Caveolae-Associated Protein 3 (Cavin-3) Influences Adipogenesis via TACE-Mediated Pref-1 Shedding



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
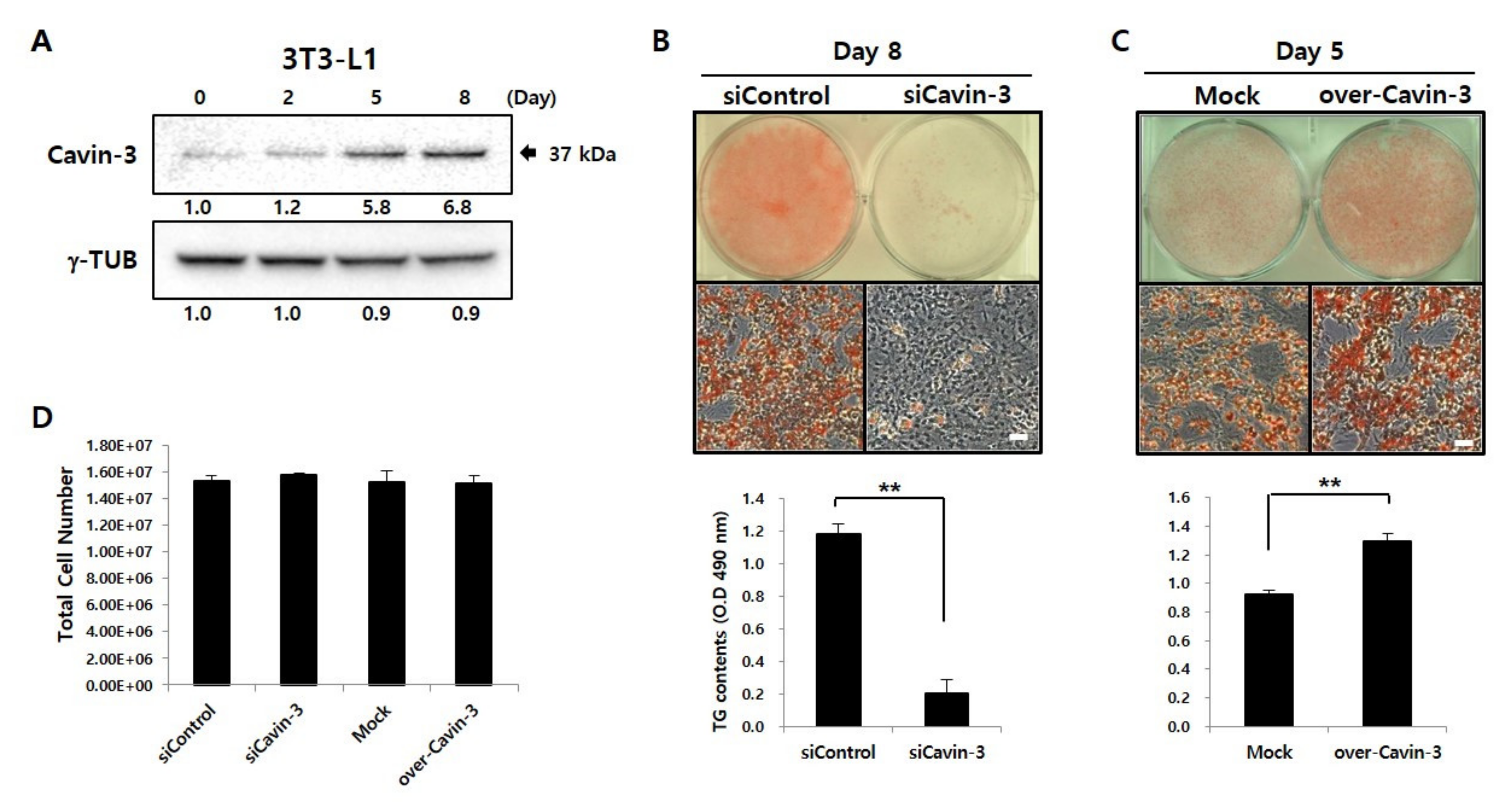
2.1. Physiological Effect of Cavin-3 in Knockdown or Overexpression Cell Lines
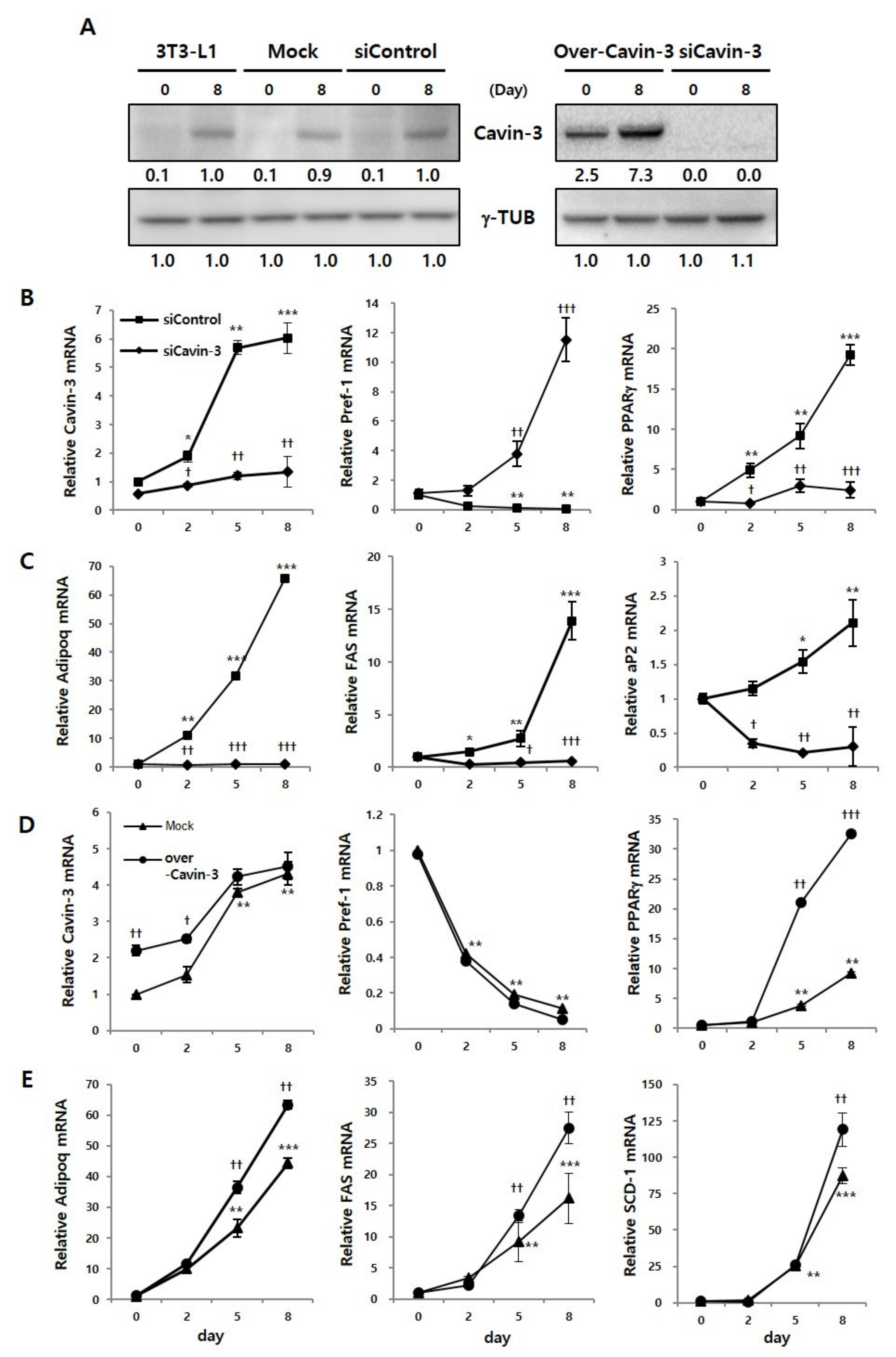
2.2. Effect of Cavin-3 Knockdown or Overexpression on Adipogenesis-Related mRNA Expression
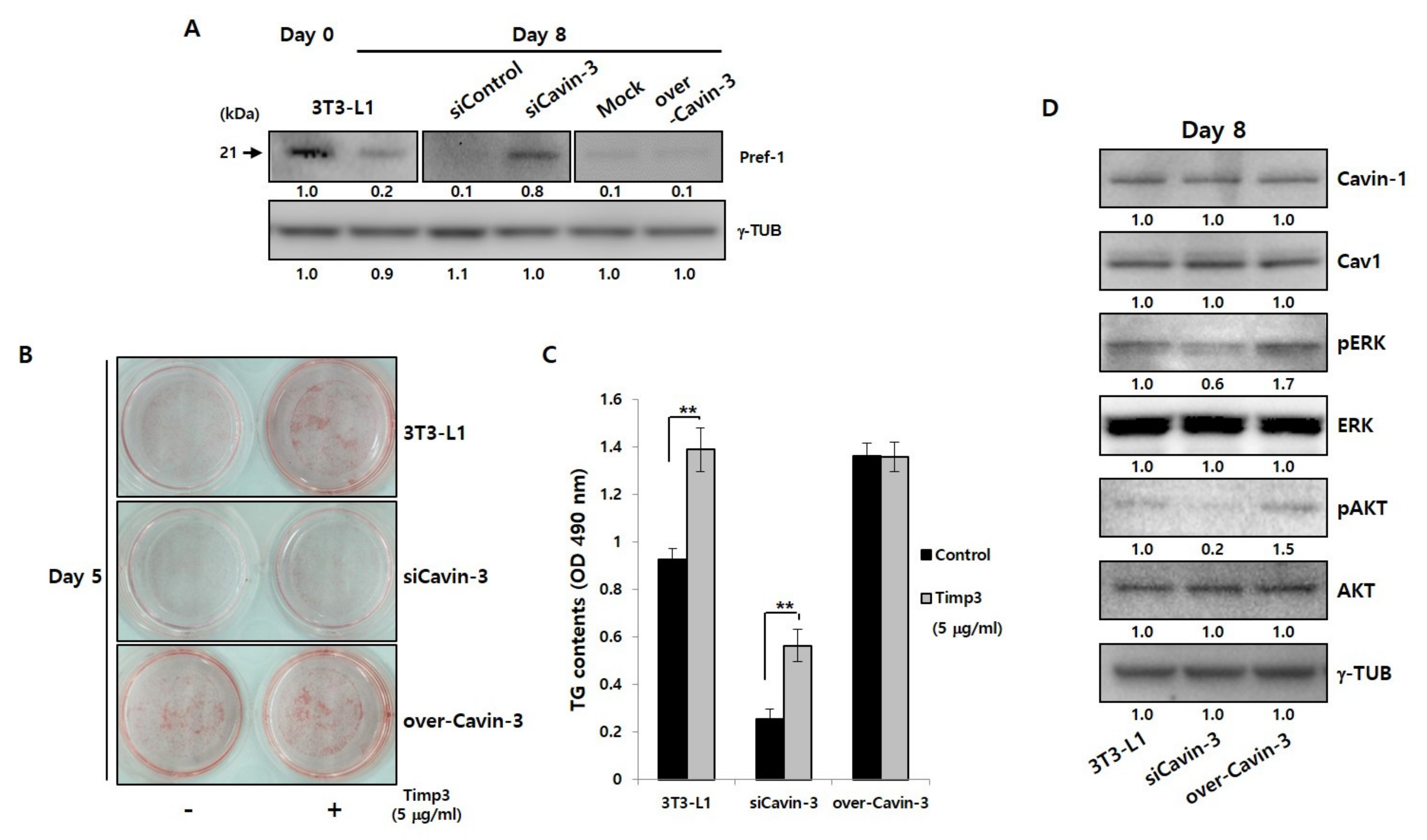
2.3. Changes in Cavin-3 Gene Expression Influence the Cleavage of Pref-1
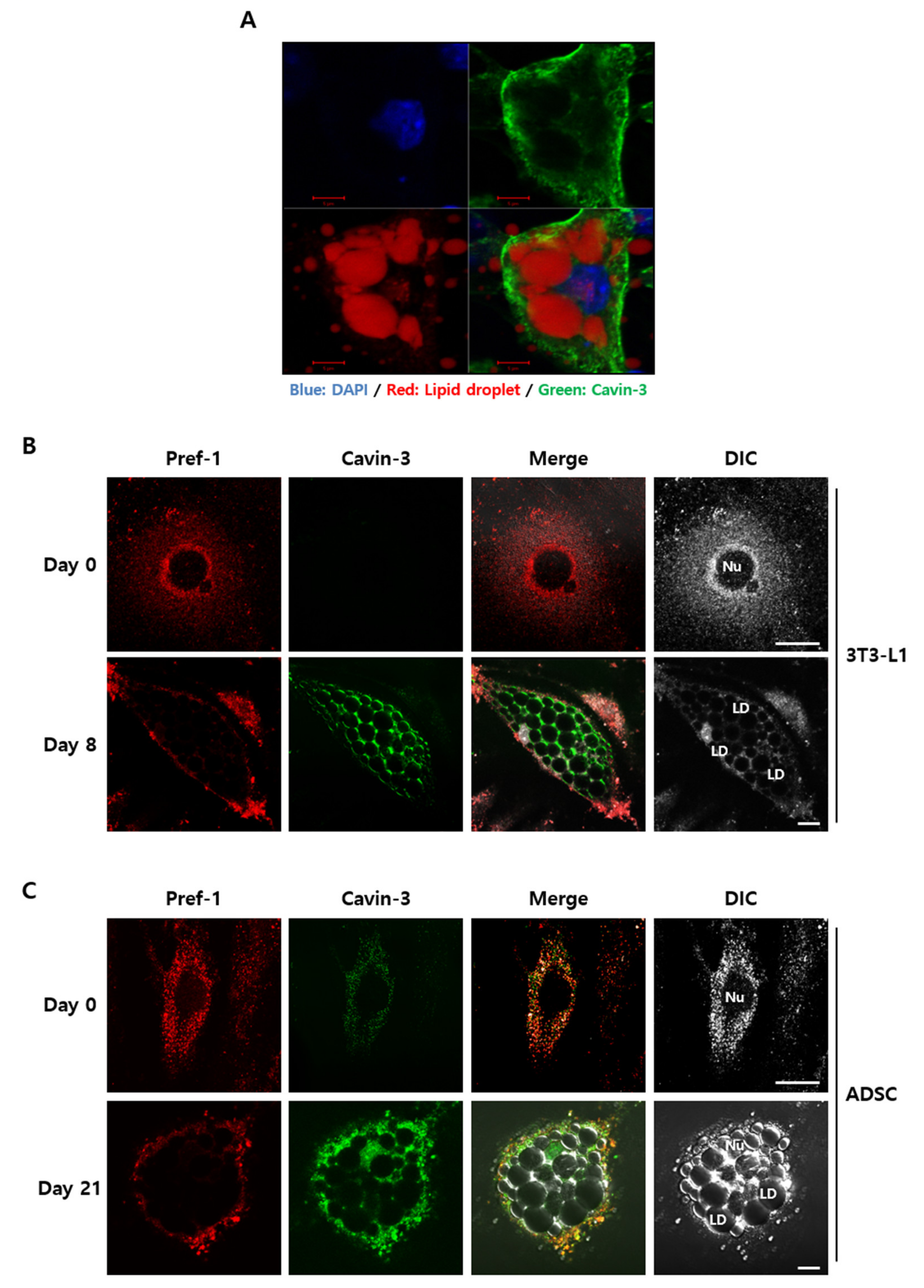
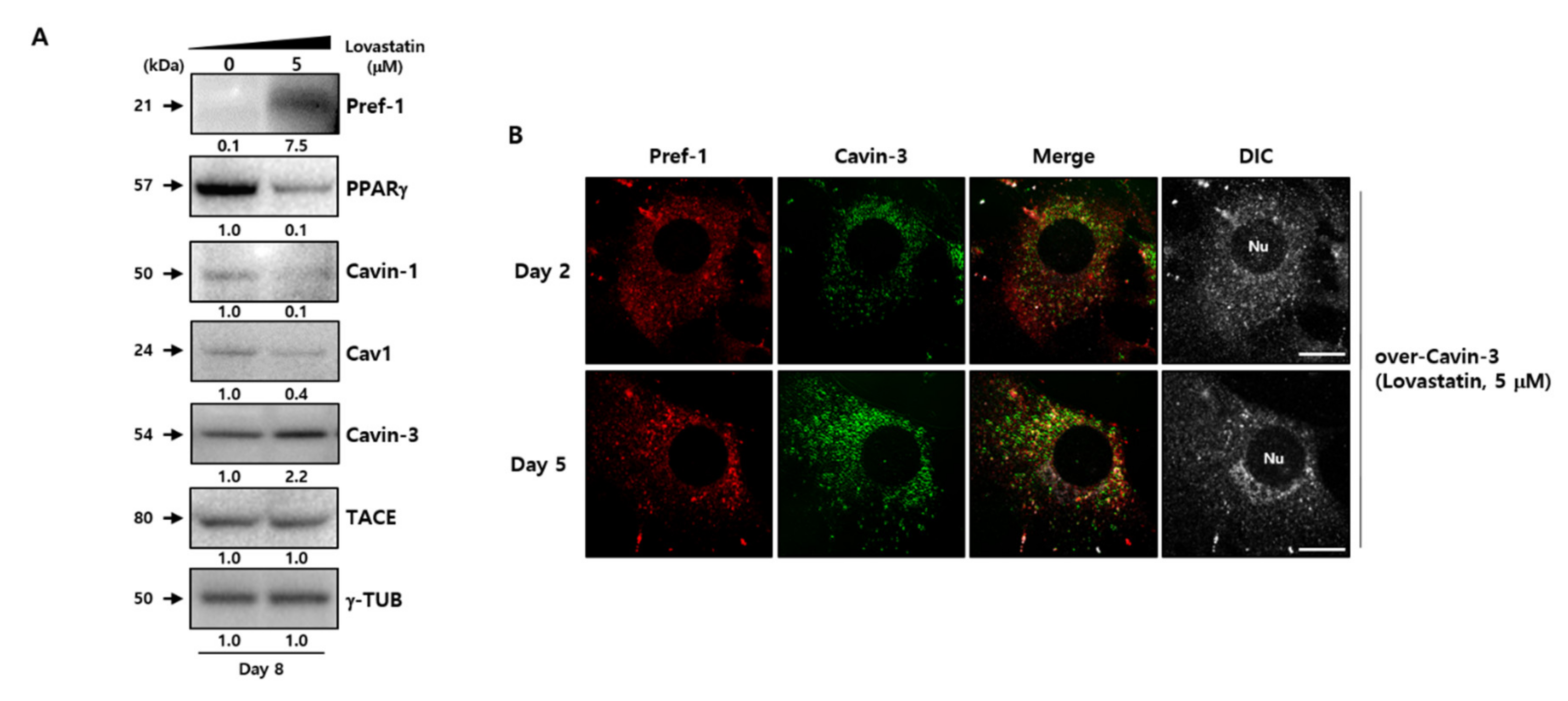
2.4. Localization of Cavin-3 Might Affect TACE-Mediated Pref-1 Shedding
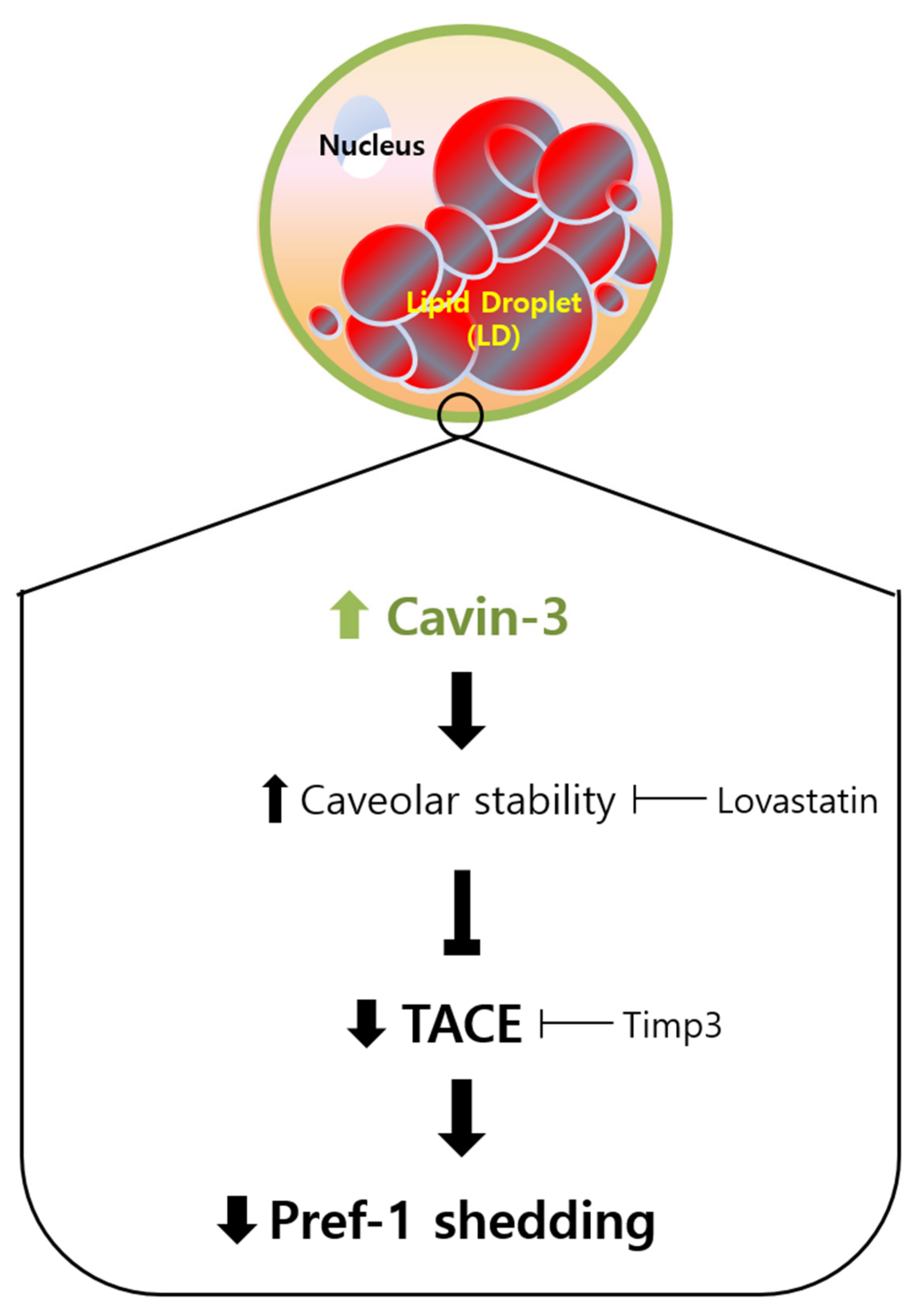
2.5. Cavin-3 Supports Caveolar Stability to Inhibit the Translocation of Pref-1 Small Fragment into Cytosol
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Differentiation
4.2. Chemicals
4.3. RNA Extraction and RT-qPCR
4.4. RNAi Experiment
4.5. Overexpression Experiment
4.6. Oil Red O Staining
4.7. Analysis of TG Content
4.8. Western Blot Analysis
4.9. Confocal Immunofluorescence Microscopy
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADSC | Human adipose derived stem cells |
| aP2 | Adipocyte protein 2 |
| Cavin-3 | Caveolae-associated protein-3 |
| siCavin-3 | Cavin-3 knockdown stable cell lines |
| over-Cavin-3 | Cavin-3 overexpression stable cell lines |
| C/EBPβ | CCAAT/enhancer-binding protein beta |
| C/EBPδ | CCAAT/enhancer-binding protein delta |
| FAS | Fatty acid synthase |
| ORO | Oil Red O |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| Pref-1 | Preadipocyte factor 1 |
