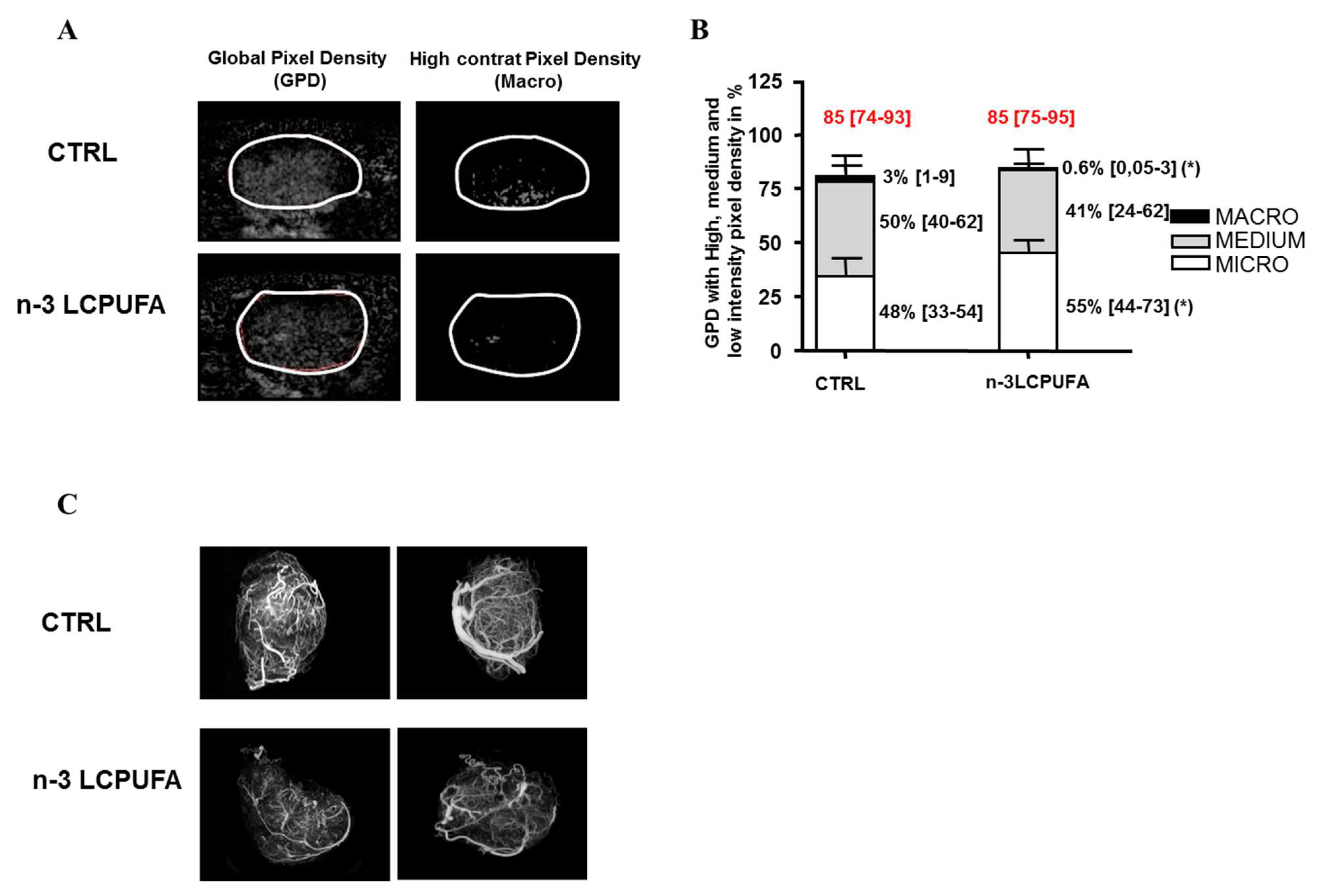
2.1. n-3 LCPUFA Supplementation Induces a Remodeling of the Mammary Tumor Vasculature Prior to the Initiation of Chemotherapy
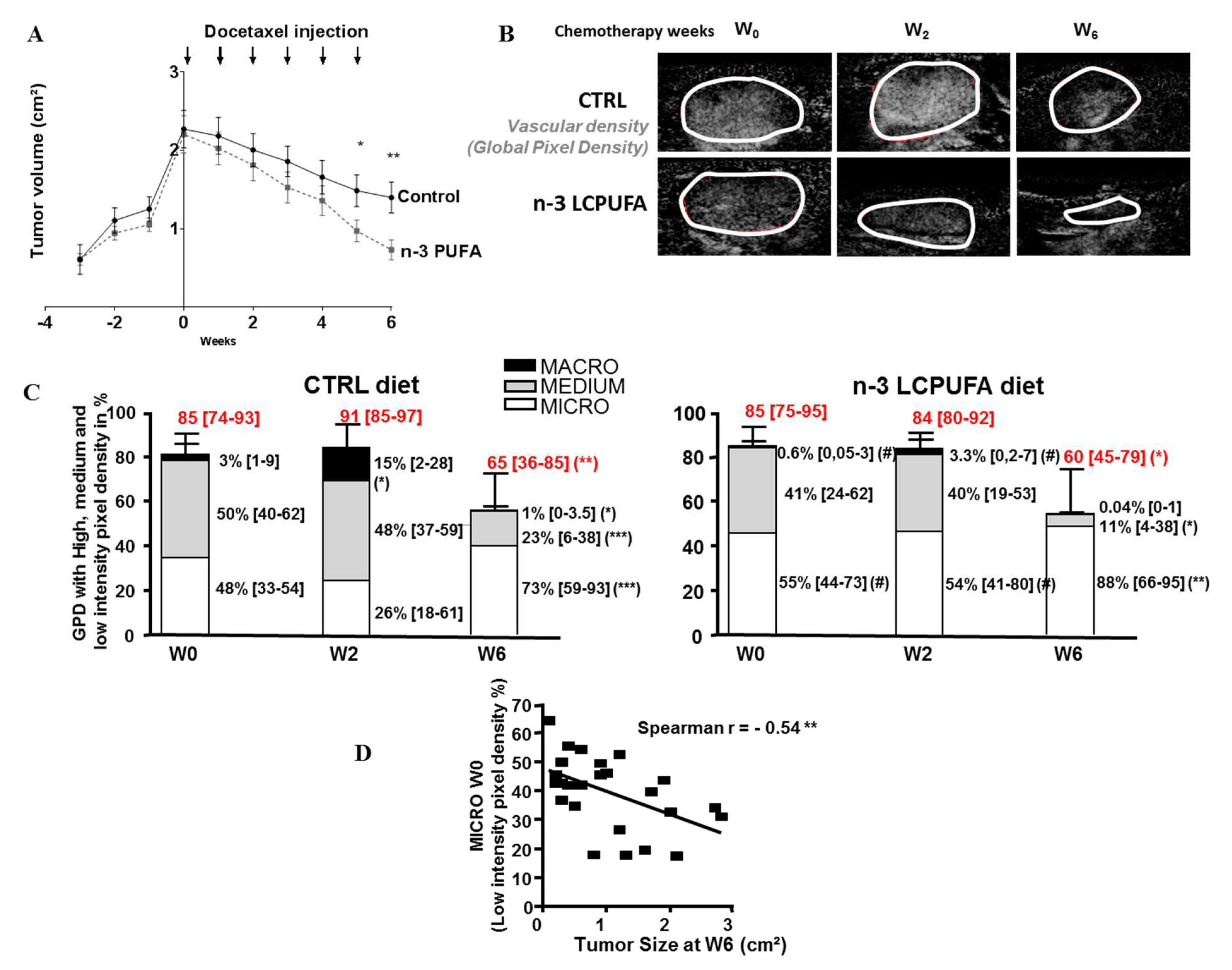
2.2. The Microvasculature Proportion Remains Preponderant in n-3 LCPUFA Tumors During Docetaxel Treatment and Correlates with Tumor Regression
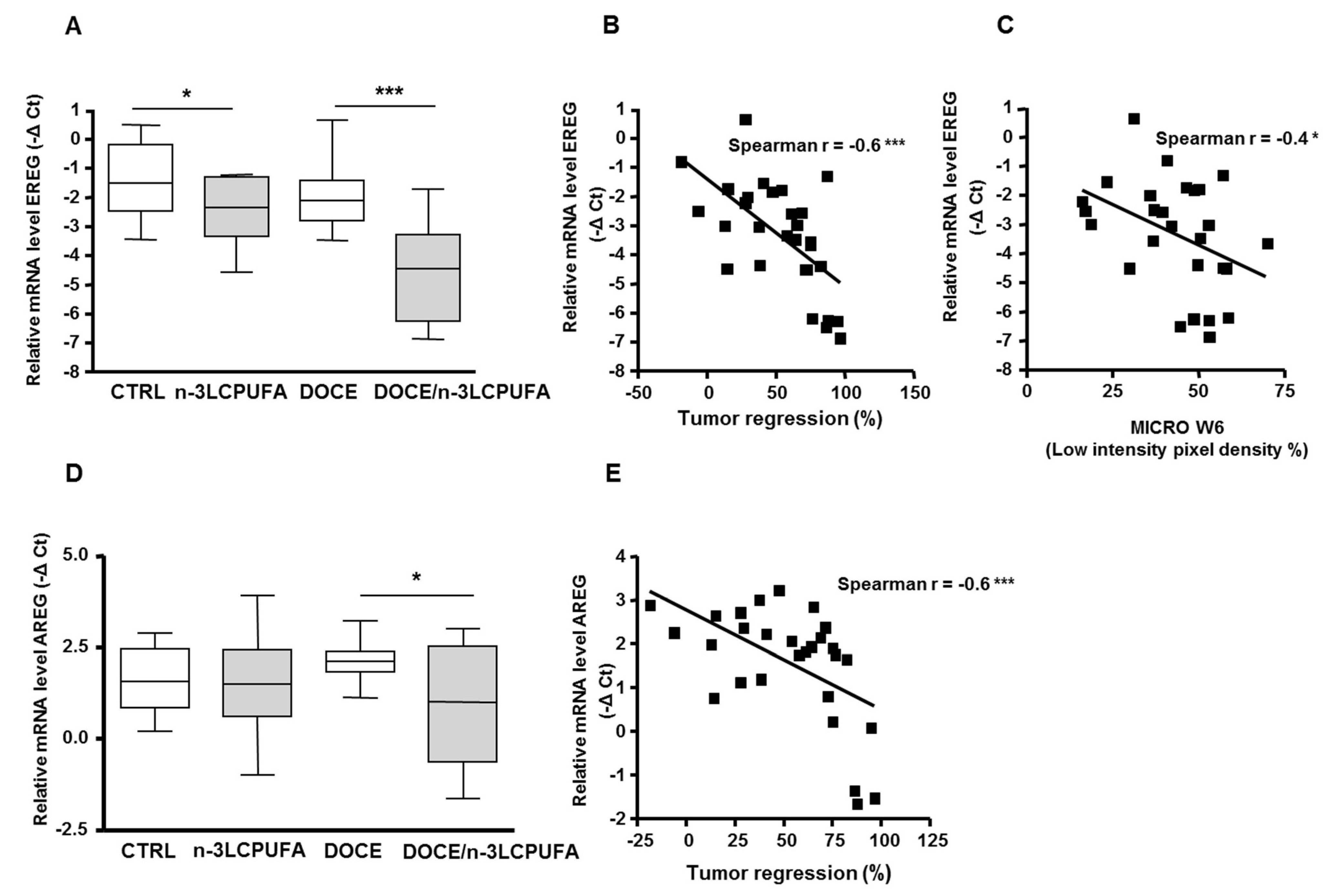
2.3. Epiregulin and Amphiregulin are New Molecular Targets of the n-3 LCPUFA-Enriched Diet, and Their mRNA Levels Correlate With Docetaxel Efficacy
3. Discussion
4. Materials and Methods
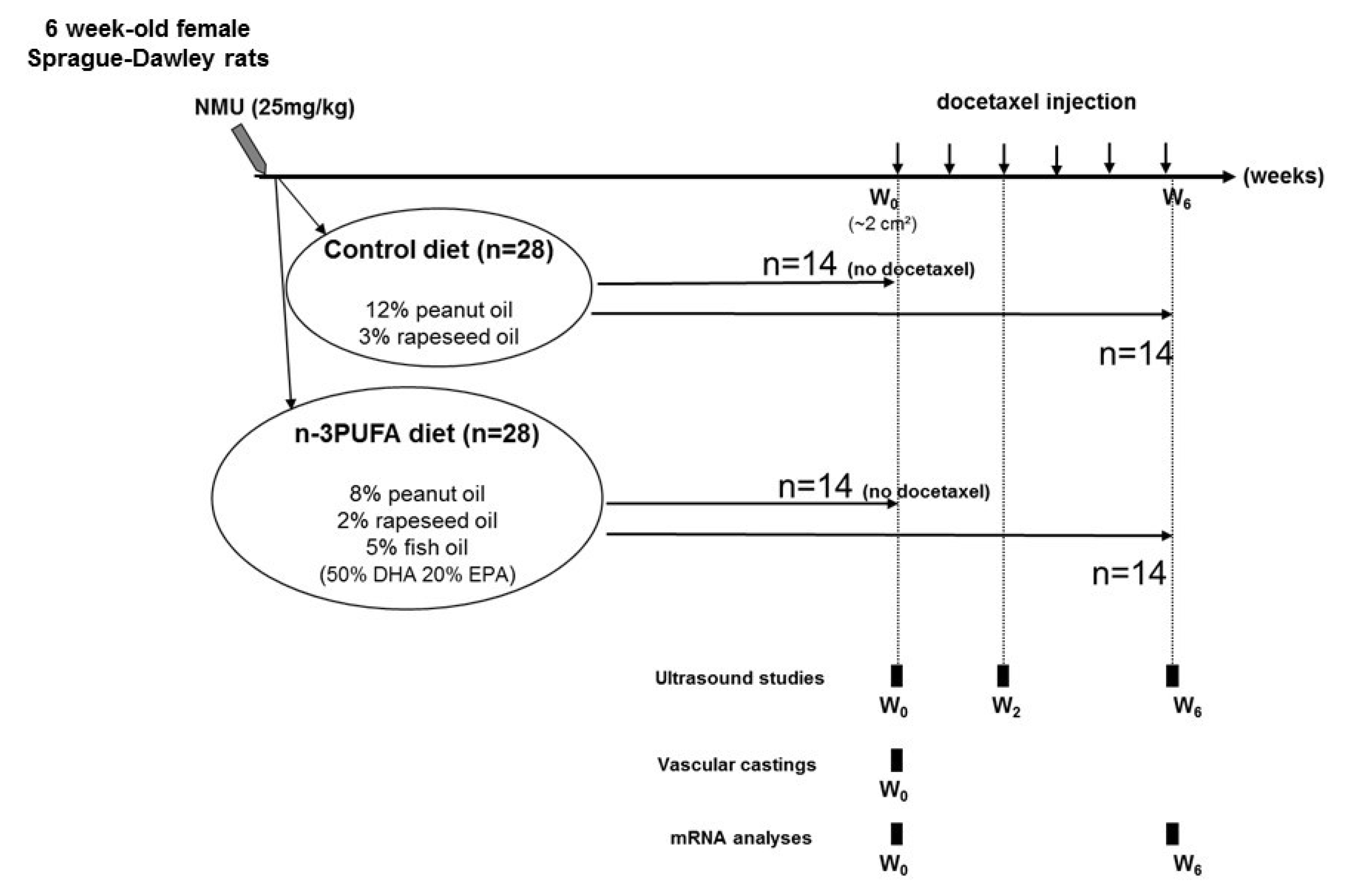
4.1. In Vivo Study
4.2. Functional Vascularization and Quantification of Micro, Medium and Macrovascularization
4.3. Casting of the Tumor Vasculature
4.4. Oligonucleotide PCR Arrays and Real-Time Quantitative PCR Analysis
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AREG | Amphiregulin |
| CEUS | Contrast-Enhanced Ultrasound |
| DHA | Docosahexaenoic Acid |
| EPA | Eicosapentaenoic Acid |
| EREG | Epiregulin |
| GPD | Global pixel density |
| n-3 LCPUFA | n-3 Long Chain Polyunsaturated Fatty Acids |
| NMU | N-methylnitrosourea |
| VEGF | Vascular endothelial growth factor |
References
- Fukumura, D.; Jain, R.K. Tumor microvasculature and microenvironment: Targets for anti-angiogenesis and normalization. Microvasc. Res. 2007, 74, 72–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, R.T.; Boucher, Y.; Kozin, S.V.; Winkler, F.; Hicklin, D.J.; Jain, R.K. Vascular Normalization by Vascular Endothelial Growth Factor Receptor 2 Blockade Induces a Pressure Gradient Across the Vasculature and Improves Drug Penetration in