Network Neuromodulation of Opioid and GABAergic Receptors Following a Combination of “Juvenile” and “Adult Stress” in Rats



Abstract
:1. Introduction
2. Results
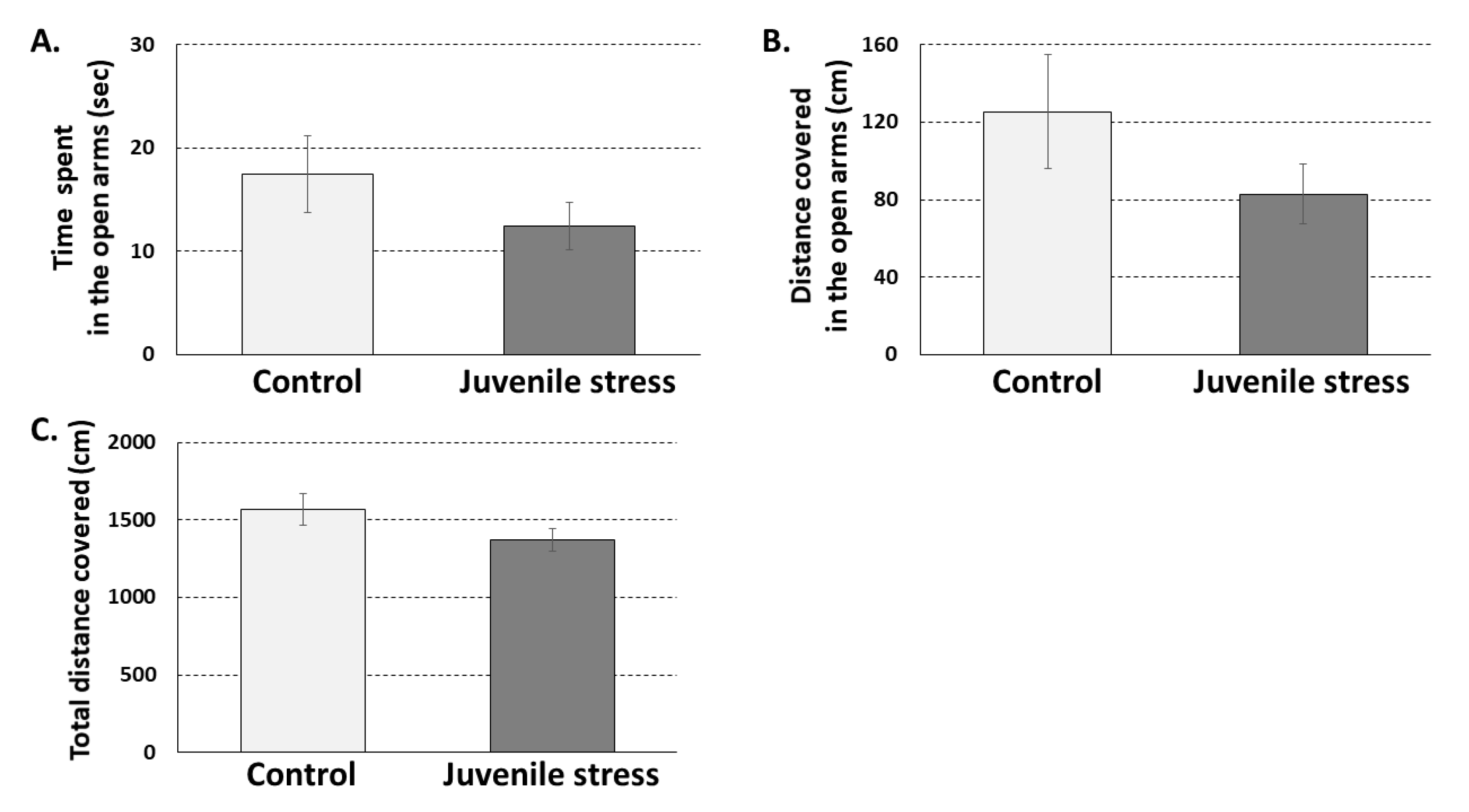
2.1. The Exposure to Juvenile Stress by Itself Has Limited Effect on Behavior in Adulthood
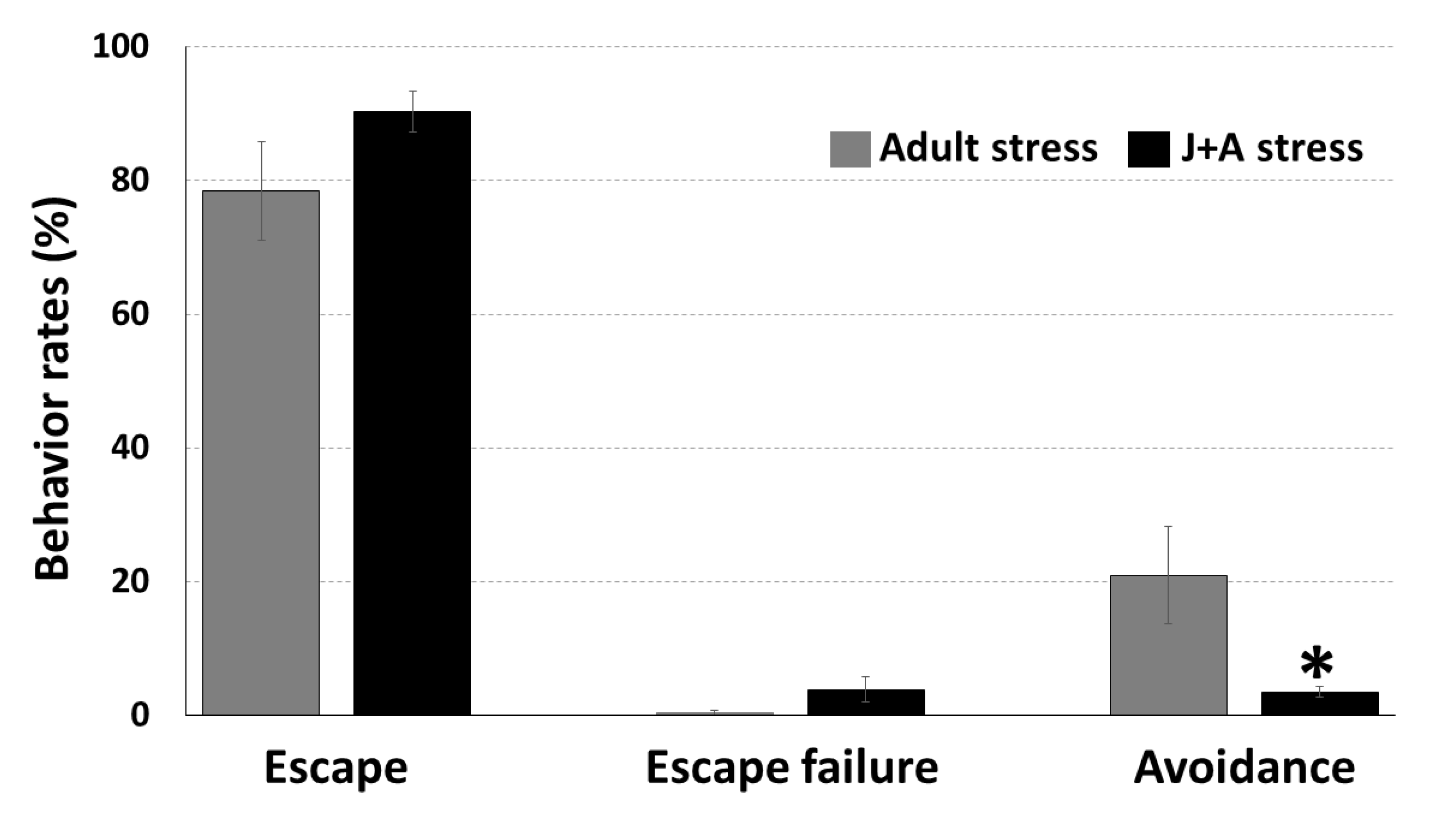
2.2. The Exposure to Juvenile Stress Affected Performance in the Two-Way Shuttle Avoidance Task in Adulthood
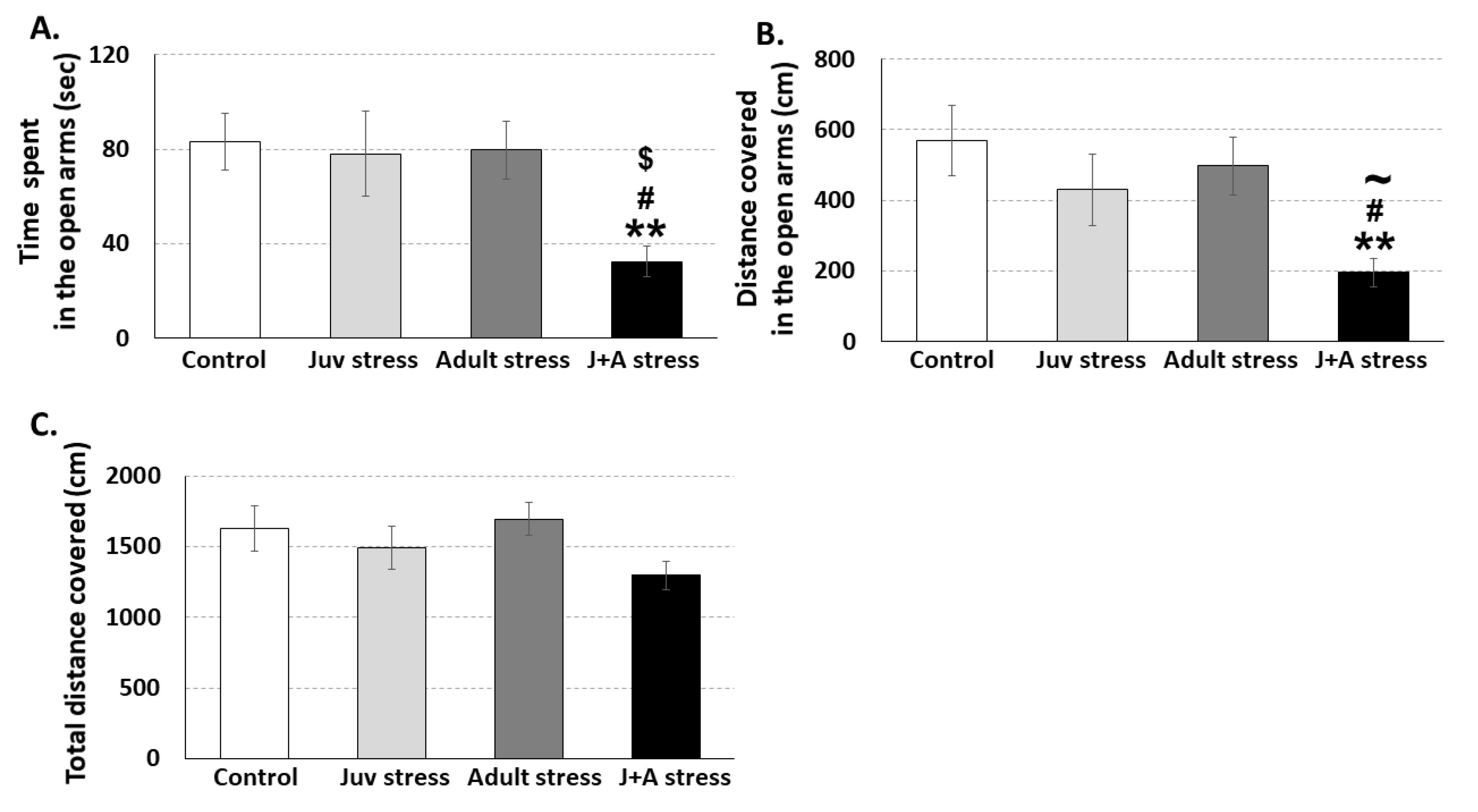
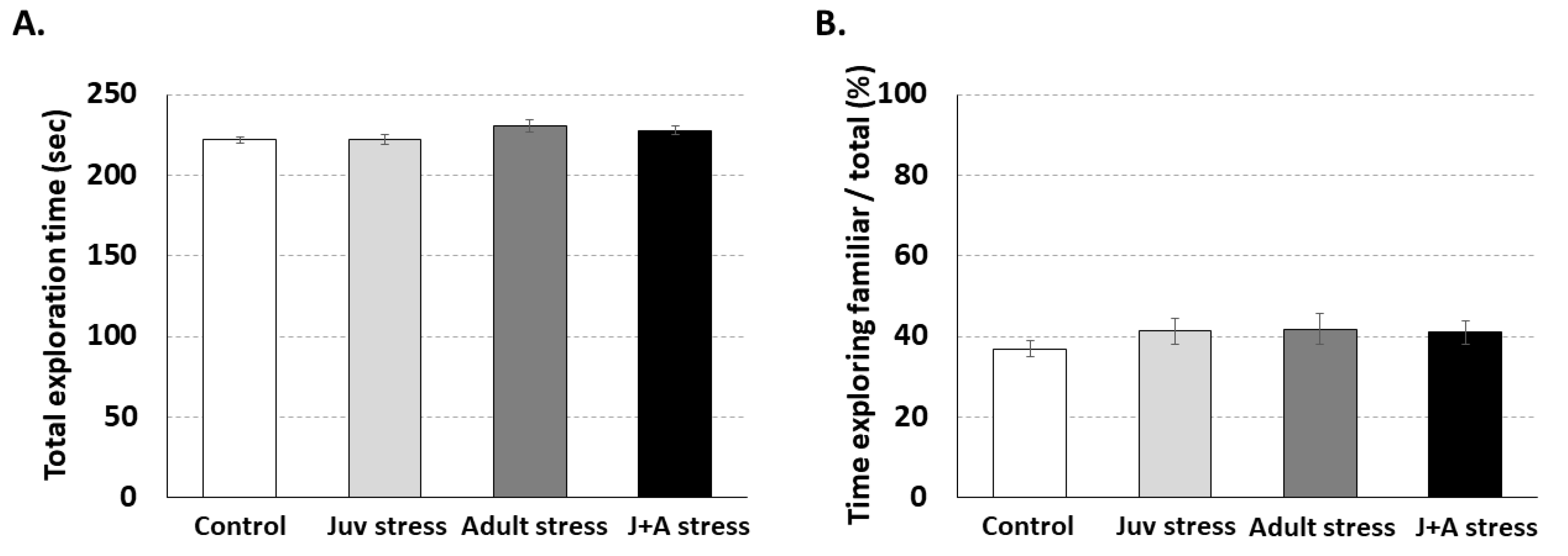
2.3. The Effects of Combined Juvenile Stress and Stress in Adulthood
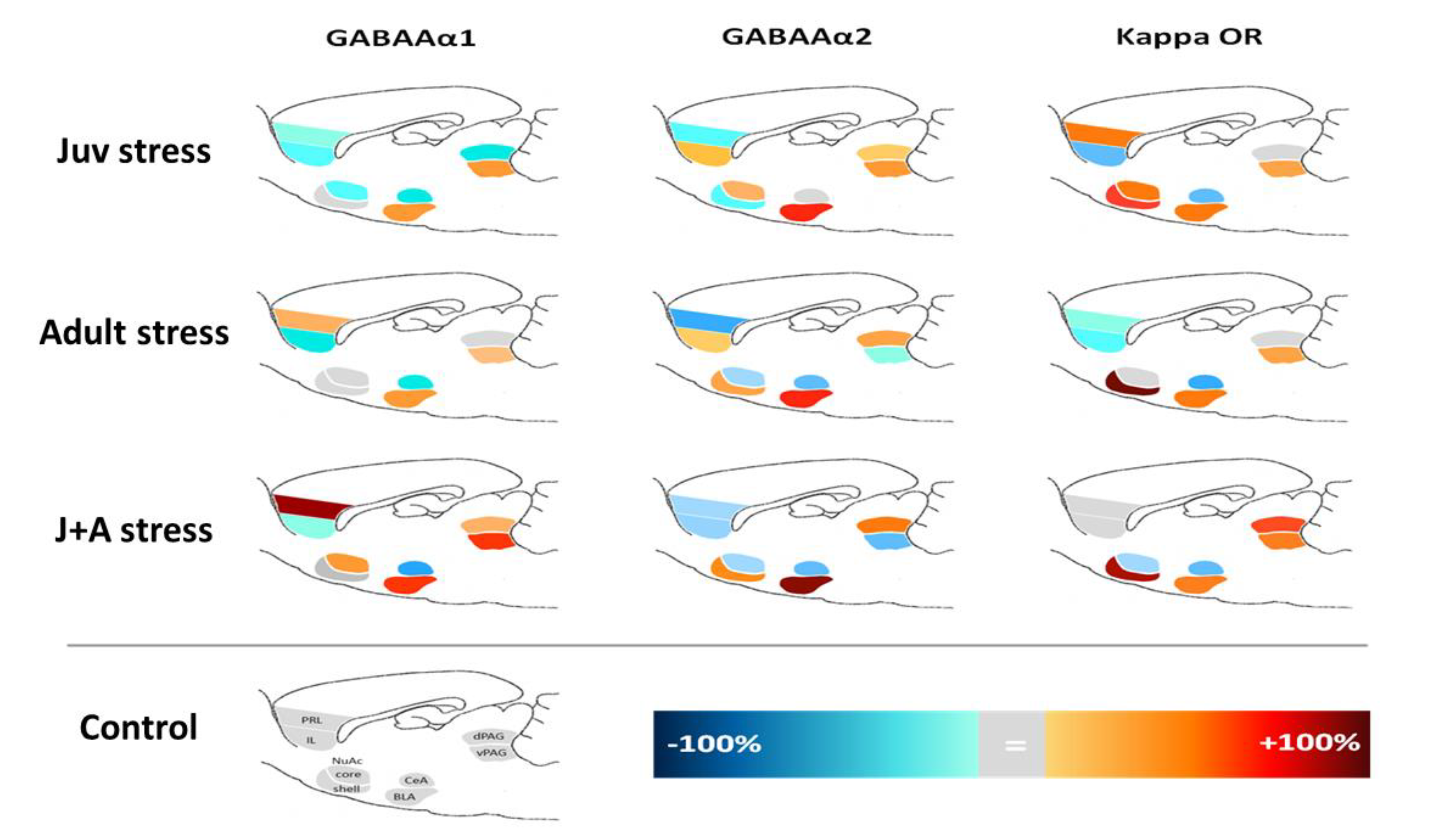
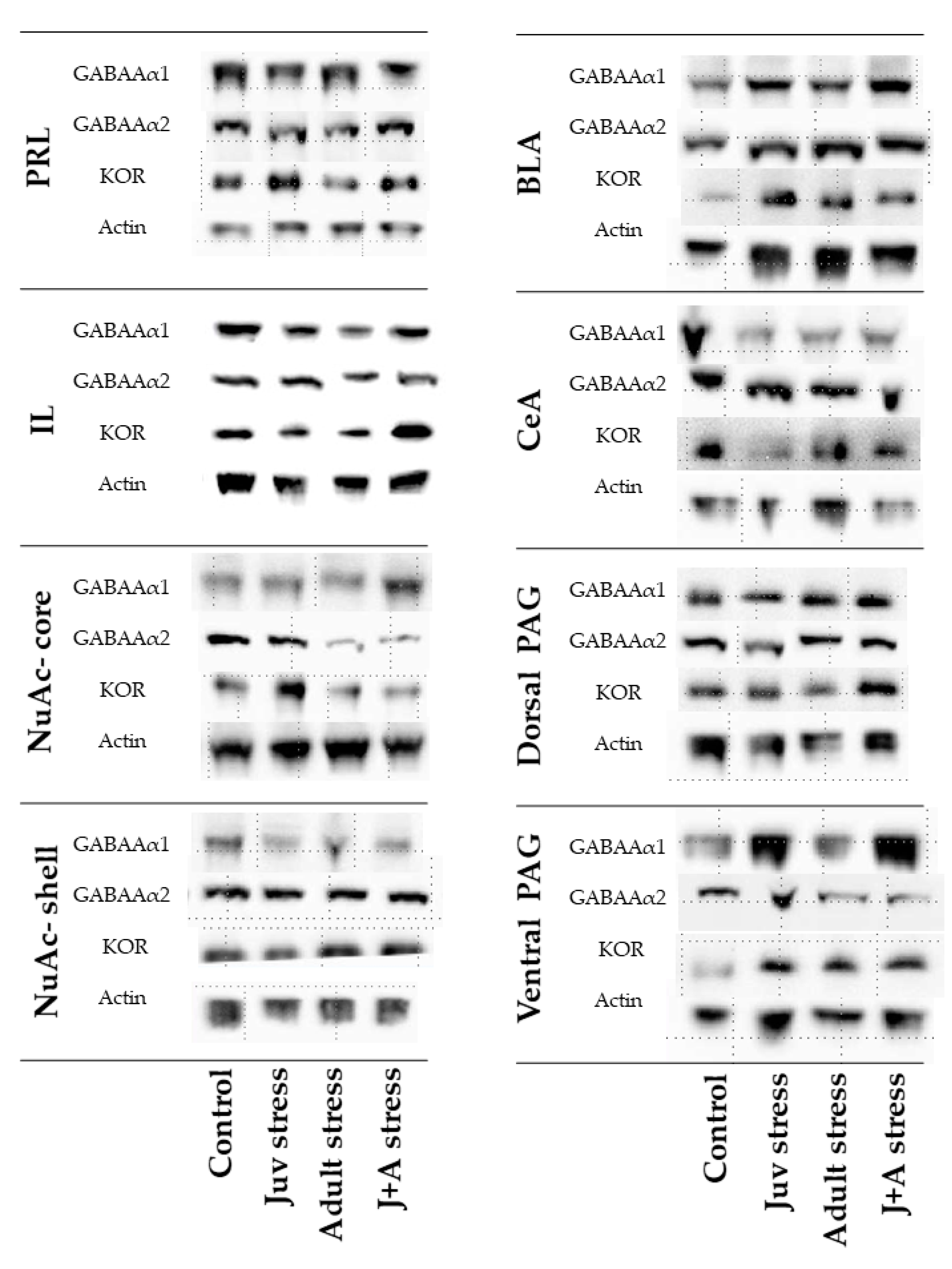
2.4. Expression of GABAAR and KOR
3. Discussion
4. Materials and Methods
4.1. Animals
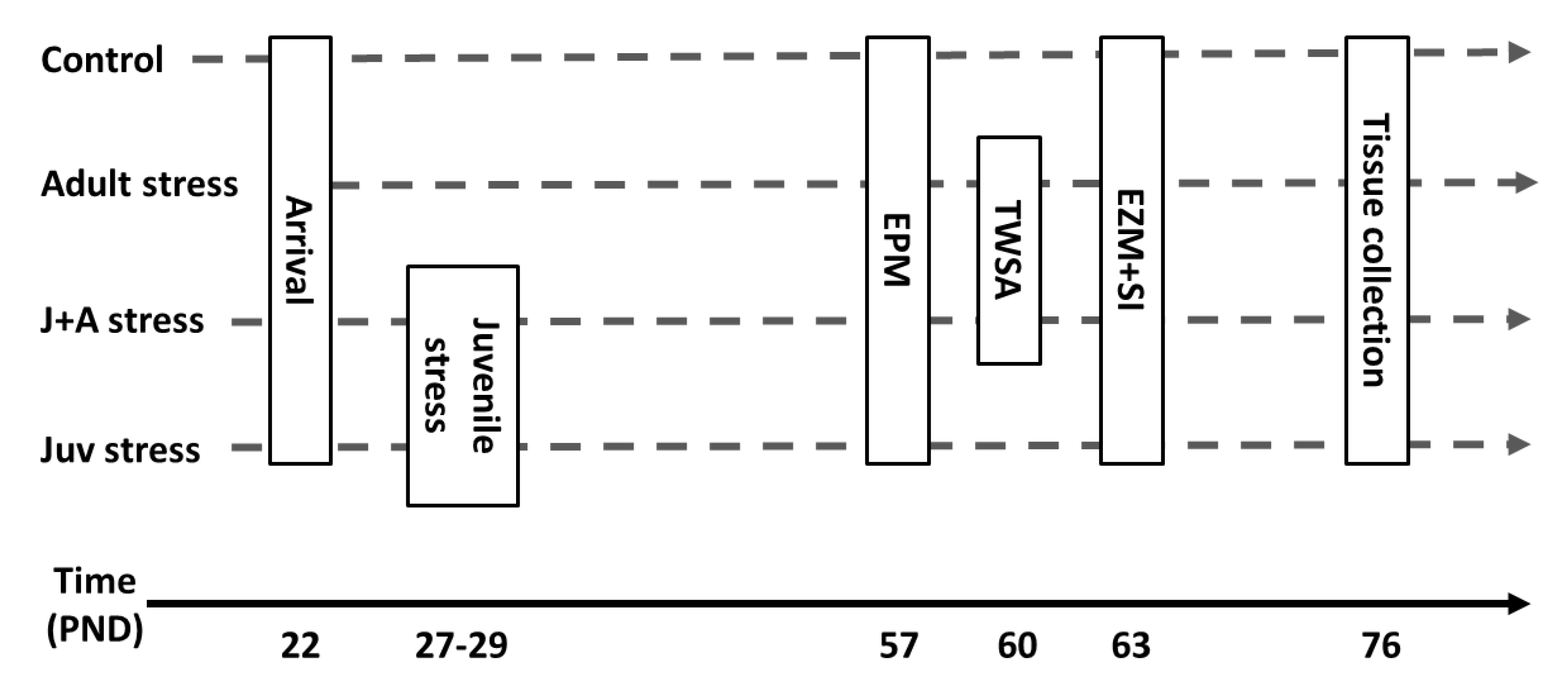
4.2. Study Design
- (1)
- Juvenile stress group (“juv stress”): animals exposed to “juvenile stress” at juvenility (27–29 PND) and to the EPM, EZM, and SI tests in adulthood (57 + PND).
- (2)
- Juvenile stress + adult stress group (“J + A stress”): animals exposed to “juvenile stress” at juvenility (27–29 PND); to the TWSA as “adult stress”; and to the EPM, EZM, and SI tests in adulthood (57 + PND).
- (3)
- Adult stress (adult stress): animals exposed to the TWSA as “adult stress” and to the EPM, EZM, and SI tests in adulthood (57 PND).
- (4)