| RNAi | RNA interference |
| RT-qPCR | Real-time quantitative polymerase chain reaction |
| SCD-1 | Stearoyl-CoA desaturase |
| TACE | Tumor necrosis factor-alpha-converting enzyme |
| Timp3 | Tissue inhibitor of metalloproteinase 3 |
| TG | Total triacylglycerol |
References
- Chao, L.; Marcus-Samuels, B.; Mason, M.M.; Moitra, J.; Vinson, C.; Arioglu, E.; Gavrilova, O.; Reitman, M.L. Adipose tissue is required for the antidiabetic, but not for the hypolipidemic, effect of thiazolidinediones. J. Clin. Investig. 2000, 106, 1221–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Takahata, T.; Kumano, T.; Ookawa, K.; Hayakari, M.; Kakizaki, I.; Tsuchida, S. Inhibition of 3T3-L1 adipocyte differentiation by 6-ethoxyzolamide: Repressed peroxisome proliferator-activated receptor gamma mRNA and enhanced CCAAT/enhancer binding protein beta mRNA levels. Biochem. Pharmacol. 2004, 67, 1667–1675. [Google Scholar] [CrossRef]
- Koutnikova, H.; Auwerx, J. Regulation of adipocyte differentiation. Ann. Med. 2001, 33, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M. Obesity in the new millennium. Nature 2000, 404, 632–634. [Google Scholar] [CrossRef] [PubMed]
- Dizdar, O.; Alyamaç, E. Obesity: An endocrine tumor? Med. Hypotheses 2004, 63, 790–792. [Google Scholar] [CrossRef]
- Smas, C.M.; Sul, H.S. Pref-1, a protein containing EGF-like repeats, inhibits adipocyte differentiation. Cell 1993, 73, 725–734. [Google Scholar] [CrossRef]
- Sul, H.S.; Smas, C.; Mei, B.; Zhou, L. Function of pref-1 as an inhibitor of adipocyte differentiation. Int. J. Obes. Relat. Metab. Disord. 2000, 24, S15–S19. [Google Scholar] [CrossRef] [Green Version]