Tumors. Cancer Res. 2004, 64, 3731–3736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, R.K. Normalization of Tumor Vasculature: An Emerging Concept in Antiangiogenic Therapy. Science 2005, 307, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Maheo, K.; Chevalier, S.; Vibet, S.; Bougnoux, P.; Richard, S.; Sérrière, S.; Bleuzen, A.; Tranquart, F.; Goupille, C. Non-invasive quantification of tumor vascular architecture during docetaxel-chemotherapy. Breast Cancer Res. Treat. 2012, 134, 1013–1025. [Google Scholar] [CrossRef]
- Biondo, P.D.; Brindley, D.N.; Sawyer, M.B.; Field, C.J. The potential for treatment with dietary long-chain polyunsaturated n-3 fatty acids during chemotherapy. J. Nutr. Biochem. 2008, 19, 787–796. [Google Scholar] [CrossRef]
- Calviello, G.; Serini, S.; Piccioni, E.; Pessina, G. Antineoplastic Effects of N-3 Polyunsaturated Fatty Acids in Combination With Drugs and Radiotherapy: Preventive and Therapeutic Strategies. Nutr. Cancer 2009, 61, 287–301. [Google Scholar] [CrossRef]
- Bougnoux, P.; Hajjaji, N.; Maheo, K.; Couet, C.; Chevalier, S. Fatty acids and breast cancer: Sensitization to treatments and prevention of metastatic re-growth. Prog. Lipid Res. 2011, 49, 76–86. [Google Scholar] [CrossRef]
- Colas, S.; Mahéo, K.; Denis, F.; Goupille, C.; Hoinard, C.; Champeroux, P.; Tranquart, F.; Bougnoux, P. Sensitization by Dietary Docosahexaenoic Acid of Rat Mammary Carcinoma to Anthracycline: A Role for Tumor Vascularization. Clin. Cancer Res. 2006, 12, 5879–5886. [Google Scholar] [CrossRef] [Green Version]