- Control group (control): animals not exposed to any stress procedure but only to the EPM, EZM, and SI tests in adulthood (57 PND).
4.3. Behavioral Procedures
4.3.1. “Juvenile Stress”
4.3.2. Elevated Plus Maze Test
4.3.3. “Adult Stress”—Two Way Shuttle Avoidance
4.3.4. Elevated Zero Maze Test
4.3.5. Social Interaction Test
4.4. Brain Tissue Harvesting
4.5. Biochemical Methods
4.5.1. Homogenization
4.5.2. Western Blot Analysis
Reagents
Quantification
4.6. Statistics
4.7. Ethical Approval
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PTSD | Posttraumatic stress disorder |
| GABA | Directory of open access journals |
| mPFC | Medial prefrontal cortex |
| NAc | Nucleus accumbens |
| PAG | Periaqueductal gray |
| BLA | Basolateral amygdala |
| TWSA | Two-way shuttle avoidance task |
| EZM | Elevated zero maze |
| EPM | Elevated plus maze |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region and Receptor Type | Control | Juv Stress | Adult Stress | J+A Stress | F Value | Sig. |
|---|---|---|---|---|---|---|
| PRL | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 10.7 100 ± 10.5 100 ± 8.3 | 87 ± 7.8 81 ± 7.9 145 ± 29.5 | 131 ± 30.0 60 ± 8.1 89 ± 11.0 | 219 ± 67.4 83 ± 14.5 108 ± 14.9 | 2.47 2.93 2.08 | 0.072 0.042 0.115 |
| IL | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 9.4 100 ± 6.9 100 ± 11.1 | 80 ± 8.8 117 ± 17.8 71 ± 16.7 | 76 ± 15.8 108 ± 18.8 76 ± 25.4 | 88 ± 17.9 76 ± 13.0 95 ± 31.9 | 0.45 1.41 0.21 | 0.717 0.251 0.888 |