- Mei, B.; Zhao, L.; Chen, L.; Sul, H.S. Only the large soluble form of preadipocyte factor-1 (Pref-1), but not the small soluble and membrane forms, inhibits adipocyte differentiation: Role of alternative splicing. Biochem. J. 2002, 364, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Kim, K.A.; Kim, J.H.; Sul, H.S. Pref-1, a preadipocyte secreted factor that inhibits adipogenesis. J. Nutr. 2006, 136, 2953–2956. [Google Scholar] [CrossRef]
- Smas, C.; Chen, L.; Sul, H.S. Cleavage of membrane-associated pref-1 generates a soluble inhibitor of adipocyte differentiation. Mol. Cell. Biol. 1997, 17, 977–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Villena, J.A.; Moon, Y.S.; Kim, K.H.; Lee, S.; Kang, C.; Sul, H.S. Inhibition of adipogenesis and development of glucose intolerance by soluble preadipocyte factor-1 (Pref-1). J. Clin. Investig. 2003, 111, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Sul, H.S. Ectodomain shedding of preadipocyte factor 1 (Pref-1) by tumor necrosis factor alpha converting enzyme (TACE) and inhibition of adipocyte differentiation. Mol. Cell. Biol. 2006, 26, 5421–5435. [Google Scholar] [CrossRef] [Green Version]
- Sul, H.S. Minireview: Pref-1: Role in adipogenesis and mesenchymal cell fate. Mol. Endocrinol. 2009, 23, 1717–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unger, R.H. Minireview: Weapons of lean body mass destruction: The role of ectopic lipids in the metabolic syndrome. Endocrinology 2003, 144, 5159–5165. [Google Scholar] [CrossRef] [Green Version]