- Rose, D.P.; Connolly, J.M. Antiangiogenicity of docosahexaenoic acid and its role in the suppression of breast cancer cell growth in nude mice. Int. J. Oncol. 1999, 15, 1011–1015. [Google Scholar] [CrossRef]
- Szymczak, M.; Murray, M.; Petrović, N. Modulation of angiogenesis by ω-3 polyunsaturated fatty acids is mediated by cyclooxygenases. Blood 2008, 111, 3514–3521. [Google Scholar] [CrossRef]
- Zhang, G.; Panigrahy, D.; Mahakian, L.M.; Yang, J.; Liu, J.-Y.; Lee, K.S.S.; Wettersten, H.I.; Ulu, A.; Hu, X.; Tam, S.; et al. Epoxy metabolites of docosahexaenoic acid (DHA) inhibit angiogenesis, tumor growth, and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 6530–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornfeld, S.; Goupille, C.; Vibet, S.; Chevalier, S.; Pinet, A.; Lebeau, J.; Tranquart, F.; Bougnoux, P.; Martel, E.; Maurin, A.; et al. Reducing endothelial NOS activation and interstitial fluid pressure with n-3 PUFA offset tumor chemoresistance. Carcinogenesis 2012, 33, 260–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, R.K. Molecular regulation of vessel maturation. Nat. Med. 2003, 9, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Baluk, P.; Hashizume, H.; McDonald, D.M. Cellular abnormalities of blood vessels as targets in cancer. Curr. Opin. Genet. Dev. 2005, 15, 102–111. [Google Scholar] [CrossRef]
- Heldin, C.-H.; Rubin, K.; Pietras, K.; Östman, A. High interstitial fluid pressure—An obstacle in cancer therapy. Nat. Rev. Cancer 2004, 4, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.; Wong, H.-K.; Jain, R.K. Vascular Normalization as a Therapeutic Strategy for Malignant and Nonmalignant Disease. Cold Spring Harb. Perspect. Med. 2011, 2, a006486. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Stylianopoulos, T.; Duda, D.G.; Fukumura, D.; Jain, R.K. Benefits of Vascular Normalization Are Dose and Time Dependent. Cancer Res. 2013, 73, 7144–7146. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Wieczorek, C.; Schöttle, J.; Siobal, M.; Hinze, Y.; Franz, T.; Florin, A.; Adamczak, J.; Heukamp, L.; Neumaier, B.; et al. Transient Antiangiogenic Treatment Improves Delivery of Cytotoxic Compounds and Therapeutic Outcome in Lung Cancer. Cancer Res. 2014, 74, 2816–2824. [Google Scholar] [CrossRef] [Green Version]
- Weiss, A.; Bonvin, D.; Berndsen, R.H.; Scherrer, E.; Wong, T.J.; Dyson, P.J.; Griffioen, A.W.; Nowak-Sliwinska, P. Angiostatic treatment prior to chemo- or photodynamic therapy improves anti-tumor efficacy. Sci. Rep. 2015, 5, 8990. [Google Scholar] [CrossRef] [Green Version]
- Claes, A.; Wesseling, P.; Jeuken, J.; Maass, C.; Heerschap, A.; Leenders, W.P.J. Antiangiogenic compounds interfere with chemotherapy of brain tumors due to vessel normalization. Mol. Cancer Ther. 2008, 7, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; Jain, R.K. Principles and mechanisms of vessel normalization for cancer and other angiogenic diseases. Nat. Rev. Drug Discov. 2011, 10, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Winkler, F.; Kozin, S.V.; Tong, R.T.; Chae, S.-S.; Booth, M.F.; Garkavtsev, I.; Xu, L.; Hicklin, D.J.; Fukumura, D.; Di Tomaso, E.; et al. Kinetics of vascular normalization by VEGFR2 blockade governs brain tumor response to radiation. Cancer Cell 2004, 6, 553–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, S.; Matsunaga, S.; Imanishi, M.; Maekawa, Y.; Kitano, H.; Takeuchi, H.; Tomita, S. Tumour