| NuAc-core | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 7.3 100 ± 6.6 100 ± 9.8 | 79 ± 8.1 128 ± 18.7 146 ± 32.6 | 104 ± 16.4 82 ± 17.7 95 ± 12.4 | 139 ± 21.8 80 ± 12.6 86 ± 11.0 | 3.23 2.40 1.97 | 0.030 0.078 0.130 |
| NuAc-shell | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 11.4 100 ± 13.2 100 ± 25.2 | 94 ± 14.3 79 ± 8.1 159 ± 42.9 | 100 ± 27.5 122 ± 26.5 285 ± 105 | 100 ± 12.9 120 ± 21.2 200 ± 29.4 | 0.04 1.14 1.32 | 0.990 0.343 0.279 |
| BLA | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 6.8 100 ± 9.1 100 ± 9.7 | 138 ± 14.2 181 ± 40.0 134 ± 13.1 | 143 ± 22.6 186 ± 29.2 148 ± 15.5 | 156 ± 27.8 195 ± 21.9 139 ± 22.7 | 2.05 2.45 1.72 | 0.119 0.074 0.174 |
| CeA | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 8.9 100 ± 9.2 100 ± 8.0 | 76 ± 12.4 93 ± 12.2 61 ± 7.8 | 75 ± 8.4 71 ± 16.4 56 ± 8.4 | 54 ± 10.4 70 ± 13.6 59 ± 12.9 | 1.82 1.35 2.98 | 0.156 0.268 0.040 |
| Dorsal PAG | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 10.1 100 ± 7.4 100 ± 21.2 | 75 ± 8.5 112 ± 8.6 97 ± 10.2 | 106 ± 17.7 133 ± 20.1 101 ± 14.6 | 131 ± 36.4 147 ± 43.1 168 ± 32.5 | 1.13 0.56 2.32 | 0.346 0.640 0.086 |
| Ventral PAG | ||||||
| GABAAα1 GABAAα2 KOR | 100 ± 11.9 100 ± 11.4 100 ± 10.4 | 140 ± 14.9 135 ± 16.2 132 ± 10.0 | 121 ± 19.6 90 ± 10.4 134 ± 14.2 | 165 ± 39.7 69 ± 9.2 *# 149 ± 24.7 | 1.34 5.35 1.32 | 0.270 0.003 0.279 |