- Briand, N.; Le Lay, S.; Sessa, W.C.; Ferré, P.; Dugail, I. Distinct roles of endothelial and adipocyte caveolin-1 in macrophage infiltration and adipose tissue metabolic activity. Diabetes 2011, 60, 448–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastiani, M.; Parton, R.G. Caveolae at a glance. J. Cell Sci. 2010, 123, 3831–3836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, K.; Gerl, M.J. Revitalizing membrane rafts: New tools and insights. Nat. Rev. Mol. Cell Biol. 2010, 11, 688–699. [Google Scholar] [CrossRef]
- Pilch, P.F.; Meshulam, T.; Ding, S.; Liu, L. Caveolae and lipid trafficking in adipocytes. Clin. Lipidol. 2011, 6, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.W.; Razani, B.; Wang, X.B.; Combs, T.P.; Williams, T.M.; Scherer, P.E.; Lisanti, M.P. Caveolin-1-deficient mice show insulin resistance and defective insulin receptor protein expression in adipose tissue. Am. J. Physiol. Cell Physiol. 2003, 285, C222–C235. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Alston, L.; Ruschman, J.; Hegele, R.A. Heterozygous CAV1 frame shift mutations (MIM 601047) in patients with atypical partial lipodystrophy and hypertriglyceridemia. Lipids Health Dis. 2008, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.A.; Delépine, M.; Boutet, E.; Mourabit, H.; Le Lay, S.; Meier, M.; Nemani, M.; Bridel, E.E.; Leite, C.C.; Bertola, D.R.; et al. Association of a homozygous nonsense caveolin-1 mutation with Berardinelli-Seip congenital lipodystrophy. J. Clin. Endocrinol. Metab. 