blood vessel normalisation by prolyl hydroxylase inhibitor repaired sensitivity to chemotherapy in a tumour mouse model. Sci. Rep. 2017, 7, 45621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuzuki, T.; Shibata, A.; Kawakami, Y.; Nakagaya, K.; Miyazawa, T. Anti-Angiogenic Effects of Conjugated Docosahexaenoic Acidin Vitroandin Vivo. Biosci. Biotechnol. Biochem. 2007, 71, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, M.; Murota, S.-I.; Morita, I. Docosapentaenoic acid (22:5, n-3) suppressed tube-forming activity in endothelial cells induced by vascular endothelial growth factor. Prostaglandins, Leukot. Essent. Fat. Acids 2003, 68, 337–342. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids and inflammation. Prostaglandins, Leukot. Essent. Fat. Acids 2006, 75, 197–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C. n-3 Fatty acids, inflammation and immunity: New mechanisms to explain old actions. Proc. Nutr. Soc. 2013, 72, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Connor, K.M.; SanGiovanni, J.P.; Löfqvist, C.; Aderman, C.M.; Chen, J.; Higuchi, A.; Hong, S.; Pravda, E.A.; Majchrzak, S.; Carper, D.; et al. Increased dietary intake of ω-3-polyunsaturated fatty acids reduces pathological retinal angiogenesis. Nat. Med. 2007, 13, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Calviello, G.; Di Nicuolo, F.; Gragnoli, S.; Piccioni, E.; Serini, S.; Maggiano, N.; Tringali, G.; Navarra, P.; Ranelletti, F.O.; Palozza, P. n-3 PUFAs reduce VEGF expression in human colon cancer cells modulating the COX-2/PGE2 induced ERK-1 and -2 and HIF-1alpha induction pathway. Carcinogenesis 2004, 25, 2303–2310. [Google Scholar] [CrossRef] [Green Version]