Appendix B
| Classification by GABAAR α1. | |||||
| Observed | Predicted | ||||
| Control | Juvenile | Adult | Juvenile + Adult | % Correct | |
| Control | 11 | 1 | 1 | 0 | 84.6% |
| Juv stress | 1 | 10 | 1 | 2 | 71.4% |
| Adult stress | 3 | 3 | 6 | 1 | 46.2% |
| J+A stress | 0 | 1 | 3 | 10 | 71.4% |
| Overall % | 27.8% | 27.8% | 20.4% | 24.1% | 68.5% |
| χ2(24) = 55.5, p < 0.001 | |||||
| Classification by GABAAR α2. | |||||
| Observed | Predicted | ||||
| Control | Juvenile | Adult | Juvenile + Adult | % Correct | |
| Control | 10 | 2 | 0 | 1 | 76.9% |
| Juv stress | 3 | 11 | 0 | 0 | 78.6% |
| Adult stress | 0 | 1 | 9 | 3 | 69.2% |
| J+A stress | 1 | 1 | 3 | 9 | 64.3% |
| Overall % | 25.9% | 27.8% | 22.2% | 24.1% | 72.2% |
| χ2(24) = 77.1, p < 0.001 | |||||
| Classification by KOR. | |||||
| Observed | Predicted | ||||
| Control | Juvenile | Adult | Juvenile + Adult | % Correct | |
| Control | 10 | 0 | 1 | 2 | 76.9% |
| Juv stress | 1 | 6 | 3 | 1 | 54.5% |
| Adult stress | 1 | 2 | 7 | 3 | 53.8% |
| J+A stress | 2 | 2 | 1 | 9 | 64.3% |
| Overall % | 27.5% | 19.6% | 23.5% | 29.4% | 62.7% |
| χ2(24) = 55.3, p < 0.001 | |||||
| Brain Region | GABAAR α1 Sig. | GABAAR α2 Sig. | KOR Sig. |
|---|---|---|---|
| PRL | 0.130 | 0.006 | 0.051 |
| IL | 0.051 | 0.051 | 0.011 |
| NuAc core | 0.025 | 0.029 | 0.254 |
| NuAc shell | 0.770 | 0.000 | 0.029 |
