Gingival Crevicular Fluid Peptidome Profiling in Healthy and in Periodontal Diseases


 and
and 
Abstract
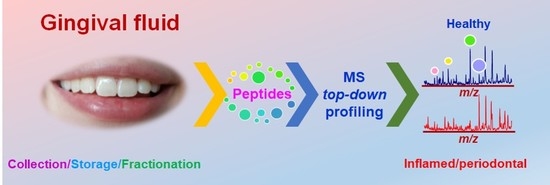
:
1. Introduction
1.1. Gingivitis and Periodontitis
1.2. Nature and Function of GCF
1.3. Low Molecular Weight Profiling of GCF
2. An Overview of MS Profiling Tools and Investigations to Detect GCF Peptides
2.1. SELDI MS GCF Peptidome Profiling
2.2. MALDI MS GCF Peptidome Profiling
2.3. ESI MS GCF Peptidome Profiling
3. The GCF Antimicrobial Peptidome
4. Relevance and Challenge of Pre-Analytical and Analytical Variables
4.1. Quantitation Issues
4.2. PIC
4.3. Storage
4.4. The Inter-Individual Variability
5. Bioinformatics Approaches
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tonetti, M.S.; Jepsen, S.; Jin, L.; Otomo-Corgel, J. Impact of the global burden of periodontal diseases on health, nutrition and wellbeing of mankind: A call for global action. J. Clin. Periodontol. 2017, 44, 456–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonetti, M.S.; Greenwell, H.; Kornman, K.S. Staging and Grading of Periodontitis: Framework and Proposal of a New Classification and Case Definition. J. Periodontol. 2018, 89, 159–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchida, S.; Satoh, M.; Sogawa, K.; Kawashima, Y.; Kado, S.; Ishige, T.; Beppu, M.; Sawai, S.; Nishimura, M.; Kodera, Y.; et al. Application of Proteomic Technologies to Discover and Identify Biomarkers for Periodontal Diseases in Gingival Crevicular Fluid: A review. Proteom. Clin. Appl. 2014, 8, 232–240. [Google Scholar] [CrossRef]
- Trindade, F.; Oppenheim, F.G.; Helmerhorst, E.J.; Amado, F.; Gomes, P.S.; Vitorino, R. Uncovering the Molecular Networks in Periodontitis. Proteom. Clin. Appl. 2014, 8, 748–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.C.; Schroeder, H.E. Pathogenesis of Inflammatory Periodontal Disease. A Summary of Current Work. Lab. Investig. J. Tech. Methods Pathol. 1976, 34, 235–249. [Google Scholar]
- Ngo, L.H.; Veith, P.D.; Chen, Y.Y.; Chen, D.; Darby, I.B.; Reynolds, E.C. Mass Spectrometric Analyses of Peptides and Proteins in Human Gingival Crevicular Fluid. J. Proteome Res. 2010, 9, 1683–1693. [Google Scholar] [CrossRef]
- Toczewska, J.; Konopka, T.; Zalewska, A.; Maciejczyk, M. Nitrosative Stress Biomarkers in the Non-Stimulated and Stimulated Saliva, as well as Gingival Crevicular Fluid of Patients with Periodontitis: Review and Clinical Study. Antioxidants 2020, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Khurshid, Z.; Mali, M.; Naseem, M.; Najeeb, S.; Zafar, M.S. Human Gingival Crevicular Fluids (GCF) Proteomics: An Overview. Dent. J. 2017, 5, 12. [Google Scholar] [CrossRef]
- Barros, S.P.; Williams, R.; Offenbacher, S.; Morelli, T. Gingival Crevicular as a Source of Biomarkers for Periodontitis. Periodontology 2000 2016, 70, 53–64. [Google Scholar] [CrossRef]
- Delima, A.J.; Van Dyke, T.E. Origin and Function of the Cellular Components in Gingival Crevice Fluid. Periodontology 2000 2003, 31, 55–76. [Google Scholar] [CrossRef]
- Subbarao, K.C.; Nattuthurai, G.S.; Sundararajan, S.K.; Sujith, I.; Joseph, J.; Syedshah, Y.P. Gingival Crevicular Fluid: An Overview. J. Pharm. Bioallied Sci. 2019, 11, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Majeed, Z.N.; Philip, K.; Alabsi, A.M.; Pushparajan, S.; Swaminathan, D. Identification of Gingival Crevicular Fluid Sampling, Analytical Methods, and Oral Biomarkers for the Diagnosis and Monitoring of Periodontal Diseases: A Systematic Review. Dis. Markers 2016, 2016, 1804727. [Google Scholar]
- Griffiths, G.S. Formation, Collection and Significance of Gingival Crevice Fluid. Periodontology 2000 2003, 31, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Silva-Boghossian, C.M.; Colombo, A.P.; Tanaka, M.; Rayo, C.; Xiao, Y.; Siqueira, W.L. Quantitative Proteomic Analysis of Gingival Crevicular Fluid in Different Periodontal Conditions. PLoS ONE 2013, 8, e75898. [Google Scholar] [CrossRef] [Green Version]
- Bostanci, N.; Bao, K. Contribution of Proteomics to Our Understanding of Periodontal Inflammation. Proteomics 2017, 17, 1500518. [Google Scholar] [CrossRef]
- Lundy, F.T.; Chalk, R.; Lamey, P.J.; Shaw, C.; Linden, G.J. Quantitative Analysis of MRP-8 in Gingival Crevicular Fluid in Periodontal Health and Disease Using Microbore HPLC. J. Clin. Periodontol. 2001, 28, 1172–1177. [Google Scholar] [CrossRef]