- Hannafon, B.N.; Carpenter, K.J.; Berry, W.R.; Janknecht, R.; Dooley, W.; Ding, W.-Q. Exosome-mediated microRNA signaling from breast cancer cells is altered by the anti-angiogenesis agent docosahexaenoic acid (DHA). Mol. Cancer 2016, 14, 133. [Google Scholar] [CrossRef] [Green Version]
- Riese, D.; Cullum, R.L. Epiregulin: Roles in normal physiology and cancer. Semin. Cell Dev. Boil. 2014, 28, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Busser, B.; Sancey, L.; Brambilla, E.; Coll, J.-L.; Hurbin, A. The multiple roles of amphiregulin in human cancer. Biochim. Biophys. Acta (BBA)—Bioenerg. 2011, 1816, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, M.; Bohrer, L.R.; Brady, N.J.; Chuntova, P.; Kemp, S.E.; Wardwell, C.T.; Nelson, A.C.; Schwertfeger, K.L. Epiregulin contributes to breast tumorigenesis through regulating matrix metalloproteinase 1 and promoting cell survival. Mol. Cancer 2015, 14, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckstein, N.; Servan, K.; Girard, L.; Cai, D.; Von Jonquieres, G.; Jaehde, U.; Kassack, M.U.; Gazdar, A.F.; Minna, J.D.; Royer, H.-D. Epidermal growth factor receptor pathway analysis identifies amphiregulin as a key factor for cisplatin resistance of human breast cancer cells. J. Boil. Chem. 2008, 283, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.; De Roock, W.; Piessevaux, H.; Van Oirbeek, R.; Biesmans, B.; De Schutter, J.; Fieuws, S.; Vandesompele, J.; Peeters, M.; Van Laethem, J.-L.; et al. Amphiregulin and Epiregulin mRNA Expression in Primary Tumors Predicts Outcome in Metastatic Colorectal Cancer Treated With Cetuximab. J. Clin. Oncol. 