| CeA | 0.244 | 0.029 | 0.001 |
| BLA | 0.146 | 0.000 | 0.013 |
| dPA | 0.006 | 0.477 | 0.044 |
| vPAG | 0.013 | 0.000 | 0.105 |
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 978-0-89042-555-8. [Google Scholar]
- Kessler, R.C. Posttraumatic Stress Disorder in the National Comorbidity Survey. Arch. Gen. Psychiatry 1995, 52, 1048. [Google Scholar] [CrossRef] [PubMed]
- DiGangi, J.A.; Gomez, D.; Mendoza, L.; Jason, L.A.; Keys, C.B.; Koenen, K.C. Pretrauma risk factors for posttraumatic stress disorder: A systematic review of the literature. Clin. Psychol. Rev. 2013, 33, 728–744. [Google Scholar] [CrossRef] [PubMed]
- Nemeroff, C.B. Neurobiological consequences of childhood trauma. J. Clin. Psychiatry 2004, 65 (Suppl. S1), 18–28. [Google Scholar] [PubMed]
- Agorastos, A.; Pervanidou, P.; Chrousos, G.P.; Baker, D.G. Developmental Trajectories of Early Life Stress and Trauma: A Narrative Review on Neurobiological Aspects Beyond Stress System Dysregulation. Front. Psychiatry 2019, 10, 118. [Google Scholar] [CrossRef] [Green Version]
- Cougle, J.R.; Timpano, K.R.; Sachs-Ericsson, N.; Keough, M.E.; Riccardi, C.J. Examining the unique relationships between anxiety disorders and childhood physical and sexual abuse in the National Comorbidity Survey-Replication. Psychiatry Res. 2010, 177, 150–155. [Google Scholar] [CrossRef]