- Dommisch, H.; Vorderwülbecke, S.; Eberhard, J.; Steglich, M.; Jepsen, S. SELDI-TOF-MS of Gingival Crevicular Fluid-A Methodological Approach. Arch. Oral Biol. 2009, 54, 803–809. [Google Scholar] [CrossRef]
- Diamond, D.L.; Kimball, J.R.; Krisanaprakornkit, S.; Ganz, T.; Dale, B.A. Detection of Beta-Defensins Secreted by Human Oral Epithelial Cells. J. Immunol. Methods 2001, 256, 65–76. [Google Scholar] [CrossRef]
- Lundy, F.T.; Orr, D.F.; Shaw, C.; Lamey, P.J.; Linden, G.J. Detection of Individual Human Neutrophil Alpha-Defensins (Human Neutrophil Peptides 1, 2 and 3) in Unfractionated Gingival Crevicular Fluid-A MALDI-MS Approach. Mol. Immunol. 2005, 42, 575–579. [Google Scholar] [CrossRef]
- Wen, X.; Gu, Y.; Chen, F. Gingival Crevicular Fluid as a Novel Potential Source of Biomarkers Distinguishes Pubertal from Post-Pubertal Subjects. Diagnostics 2016, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Ngo, L.H.; Darby, I.B.; Veith, P.D.; Locke, A.G.; Reynolds, E.C. Mass Spectrometric Analysis of Gingival Crevicular Fluid Biomarkers Can Predict Periodontal Disease Progression. J. Periodontal Res. 2013, 48, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Preianò, M.; Falcone, D.; Maggisano, G.; Montalcini, T.; Navarra, M.; Paduano, S.; Savino, R.; Terracciano, R. Assessment of Pre-Analytical and Analytical Variables Affecting Peptidome Profiling of Gingival Crevicular Fluid by MALDI-TOF Mass Spectrometry. Clin. Chim. Acta 2014, 437, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Preianò, M.; Maggisano, G.; Lombardo, N.; Montalcini, T.; Paduano, S.; Pelaia, G.; Savino, R.; Terracciano, R. Influence of Storage Conditions on MALDI-TOF MS Profiling of Gingival Crevicular Fluid: Implications on the Role of S100A8 and S100A9 for Clinical and Proteomic Based Diagnostic Investigations. Proteomics 2016, 16, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Preianò, M.; Maggisano, G.; Murfuni, M.S.; Villella, C.; Pelaia, C.; Montalcini, T.; Lombardo, N.; Pelaia, G.; Savino, R.; Terracciano, R. An Analytical Method for Assessing Optimal Storage Conditions of Gingival Crevicular Fluid and Disclosing a Peptide Biomarker Signature of Gingivitis by MALDI-TOF MS. Proteom. Clin. Appl. 2018, 12, e1800005. [Google Scholar] [CrossRef]
- Tang, H.; Yuan, C.; Ma, Z.; Zhu, C.; Tong, P.; Gallagher, J.E.; Sun, X.; Zheng, S. The Potentiality of Salivary Peptide Biomarkers for Screening Patients with Periodontal Diseases by Mass Spectrometry. Clin. Chim. Acta 2019, 495, 278–286. [Google Scholar] [CrossRef]
- Antezack, A.; Chaudet, H.; Tissot-Dupont, H.; Brouqui, P.; Monnet-Corti, V. Rapid Diagnosis of Periodontitis, a Feasibility Study Using MALDI-TOF Mass Spectrometry. PLoS ONE 2020, 15, e0230334. [Google Scholar] [CrossRef]
- Pisano, E.; Cabras, T.; Montaldo, C.; Piras, V.; Inzitari, R.; Olmi, C.; Castagnola, M.; Messana, I. Peptides of Human Gingival Crevicular Fluid Determined by HPLC-ESI-MS. Eur. J. Oral. Sci. 2005, 113, 462–468. [Google Scholar] [CrossRef]
- Inzitari, R.; Cabras, T.; Pisano, E.; Fanali, C.; Manconi, B.; Scarano, E.; Fiorita, A.; Paludetti, G.; Manni, A.; Nemolato, S.; et al. HPLC-ESI-MS analysis of oral human fluids reveals that gingival crevicular fluid is the main source of oral thymosins beta (4) and beta (10). J. Sep. Sci. 2009, 32, 57–63. [Google Scholar] [CrossRef]
- Dassatti, L.; Manicone, P.F.; Iavarone, F.; Stefanelli, N.; Nicoletti, F.; Lazzareschi, I.; Luciano, R.P.M.; Castagnola, M.; D’Addona, A. Proteomic Evaluation of GCF in the Development of Pregnancy Related Periodontal Disease: A Pilot Clinical Study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5030–5039. [Google Scholar]
- Grant, M.M.; Creese, A.J.; Barr, G.; Ling, M.R.; Scott, A.E.; Matthews, J.B.; Griffiths, H.R.; Cooper, H.J.; Chapple, I.L. Proteomic Analysis of a Noninvasive Human Model of Acute Inflammation and Its Resolution: The Twenty-One Day Gingivitis Model. J. Proteome Res. 2010, 9, 4732–4744. [Google Scholar] [CrossRef] [Green Version]