2009, 27, 5068–5074. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.A.; Dos Santos, M.L.; Gimenes, K.P.; Silva, W.A. Transcriptome changes induced by docetaxel in human mammary cell lines expressing different levels of ERBB2. Int. J. Mol. Med. 2009, 23, 733–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Vargas, H.; Palacios, J.; Moreno-Bueno, G. Molecular profiling of docetaxel cytotoxicity in breast cancer cells: Uncoupling of aberrant mitosis and apoptosis. Oncogene 2007, 26, 2902–2913. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-W.; Kim, D.K.; Min, A.; Lee, K.-H.; Nam, H.-J.; Kim, J.H.; Kim, J.-S.; Kim, T.Y.; Im, S.-A.; Park, I.A. Amphiregulin confers trastuzumab resistance via AKT and ERK activation in HER2-positive breast cancer. J. Cancer Res. Clin. Oncol. 2016, 142, 157–165. [Google Scholar] [CrossRef]
- Tinhofer, I.; Klinghammer, K.; Weichert, W.; Stenzinger, A.; Gauler, T.; Budach, V.; Keilholz, U.; Knödler, M. Expression of Amphiregulin and EGFRvIII Affect Outcome of Patients with Squamous Cell Carcinoma of the Head and Neck Receiving Cetuximab-Docetaxel Treatment. Clin. Cancer Res. 2011, 17, 5197–5204. [Google Scholar] [CrossRef] [Green Version]
- Yamagata, K. Docosahexaenoic acid regulates vascular endothelial cell function and prevents cardiovascular disease. Lipids Heal. Dis. 2017, 16, 118. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.A.; Mourtzakis, M.; Chu, Q.S.C.; Baracos, V.E.; Reiman, T.; Mazurak, V.C. Supplementation with fish oil increases first-line chemotherapy efficacy in patients with advanced nonsmall cell lung cancer. Cancer 2011, 117, 3774–3780. [Google Scholar] [CrossRef] [PubMed]