- Pratchett, L.C.; Yehuda, R. Foundations of posttraumatic stress disorder: Does early life trauma lead to adult posttraumatic stress disorder? Dev. Psychopathol. 2011, 23, 477–491. [Google Scholar] [CrossRef]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef]
- Tsoory, M.; Richter-Levin, G. Learning under stress in the adult rat is differentially affected by ‘juvenile’ or ‘adolescent’ stress. Int. J. Neuropsychopharmacol. 2006, 9, 713. [Google Scholar] [CrossRef] [Green Version]
- Avital, A.; Richter-Levin, G. Exposure to juvenile stress exacerbates the behavioural consequences of exposure to stress in the adult rat. Int. J. Neuropsychopharmacol. 2005, 8, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Horovitz, O.; Tsoory, M.M.; Hall, J.; Jacobson-Pick, S.; Richter-Levin, G. Post-Weaning to Pre-Pubertal (‘Juvenile’) Stress: A Model of Induced Predisposition to Stress-Related Disorders. Neuroendocrinology 2012, 95, 56–64. [Google Scholar] [CrossRef]
- Chen, F.; Ke, J.; Qi, R.; Xu, Q.; Zhong, Y.; Liu, T.; Li, J.; Zhang, L.; Lu, G. Increased Inhibition of the Amygdala by the mPFC may Reflect a Resilience Factor in Post-traumatic Stress Disorder: A Resting-State fMRI Granger Causality Analysis. Front. Psychiatry 2018, 9, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberzon, I.; Sripada, C.S. The functional neuroanatomy of PTSD: A critical review. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2007; Volume 167, pp. 151–169. ISBN 978-0-444-53140-7. [Google Scholar]