- Kido, J.; Bando, M.; Hiroshima, Y.; Iwasaka, H.; Yamada, K.; Ohgami, N.; Nambu, T.; Kataoka, M.; Yamamoto, T.; Shinohara, Y.; et al. Analysis of Proteins in Human Gingival Crevicular Fluid by Mass Spectrometry. J. Periodontal Res. 2012, 47, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, S.; Satoh, M.; Umemura, H.; Sogawa, K.; Kawashima, Y.; Kado, S.; Sawai, S.; Nishimura, M.; Kodera, Y.; Matsushita, K.; et al. Proteomic Analysis of Gingival Crevicular Fluid for Discovery of Novel Periodontal Disease Markers. Proteomics 2012, 12, 2190–2202. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, S.; Satoh, M.; Kawashima, Y.; Sogawa, K.; Kado, S.; Sawai, S.; Nishimura, M.; Ogita, M.; Takeuchi, Y.; Kobyashi, H.; et al. Application of Quantitative Proteomic Analysis Using Tandem Mass Tags for Discovery and Identification of Novel Biomarkers in Periodontal Disease. Proteomics 2013, 13, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Baliban, R.C.; Sakellari, D.; Zukui, L.; Li, Z.; DiMaggio, P.A.; Garcia, B.A.; Floudas, C.A. Novel Protein Identification Methods for Biomarker Discovery via a Proteomic Analysis of Periodontally Healthy and Diseased Gingival Crevicular Fluid Samples. J. Clin. Periodontol. 2012, 39, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baliban, R.C.; Sakellari, D.; Li, Z.; Guzman, Y.A.; Garcia, B.A.; Floudas, C.A. Discovery of biomarker combinations that predict periodontal health or disease with high accuracy from GCF samples based on high-throughput proteomic analysis and mixed-integer linear optimization. J. Clin. Periodontol. 2013, 40, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.J.; Heo, S.H.; Lee, J.M.; Cho, J.Y. Identification of Azurocidin as a Potential Periodontitis Biomarker by a Proteomic Analysis of Gingival Crevicular Fluid. Proteome Sci. 2011, 28. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, L.G.; Venuleo, C.; Oppenheim, F.G.; Salih, E. Proteome Data Set of Human Gingival Crevicular Fluid From Healthy Periodontium Sites by Multidimensional Protein Separation and Mass Spectrometry. J. Periodontal Res. 2012, 47, 248–262. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, L.G.; Nouh, H.; Salih, E. Quantitative Gingival Crevicular Fluid Proteome in Health and Periodontal Disease Using Stable Isotope Chemistries and Mass Spectrometry. J. Clin. Periodontol. 2014, 41, 733–747. [Google Scholar] [CrossRef] [Green Version]
- Bostanci, N.; Heywood, W.; Mills, K.; Parkar, M.; Nibali, L.; Donos, N. Application of Label-Free Absolute Quantitative Proteomics in Human Gingival Crevicular Fluid by LC/MS E (Gingival Exudatome). J. Proteome Res. 2010, 9, 2191–2199. [Google Scholar] [CrossRef] [Green Version]
- Bostanci, N.; Ramberg, P.; Wahlander, Å.; Grossman, J.; Jönsson, D.; Barnes, V.M.; Papapanou, P.N. Label-free quantitative proteomics reveals differentially regulated proteins in experimental gingivitis. J. Proteome Res. 2013, 12, 657–678. [Google Scholar] [CrossRef]
- Huynh, A.H.; Veith, P.D.; McGregor, N.R.; Adams, G.G.; Chen, D.; Reynolds, E.C.; Ngo, L.H.; Darby, I.B. Gingival Crevicular Fluid Proteomes in Health, Gingivitis and Chronic Periodontitis. J. Periodontal Res. 2015, 50, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Guzman, Y.A.; Sakellari, D.; Papadimitriou, K.; Floudas, C.A. High-throughput proteomic analysis of candidate biomarker changes in gingival crevicular fluid after treatment of chronic periodontitis. J. Periodontal Res. 2018, 53, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Batschkus, S.; Cingoez, G.; Urlaub, H.; Miosge, N.; Kirschneck, C.; Meyer-Marcotty, P.; Lenz, C. A new albumin-depletion strategy improves proteomic research of gingival crevicular flflfluid from periodontitis patients. Clin. Oral Investig. 2017, 22, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Marinho, M.C.; Pacheco, A.; Costa, G.; Ortiz, N.D.; Zajdenverg, L.; Sansone, C. Quantitative gingival crevicular fluid proteome in type 2 diabetes mellitus and chronic periodontitis. Oral Dis. 2019, 25, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Silva-Boghossian, C.M.; Colombo, A.P.; Tanaka, M.; Rayo, C.; Xiao, Y.; Siqueira, W.L. Quantitative proteomic analysis of gingival crevicular flflfluids from deciduous and permanent teeth. J. Clin. Periodontol. 2017, 44, 353–362. [Google Scholar]
- Odanaka, H.; Obama, T.; Sawada, N.; Sugano, M.; Itabe, H.; Yamamoto, M. Comparison of protein profiles of the pellicle, gingival crevicular fluid, and saliva: Possible origin of pellicle proteins. Biol. Res. 2020, 53, 3. [Google Scholar] [CrossRef]
- Colares, V.; Lima, S.; Sousa, N.; Araújo, M.C.; Pereira, D.; Mendes, S.; Teixeira, S.A.; Monteiro, C.A.; Bandeca, M.C.; Siqueira, W.L.; et al. Hydrogen peroxide-based products alter inflammatory and tissue damage-related proteins in the gingival crevicular fluid of healthy volunteers: A randomized trial. Sci. Rep. 2019, 9, 3457. [Google Scholar] [CrossRef]