2008, 93, 1129–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briand, N.; Prado, C.; Mabilleau, G.; Lasnier, F.; Le Lièpvre, X.; Covington, J.D.; Ravussin, E.; Le Lay, S.; Dugail, I. Caveolin-1 expression and cavin stability regulate caveolae dynamics in adipocyte lipid store fluctuation. Diabetes 2014, 63, 4032–4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, Y.; Hirai, S.; Tamai, Y.; Fujise-Matsuoka, A.; Nishimura, Y.; Ohno, S. A protein kinase C delta-binding protein SRBC whose expression is induced by serum starvation. J. Biol. Chem. 1997, 272, 7381–7389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansa, P.; Mason, S.W.; Hoffmann-Rohrer, U.; Grummt, I. Cloning and functional characterization of PTRF, a novel protein which induces dissociation of paused ternary transcription complexes. EMBO J. 1998, 17, 2855–2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, K.A.; Zajicek, H.; Li, W.P.; Peyton, M.J.; Minna, J.D.; Hernandez, V.J.; Luby-Phelps, K.; Anderson, R.G. SRBC/cavin-3 is a caveolin adapter protein that regulates caveolae function. EMBO J. 2009, 28, 1001–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.L.; Wu, L.C.; Du, F.; Davis, A.; Peyton, M.; Tomizawa, Y.; Maitra, A.; Tomlinson, G.; Gazdar, A.F.; Weissman, B.E.; et al. Inactivation of human SRBC, located within the 11p15.5-p15.4 tumor suppressor region, in breast and lung cancers. Cancer Res. 2001, 61, 7943–7949. [Google Scholar]
- Zöchbauer-Müller, S.; Fong, K.M.; Geradts, J.; Xu, X.; Seidl, S.; End-Pfützenreuter, A.; Lang, G.; Heller, G.; Zielinski, C.C.; Gazdar, A.F.; et al. Expression of the candidate tumor suppressor gene hSRBC is frequently lost in primary lung cancers with and without DNA methylation. Oncogene 2005, 24, 6249–6255. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Byun, D.S.; Lee, M.G.; Ryu, B.K.; Kang, M.J.; Chae, K.S.; Lee, K.Y.; Kim, H.J.; Park, H.; Chi, S.G. Frequent epigenetic inactivation of hSRBC in gastric cancer and its implication in attenuated p53 response to stresses. Int. J. Cancer 2008, 122, 1573–1584. [Google Scholar] [CrossRef]
- Bastiani, M.; Liu, L.; Hill, M.M.; Jedrychowski, M.P.; Nixon, S.J.; Lo, H.P.; Abankwa, D.; Luetterforst, R.; Fernandez-Rojo, M.; Breen, M.R.; et al. MURC/Cavin-4 and cavin family members form tissue-specific caveolar complexes. J. Cell Biol. 2009, 185, 1259–1273. [Google Scholar] [CrossRef]
- Hernandez, V.J.; Weng, J.; Ly, P.; Pompey, S.; Dong, H.; Mishra, L.; Schwarz, M.; Anderson, R.G.; Michaely, P. Cavin-3 dictates the balance between ERK and Akt signaling. eLife 2013, 2, e00905. [Google Scholar] [CrossRef]
- Liu, L.; Hansen, C.G.; Honeyman, B.J.; Nichols, B.J.; Pilch, P.F. Cavin-3 knockout mice show that cavin-3 is not essential for caveolae formation, for maintenance of body composition, or for glucose tolerance. PLoS ONE 2014, 9, e102935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, S.R. Regulation of PPARgamma activity during adipogenesis. Int. J. Obes. 2005, 29, S13–S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafontan, M.; Viguerie, N. Role of adipokines in the control of energy metabolism: Focus on adiponectin. Curr. Opin. Pharmacol. 2006, 6, 580–585. [Google Scholar] [CrossRef]
- Smith, S.; Witkowski, A.; Joshi, A.K. Structural and functional organization of the animal fatty acid synthase. Prog. Lipid Res. 2003, 42, 289–317. [Google Scholar] [CrossRef]