- Cockbain, A.J.; Toogood, G.; Hull, M.A. Omega-3 polyunsaturated fatty acids for the treatment and prevention of colorectal cancer. Gut 2011, 61, 135–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broillet, A.; Hantson, J.; Ruegg, C.; Messager, T.; Schneider, M. Assessment of microvascular perfusion changes in a rat breast tumor model using SonoVue to monitor the effects of different anti-angiogenic therapies. Acad. Radiol. 2005, 12 (Suppl. S1), S28–S33. [Google Scholar] [CrossRef]
- Schneider, M. Characteristics of SonoVuetrade mark. Echocardiography 1999, 16, 743–746. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goupille, C.; Vibet, S.; Frank, P.G.; Mahéo, K. EPA and DHA Fatty Acids Induce a Remodeling of Tumor Vasculature and Potentiate Docetaxel Activity. Int. J. Mol. Sci. 2020, 21, 4965. https://doi.org/10.3390/ijms21144965
Goupille C, Vibet S, Frank PG, Mahéo K. EPA and DHA Fatty Acids Induce a Remodeling of Tumor Vasculature and Potentiate Docetaxel Activity. International Journal of Molecular Sciences. 2020; 21(14):4965. https://doi.org/10.3390/ijms21144965
Chicago/Turabian StyleGoupille, Caroline, Sophie Vibet, Philippe G. Frank, and Karine Mahéo. 2020. "EPA and DHA Fatty Acids Induce a Remodeling of Tumor Vasculature and Potentiate Docetaxel Activity" International Journal of Molecular Sciences 21, no. 14: 4965. https://doi.org/10.3390/ijms21144965
APA StyleGoupille, C., Vibet, S., Frank, P. G., & Mahéo, K. (2020). EPA and DHA Fatty Acids Induce a Remodeling of Tumor Vasculature and Potentiate Docetaxel Activity. International Journal of Molecular Sciences, 21(14), 4965. https://doi.org/10.3390/ijms21144965