- Bostanci, N.; Belibasakis, G.N. Gingival crevicular fluid and its immune mediators in the proteomic era. Periodontology 2000 2018, 76, 68–84. [Google Scholar] [CrossRef]
- Tsuchida, S.; Satoh, M.; Takiwaki, M.; Nomura, F. Current Status of Proteomic Technologies for Discovering and Identifying Gingival Crevicular Fluid Biomarkers for Periodontal Disease. Int. J. Mol. Sci. 2018, 20, 86. [Google Scholar] [CrossRef] [Green Version]
- Hutchens, T.W.; Yip, T.T. New desorption strategies for the mass spectrometric analysis of macromolecules. Rapid Commun. Mass Spectrom. 1993, 7, 576–580. [Google Scholar] [CrossRef]
- Pitteri, S.J.; Hanash, S.M. Proteomic approaches for cancer biomarker discovery in plasma. Expert Rev. Proteom. 2007, 4, 589–590. [Google Scholar] [CrossRef] [PubMed]
- Hodgetts, A.; Levin, M.; Kroll, J.S.; Langford, P.R. Biomarker discovery in infectious diseases using SELDI. Future Microbiol. 2007, 2, 35–49. [Google Scholar] [CrossRef]
- Whelan, L.C.; Power, K.A.; McDowell, D.T.; Kennedy, J.; Gallagher, W.M. Applications of SELDI-MS technology in oncology. J. Cell. Mol. Med. 2008, 12, 1535–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamandis, E.P. Mass spectrometry as a diagnostic and a cancer biomarker discovery tool: Opportunities and potential limitations. Mol. Cell. Proteom. 2004, 3, 367–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wei, D.; Yap, Y.; Li, L.; Guo, S.; Chen, F. Mass spectrometry-based “omics” technologies in cancer diagnostics. Mass Spectrom. Rev. 2007, 26, 403–431. [Google Scholar] [CrossRef]
- Semmes, O.J.; Feng, Z.; Adam, B.L.; Banez, L.L.; Bigbee, W.L.; Campos, D.; Cazares, L.H.; Chan, D.W.; Grizzle, W.E.; Izbicka, E.; et al. Evaluation of serum protein profiling by surface-enhanced laser desorption/ionization time-of-flight mass spectrometry for the detection of prostate cancer: I. Assessment of platform reproducibility. Clin. Chem. 2005, 51, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Savino, R.; Terracciano, R. Mesopore-assisted profiling strategies in clinical proteomics for drug/target discovery. Drug Discov. Today 2012, 17, 143–152. [Google Scholar] [CrossRef]
- Mann, M.; Hendrickson, R.C.; Pandey, A. Analysis of proteins and proteomes by mass spectrometry. Annu. Rev. Biochem. 2001, 70, 437–473. [Google Scholar] [CrossRef]
- Cho, Y.T.; Su, H.; Huang, T.L.; Chen, H.C.; Wu, W.J.; Wu, P.C.; Wu, D.C.; Shiea, J. Matrix-assisted laser desorption ionization/time-of-flight mass spectrometry for clinical diagnosis. Clin. Chim. Acta 2013, 415, 266–275. [Google Scholar] [CrossRef]
- Terracciano, R.; Pasqua, L.; Casadonte, F.; Frascà, S.; Preianò, M.; Falcone, D.; Savino, R. Derivatized mesoporous silica beads for MALDI-TOF MS profiling of human plasma and urine. Bioconjug. Chem. 2009, 20, 913–923. [Google Scholar] [CrossRef]
- Savino, R.; Paduano, S.; Preianò, M.; Terracciano, R. The proteomics big challenge for biomarkers and new drug-targets discovery. Int. J. Mol. Sci. 2012, 13, 13926–13948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, V.; Piras, C.; Pieroni, L.; Ronci, M.; Putignani, L.; Roncada, P.; Urbani, A. Applications of MALDI-TOF mass spectrometry in clinical proteomics. Expert Rev. Proteom. 2018, 15, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, R.; Casadonte, F.; Pasqua, L.; Candeloro, P.; Di Fabrizio, E.; Urbani, A.; Savino, R. Enhancing plasma peptide MALDI-TOF-MS profiling by mesoporous silica assisted crystallization. Talanta 2010, 80, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi-Ishii, Y.; Hasebe, T.; Nagaoka, I. Role of CCAAT/enhancer-binding protein site in transcription of human neutrophil peptide-1 and -3 defensin genes. J. Immunol. 2000, 164, 3264–3273. [Google Scholar] [CrossRef] [Green Version]
- Gibb, S.; Strimmer, K. Differential protein expression and peak selection in mass spectrometry data by binary discriminant analysis. Bioinformatics 2015, 31, 3156–3162. [Google Scholar] [CrossRef] [Green Version]
- Bellagambi, F.G.; Degano, I.; Ghimenti, S.; Lomonaco, T.; Dini, V.; Romanelli, M.; Mastorci, F.; Gemignani, A.; Salvo, P.; Fuoco, R.; et al. Determination of salivary α-amylase and cortisol in psoriatic subjects undergoing the Trier Social Stress Test. Microchem. J. 2018, 136, 177–184. [Google Scholar] [CrossRef]
- Lomonaco, T.; Ghimenti, S.; Biagini, D.; Bramanti, E.; Onor, M.; Bellagambi, F.G.; Fuoco, R.; Di Francesco, F. The effect of sampling procedures on the urate in oral fluid and lactate concentration in oral fluid. Microchem. J. 2018, 136, 255–262. [Google Scholar] [CrossRef]