- Sun, L.; Nicholson, A.C.; Hajjar, D.P.; Gotto, A.M.; Han, J.H. Adipogenic differentiating agents regulate expression of fatty acid binding protein and CD36 in the J744 macrophage cell line. J. Lipid Res. 2003, 44, 1877–1886. [Google Scholar] [CrossRef] [Green Version]
- Yao-Borengasser, A.; Rassouli, N.; Varma, V.; Bodles, A.M.; Rasouli, N.; Unal, R.; Phanavanh, B.; Ranganathan, G.; McGehee, R.E., Jr.; Kern, P.A. Stearoyl-coenzyme A desaturase 1 gene expression increases after pioglitazone treatment and is associated with peroxisomal proliferator-activated receptor-γ responsiveness. J. Clin. Endocrinol. Metab. 2008, 93, 4431–4439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amour, A.; Slocombe, P.M.; Webster, A.; Butler, M.; Knight, C.G.; Smith, B.J.; Stephens, P.E.; Shelley, C.; Hutton, M.; Knäuper, V.; et al. TNF-alpha converting enzyme (TACE) is inhibited by TIMP-3. FEBS Lett. 1998, 435, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Fréour, T.; Jarry, A.; Bach-Ngohou, K.; Dejoie, T.; Bou-Hanna, C.; Denis, M.G.; Mosnier, J.F.; Laboisse, C.L.; Masson, D. TACE inhibition amplifies TNF-alpha-mediated colonic epithelial barrier disruption. Int. J. Mol. Med. 2009, 23, 41–48. [Google Scholar]
- Mohan, J.; Morén, B.; Larsson, E.; Holst, M.R.; Lundmark, R. Cavin3 interacts with cavin1 and caveolin1 to increase surface dynamics of caveolae. J. Cell Sci. 2015, 128, 979–991. [Google Scholar] [CrossRef] [Green Version]