- Aebersold, R.; Mann, M. Mass spectrometry-based proteomics. Nature 2003, 422, 198–207. [Google Scholar] [CrossRef]
- Whitehouse, C.M.; Dreyer, R.N.; Yamashita, M.; Fenn, J.B. Electrospray interface for liquid chromatographs and mass spectrometers. Anal. Chem. 1985, 57, 675–679. [Google Scholar] [CrossRef]
- Douglas, D.J.; Frank, A.J.; Mao, D. Linear ion traps in mass spectrometry. Mass Spectrom. Rev. 2005, 24, 1–29. [Google Scholar] [CrossRef]
- Nolting, D.; Malek, R.; Makarov, A. Ion traps in modern mass spectrometry. Mass Spectrom. Rev. 2019, 38, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Noll, R.J.; Li, H.; Makarov, A.; Hardman, M.; Graham Cooks, R. The Orbitrap: A new mass spectrometer. J. Mass Spectrom. 2005, 40, 430–443. [Google Scholar] [CrossRef] [PubMed]
- Michalski, A.; Damoc, E.; Lange, O.; Denisov, E.; Nolting, D.; Müller, M.; Viner, R.; Schwartz, J.; Remes, P.; Belford, M.; et al. Ultra high resolution linear ion trap Orbitrap mass spectrometer (Orbitrap Elite) facilitates top down LC MS/MS and versatile peptide fragmentation modes. Mol. Cell. Proteom. 2012, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitorino, R. Digging Deep into Peptidomics Applied to Body Fluids. Proteomics 2018, 18, 1700401. [Google Scholar] [CrossRef] [PubMed]
- Maes, E.; Dyer, J.M.; McKerchar, H.J.; Deb-Choudhury, S.; Clerens, S. Protein-protein cross-linking and human health: The challenge of elucidating with mass spectrometry. Expert Rev. Proteom. 2017, 14, 917–929. [Google Scholar] [CrossRef]
- Wang, Y.; Griffiths, W.J.; Jörnvall, H.; Agerberth, B.; Johansson, J. Antibacterial peptides in stimulated human granulocytes: Characterization of ubiquitinated histone H1A. Eur. J. Biochem. 2002, 269, 512–518. [Google Scholar] [CrossRef]
- Theilgaard-Mönch, K.; Jacobsen, L.C.; Nielsen, M.J.; Rasmussen, T.; Udby, L.; Gharib, M.; Arkwright, P.D.; Gombart, A.F.; Calafat, J.; Moestrup, S.K.; et al. Haptoglobin is synthesized during granulocyte differentiation, stored in specific granules, and released by neutrophils in response to activation. Blood 2006, 108, 353–361. [Google Scholar] [CrossRef]
- Weisel, J.W. Fibrinogen and fibrin. Adv. Protein Chem. 2005, 70, 247–299. [Google Scholar]
- Philp, D.; Goldstein, A.L.; Kleinman, H.K. Thymosin beta4 promotes angiogenesis, wound healing, and hair follicle development. Mech. Ageing Dev. 2004, 125, 113–115. [Google Scholar] [CrossRef]
- Reti, R.; Kwon, E.; Qiu, P.; Wheater, M.; Sosne, G. Thymosin beta4 is cytoprotective in human gingival fibroblasts. Eur. J. Oral Sci. 2008, 116, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Liu, M.; Xiong, H. Role of Calprotectin as a Biomarker in Periodontal Disease. Mediat. Inflamm. 2019, 2019, 3515026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terracciano, R.; Pelaia, G.; Preianò, M.; Savino, R. Asthma and COPD proteomics: Current approaches and future directions. Proteom. Clin. Appl. 2015, 9, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Chapple, I.L.; Brock, G.; Eftimiadi, C.; Matthews, J.B. Glutathione in gingival crevicular fluid and its relation to local antioxidant capacity in periodontal health and disease. Mol. Pathol. 2002, 55, 367–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.Y.; Raftery, M.J.; Goyette, J.; Geczy, C.L. S-glutathionylation regulates inflammatory activities of S100A9. J. Biol. Chem. 2010, 285, 14377–14388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, B.A.; Fredericks, L.P. Antimicrobial peptides in the oral environment: Expression and function in health and disease. Curr. Issues Mol. Biol. 2005, 7, 119–133. [Google Scholar] [PubMed]
- Sakamoto, N.; Mukae, H.; Fujii, T.; Ishii, H.; Yoshioka, S.; Kakugawa, T.; Sugiyama, K.; Mizuta, Y.; Kadota, J.; Nakazato, M.; et al. Differential effects of alpha- and beta-defensin on cytokine production by cultured human bronchial epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 288, L508–L513. [Google Scholar] [CrossRef]
- Panyutich, A.V.; Hiemstra, P.S.; van Wetering, S.; Ganz, T. Human neutrophil defensin and serpins form complexes and inactivate each other. Am. J. Respir. Cell Mol. Biol. 1995, 12, 351–357. [Google Scholar] [CrossRef]
- Gursoy, U.K.; Könönen, E. Understanding the roles of gingival beta-defensins. J. Oral Microbiol. 2012, 1, 15127. [Google Scholar] [CrossRef]
- Türkoğlu, O.; Emingil, G.; Kütükçüler, N.; Atilla, G. Gingival crevicular fluid levels of cathelicidin LL-37 and interleukin-18 in patients with chronic periodontitis. J. Periodontol. 2009, 80, 969–976. [Google Scholar] [CrossRef]
- Janciauskiene, S.; Zelvyte, I.; Jansson, L.; Stevens, T. Divergent effects of alpha1-antitrypsin on neutrophil activation, in vitro. Biochem. Biophys. Res. Commun. 2004, 315, 288–296. [Google Scholar] [CrossRef]
- Subramaniyam, D.; Glader, P.; von Wachenfeldt, K.; Burneckiene, J.; Stevens, T.; Janciauskiene, S. C-36 peptide, a degradation product of alpha1-antitrypsin, modulates human monocyte activation through LPS signaling pathways. Int. J. Biochem. Cell Biol. 2006, 38, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Blaurock, N.; Schmerler, D.; Hünniger, K.; Kurzai, O.; Ludewig, K.; Baier, M.; Brunkhorst, F.M.; Imhof, D.; Kiehntopf, M. C-Terminal Alpha-1 Antitrypsin Peptide: A New Sepsis Biomarker with Immunomodulatory Function. Mediat. Inflamm. 2016, 2016, 6129437. [Google Scholar] [CrossRef] [Green Version]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Lamster, I.B.; Ahlo, J.K. Analysis of gingival crevicular fluid as applied to the diagnosis of oral and systemic diseases. Ann. N. Y. Acad. Sci. 2007, 1098, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Preianò, M.; Pasqua, L.; Gallelli, L.; Galasso, O.; Gasparini, G.; Savino, R.; Terracciano, R. Simultaneous extraction and rapid visualization of peptidomic and lipidomic body fluids fingerprints using mesoporous aluminosilicate and MALDI-TOF MS. Proteomics 2012, 12, 3286–3294. [Google Scholar] [CrossRef] [PubMed]
- Albalat, A.; Stalmach, A.; Bitsika, V.; Siwy, J.; Schanstra, J.P.; Petropoulos, A.D.; Vlahou, A.; Jankowski, J.; Persson, F.; Rossing, P.; et al. Improving peptide relative quantification in MALDI-TOF MS for biomarker assessment. Proteomics 2013, 13, 2967–2975. [Google Scholar] [CrossRef]
- Hillenkamp, F.; Peter-Katalinic, J. MALDI MS: A Practical Guide to Instrumentation, Methods and Applications, 2nd ed.; Whiley-VCH Verlag GmbH & Co KGaA: Weinheim, Germany, 2007; pp. 393–394. [Google Scholar]
- Szájli, E.; Fehér, T.; Medzihradszky, K.F. Investigating the Quantitative Nature of MALDI-TOF MS. Mol. Cell. Proteom. 2008, 7, 2410–2418. [Google Scholar] [CrossRef] [Green Version]
- Bucknall, M.; Fung, K.Y.; Duncan, M. Practical quantitative biomedical applications of MALDI-TOF mass spectrometry. J. Am. Soc. Mass Spectrom. 2002, 13, 1015–1027. [Google Scholar] [CrossRef] [Green Version]
- Urso, E.; Le Pera, M.; Bossio, S.; Sprovieri, T.; Qualtieri, A. Quantification of thymosin beta(4) in human cerebrospinal fluid using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Anal. Biochem. 2010, 402, 13–19. [Google Scholar] [CrossRef]
- Xu, H.; Liu, M.; Huang, X.; Min, Q.; Zhu, J.J. Multiplexed Quantitative MALDI MS Approach for Assessing Activity and Inhibition of Protein Kinases Based on Postenrichment Dephosphorylation of Phosphopeptides by Metal-Organic Framework-Templated Porous CeO2. Anal. Chem. 2018, 90, 9859–9867. [Google Scholar] [CrossRef]
- Guitot, K.; Scarabelli, S.; Drujon, T.; Bolbach, G.; Amoura, M.; Burlina, F.; Jeltsch, A.; Sagan, S.; Guianvarc’h, D. Label-free measurement of histone lysine methyltransferases activity by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Anal. Biochem. 2014, 456, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Tucher, J.; Somasundaram, P.; Tholey, A. Quantitative MALDI MS Using Ionic Liquid Matrices in Advances. In MALDI and Laser-Induced Soft Ionization Mass Spectrometry; Cramer, R., Ed.; Springer International Publishing: Switzerland, Germany, 2016; pp. 77–90. [Google Scholar]
- Deng, H.; Xu, Y.; Zhang, L. Proteomic analysis in drug discovery. In Handbook of Drug Screening, 2nd ed.; Sethala, R., Zhang, L., Eds.; Informa Healthcare: New York, NY, USA, 2009; pp. 117–138. [Google Scholar]
- Terracciano, R.; Preianò, M.; Maggisano, G.; Pelaia, C.; Savino, R. Hexagonal Mesoporous Silica as a Rapid, Efficient and Versatile Tool for MALDI-TOF MS Sample Preparation in Clinical Peptidomics Analysis: A Pilot Study. Molecules 2019, 24, 2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteom. 2012, 11, M111–M010587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G. PEAKS: Powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef] [PubMed]