- Prusty, D.; Park, B.H.; Davis, K.E.; Farmer, S.R. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor gamma (PPARγ) and C/EBPα gene expression during the differentiation of 3T3-L1 preadipocytes. J. Biol. Chem. 2002, 277, 46226–46232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.; Pietiaïnen, V.M.; Pölönen, H.; Rasilainen, L.; Koivusalo, M.; Ruotsalainen, U.; Jokitalo, E.; Ikonen, E. Cholesterol substitution increases the structural heterogeneity of caveolae. J. Biol. Chem. 2008, 283, 14610–14618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pengde, K.; Fuxing, P.; Bin, S.; Jing, Y.; Jingqiu, C. Lovastatin inhibits adipogenesis and prevents osteonecrosis in steroid-treated rabbits. Jt. Bone Spine 2008, 75, 696–701. [Google Scholar] [CrossRef]
- Ramji, D.P.; Foka, P. CCAAT/enhancer-binding proteins: Structure, function and regulation. Biochem. J. 2002, 365, 561–575. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, P.J.; Kim, S.T. Caveolae-Associated Protein 3 (Cavin-3) Influences Adipogenesis via TACE-Mediated Pref-1 Shedding. Int. J. Mol. Sci. 2020, 21, 5000. https://doi.org/10.3390/ijms21145000
Park PJ, Kim ST. Caveolae-Associated Protein 3 (Cavin-3) Influences Adipogenesis via TACE-Mediated Pref-1 Shedding. International Journal of Molecular Sciences. 2020; 21(14):5000. https://doi.org/10.3390/ijms21145000
Chicago/Turabian StylePark, Phil June, and Sung Tae Kim. 2020. "Caveolae-Associated Protein 3 (Cavin-3) Influences Adipogenesis via TACE-Mediated Pref-1 Shedding" International Journal of Molecular Sciences 21, no. 14: 5000. https://doi.org/10.3390/ijms21145000
APA StylePark, P. J., & Kim, S. T. (2020). Caveolae-Associated Protein 3 (Cavin-3) Influences Adipogenesis via TACE-Mediated Pref-1 Shedding. International Journal of Molecular Sciences, 21(14), 5000. https://doi.org/10.3390/ijms21145000