- Budamgunta, H.; Olexiouk, V.; Luyten, W.; Schildermans, K.; Maes, E.; Boonen, K.; Menschaert, G.; Baggerman, G. Comprehensive Peptide Analysis of Mouse Brain Striatum Identifies Novel sORF-Encoded Polypeptides. Proteomics 2018, 18, e1700218. [Google Scholar] [CrossRef]
- Taylor, J.J.; Preshaw, P.M. Gingival crevicular fluid and saliva. Periodontology 2000 2016, 70, 7–10. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Ref. | Collection Devices/Elution Buffer/ Pre-Processing | Storage Conditions | Protein Quantitation | MS Normalization | MS-Approach | Naturally Occurring Peptides &Small Proteins Detected: Main Results |
|---|---|---|---|---|---|---|
| Lundy et al. [16] | Paper strips/sodium phosphate containing NaCl; NO PIC; RP-HPLC. | −70 °C, time not specified. | Yes | Not required (a) | N-terminal aminoacid sequencing and LC-MS | S100-A8 |
| Diamond et al. [18] | Periopaper strips/5% acetic acid; NO PIC; ProteinChip. | Not specified. | No | No | SELDI-TOF | α-defensins 1–3, hBD-1 derived peptides, hBD-2 |
| Dommisch et al. [17] | Periopaper strips/5% acetic acid; NO PIC; ProteinChip. | Not specified. | Yes (b) | No | SELDI-TOF | α-defensins 1–3, Cathelicidin antimicrobial peptide LL-37 |
| Lundy et al. [19] | Periopaper strips/sodium phosphate containing NaCl; NO PIC; Unfractionated. | −80 °C, time not specified. | Yes | No | MALDI-TOF | α-defensins 1–3 |
| Wen et al. [20] | Paper points/2.5% TFA; NO PIC; Unfractionated. | −80 °C, time not specified. | No | Yes | MALDI-TOF | m/z = 1660.2, 1783.0, 2912.5, 4178.6, 5064.9, 6108.9 |
| Ngo et al. [6] | Glass microcapillary tubes/water or 0.1% TFA for HPLC fractionation; NO PIC; ZipTips or RP-HPLC. | −70 °C, time not specified. | No | No | MALDI-TOF/TOF | α-defensins 1–3, Peptides fragments from: Peptide salivary low MW, Proline-rich phosphoprotein, Beta globin, Peptide PA saliva, (Peptide PB saliva, Peptide PC saliva, H2A histone family, Ig light chain variable region, Glyceraldehyde-3-phosphate, Fibroblast growth factor, Albumin, Collagen, Ig heavy chain, Thioredoxin, Statherin, Hemoglobin alpha 1, Albumin Dbox binding protein, Collagen R-1 type III, Metallopeptidase |
| Ngo et al. [21] | Glass microcapillary tubes/GCF (0.2–1.5 µL) was dispensed with 2.5% TFA; NO PIC; ZipTips. | −70 °C, time not specified. | No | Yes | MALDI-TOF | Attachment loss sites vs. stable sites (m/z): 2023.50, 4042.80, 4490.30, 4525.30, 5232.30, 5502.20, 6890.10, 10939.00, 12833.00, 14008.00 |
| Preianò et al. [22] | Paper points, Paper strips/2.5% TFA or 5% acetic acid; PIC and NO PIC; Unfractionated. | GCF was immediately analyzed by MS. | Yes | Yes | MALDI-TOF/TOF | α-defensins 1–4, Thymosin β4 |
| Preianò et al. [23] | Paper strips/2.5%TFA; PIC and NO PIC; Unfractionated. | −20 °C/1 or 3 months, −80 °C/1 or 3 months and immediately analyzed | Yes | Yes | MALDI-TOF/TOF | α-defensins 1–4, hBD-2, Thymosin β4, S100-A8 and its oxidized forms, S100-A9 and its isoforms, Lysozyme C |
| Preianò et al. [24] | Paper strips/2.5%TFA; PIC and NO PIC, Unfractionated. | −20 °C/1 month, −80 °C/1 month and immediately analyzed. | Yes | Yes | MALDI-TOF/TOF | α-defensins 1–3, hBD-2, C-36 peptide of AAT, Thymosin β4, Thymosin β10, S100-A8 and its oxidized forms, S100-A9 and its isoforms, Lysozyme C |
| Tang et al. [25] | Paper strips/phosphate-buffered saline; NO PIC; Weak cation exchange magnetic beads. | −80 °C, time not specified (c). | No | Yes | MALDI-TOF | Haptoglobin derived peptide and other unidentified peptides: m/z = 4126.6, 5407.7, 5416.0 |
| Antezack et al. [26] | Paper points/HPLC-grade water; NO PIC; Unfractionated. | +4 °C and analyzed within 24 to 48 h. | No | Yes | MALDI-TOF | m/z = 3775, 4235, 4944, 5296, 5728, 5893, 10586, 11324, 11,359 and 11447 |
| Pisano et al. [27] | Paper cones/0.2% TFA aqueous solution; NO PIC; RP-HPLC. | −80 °C, time not specified. | No | Yes | ESI-Ion Trap | α-defensins 1–4, Statherin, Peptide PB, Cystatin A |
| Inzitari et al. [28] | Paper cones/0.2% TFA aqueous solution; NO PIC; RP-HPLC. | Storage conditions not specified. | Yes | Yes | ESI-Ion Trap | Thymosin β4, Thymosin β4 sulfoxide, Thymosin β10 |
| Dassatti et al. [29] | Paper cones/Buffer not specified; Use of PIC not specified; RP-HPLC. | −80 °C, time not specified. | No | Yes | ESI-Ion Trap/ Orbitrap | α-defensins 1–4, Thymosin β4, Thymosin β4 fragment (21–44), Thymosin β10, Fibrinopeptide A, Fibrinopeptide A fragments (21–35) and (22–35), Fibrinopeptide B |
| Naturally Occurring Peptides | Methods | Study Groups and Number of Subjects (n) | Study Groups and Protein Expression Level | Ref. |
|---|---|---|---|---|
| C-36 AAT | MALDI-TOF | H (10) vs. G (10) | ↓H/↑G * | Preianò et al. [24] |
| Cathelicidin antimicrobial peptide LL-37 | SELDI-TOF | H sites (n = 8) and G sites (n = 8) in subjects with good general health (4). | ↓H sites/↑G sites in H * | Dommisch et al. [17] |
| Fibrinopeptide A | ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] |
| Fibrinopeptide A fragment (21–35) | ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] |
| Fibrinopeptide A fragment (22–35) | ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] |
| Fibrinopeptide B | ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] |
| Haptoglobin derived peptide | MALDI-TOF | CP (17) and H (16). | ↓H/↑CP * | Tang et al. [25] |
| hBD-1 | SELDI-TOF | Patients with mild to moderate gingival inflammation (2). | ↑mild gingival inflammation/↓moderate gingival inflammation | Diamond et al. [18] |
| hBD-2 | SELDI-TOF | Patients with mild to moderate gingival inflammation (2). | ↓mild gingival inflammation/↑moderate gingival inflammation | Diamond et al. [18] |
| HNP-1 | SELDI-TOF | Patients with mild to moderate gingival inflammation (2). | ↓mild gingival inflammation/↑moderate gingival inflammation | Diamond et al. [18] |
| SELDI-TOF | H sites (n = 8) and G sites (n = 8) in subjects with good general health (4). | ↓H sites/↑G sites in H | Dommisch et al. [17] | |
| MALDI-TOF | P(11) and H (12) | ↑H/↓P | Lundy et al. [19] | |
| ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] | |
| HNP-2 | SELDI-TOF | Patients with mild to moderate gingival inflammation (2) | ↓mild gingival inflammation/↑moderate gingival inflammation | Diamond et al. [18] |
| SELDI-TOF | H sites (n = 8) and G sites (n = 8) in subjects with good general health (4) | ↓H sites/↑G sites in H | Dommisch et al. [17] | |
| MALDI-TOF | P (11) and H (12) | ↑H/↓P | Lundy et al. [19] | |
| MALDI-TOF | H (10) vs. G (10) | ↓H/↑G * | Preianò et al. [24] | |
| ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] | |
| HNP-3 | SELDI-TOF | Patients with mild to moderate gingival inflammation (2). | ↓mild gingival inflammation/↑moderate gingival inflammation | Diamond et al. [18] |
| MALDI-TOF | P (11) and H (12) | ↑H/↓P | Lundy et al. [19] | |
| ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] | |
| HNP-4 | ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites ↓H/↑G sites | Dassatti et al. [29] |
| S100-A8 | N-terminal amino acid sequencing and LC-MS | H, G and P sites in patients with P (15) and H sites in H subjects (5). | ↓H/↑G sites in P * ↓H/↑P sites in P * ↓H sites in P/↑G sites in P * ↓H sites in P/↑P sites in P * | Lundy et al. [16] |
| S100-A9 | MALDI-TOF | H (10) vs. G (10) | ↑H/↓G * | Preianò et al. [24] |
| S100-A9 Glutathionylated | MALDI-TOF | H (10) vs. G (10) | ↓H/↑G * | Preianò et al. [24] |
| Thymosin β-4 | MALDI-TOF | H (10) vs. G (10) | ↑H/↓G * | Preianò et al. [24] |
| ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↑H/↓P sites * ↑H/↓G sites * | Dassatti et al. [29] | |
| Thymosin β-4 fragment (21–44) | ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↓H/↑P sites * ↓H/↑G sites * | Dassatti et al. [29] |
| Thymosin β-10 | ESI-Ion Trap/Orbitrap | G and P sites in women after pregnancy (10) and not pregnant women as H controls (10). | ↑H/↓P sites * ↑H/↓G sites * | Dassatti et al. [29] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preianò, M.; Savino, R.; Villella, C.; Pelaia, C.; Terracciano, R. Gingival Crevicular Fluid Peptidome Profiling in Healthy and in Periodontal Diseases. Int. J. Mol. Sci. 2020, 21, 5270. https://doi.org/10.3390/ijms21155270
Preianò M, Savino R, Villella C, Pelaia C, Terracciano R. Gingival Crevicular Fluid Peptidome Profiling in Healthy and in Periodontal Diseases. International Journal of Molecular Sciences. 2020; 21(15):5270. https://doi.org/10.3390/ijms21155270
Chicago/Turabian StylePreianò, Mariaimmacolata, Rocco Savino, Chiara Villella, Corrado Pelaia, and Rosa Terracciano. 2020. "Gingival Crevicular Fluid Peptidome Profiling in Healthy and in Periodontal Diseases" International Journal of Molecular Sciences 21, no. 15: 5270. https://doi.org/10.3390/ijms21155270
APA StylePreianò, M., Savino, R., Villella, C., Pelaia, C., & Terracciano, R. (2020). Gingival Crevicular Fluid Peptidome Profiling in Healthy and in Periodontal Diseases. International Journal of Molecular Sciences, 21(15), 5270. https://doi.org/10.3390/ijms21155270