- Francati, V.; Vermetten, E.; Bremner, J.D. Functional neuroimaging studies in posttraumatic stress disorder: Review of current methods and findings. Depress. Anxiety 2007, 24, 202–218. [Google Scholar] [CrossRef]
- Maeng, L.Y.; Milad, M.R. Post-Traumatic Stress Disorder: The Relationship Between the Fear Response and Chronic Stress. Chronic Stress 2017, 1, 247054701771329. [Google Scholar] [CrossRef] [PubMed]
- Rau, V.; DeCola, J.P.; Fanselow, M.S. Stress-induced enhancement of fear learning: An animal model of posttraumatic stress disorder. Neurosci. Biobehav. Rev. 2005, 29, 1207–1223. [Google Scholar] [CrossRef] [PubMed]
- Knapska, E.; Radwanska, K.; Werka, T.; Kaczmarek, L. Functional Internal Complexity of Amygdala: Focus on Gene Activity Mapping After Behavioral Training and Drugs of Abuse. Physiol. Rev. 2007, 87, 1113–1173. [Google Scholar] [CrossRef]
- Knapska, E.; Maren, S. Reciprocal patterns of c-Fos expression in the medial prefrontal cortex and amygdala after extinction and renewal of conditioned fear. Learn. Mem. 2009, 16, 486–493. [Google Scholar] [CrossRef] [Green Version]
- Sousa, N. The dynamics of the stress neuromatrix. Mol. Psychiatry 2016, 21, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Joëls, M.; Baram, T.Z. The neuro-symphony of stress. Nat. Rev. Neurosci. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Dedovic, K.; Duchesne, A.; Andrews, J.; Engert, V.; Pruessner, J.C. The brain and the stress axis: The neural correlates of cortisol regulation in response to stress. NeuroImage 2009, 47, 864–871. [Google Scholar] [CrossRef]
- Benarroch, E.E. Periaqueductal gray: An interface for behavioral control. Neurology 2012, 78, 210–217. [Google Scholar] [CrossRef]
- Brandão, M.L.; Lovick, T.A. Role of the dorsal periaqueductal gray in posttraumatic stress disorder: Mediation by dopamine and neurokinin. Transl. Psychiatry 2019, 9, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.J.; Horovitz, O.; Pellman, B.A.; Tan, L.M.; Li, Q.; Richter-Levin, G.; Kim, J.J. Dorsal periaqueductal gray-amygdala pathway conveys both innate and learned fear responses in rats. Proc. Natl. Acad. Sci. USA 2013, 110, 14795–14800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, S.; McNally, G.P. Complementary roles for amygdala and periaqueductal gray in temporal-difference fear learning. Learn. Mem. 2008, 16, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, S.J.; Nestler, E.J. The brain reward circuitry in mood disorders. Nat. Rev. Neurosci. 2013, 14, 609–625. [Google Scholar] [CrossRef] [Green Version]
- Noh, S.J.; Kang, D.-W.; Yoo, S.B.; Lee, J.Y.; Kim, J.Y.; Kim, B.-T.; Lee, J.-H.; Jahng, J.W. Stress-responsive hypothalamic-nucleus accumbens regulation may vary depending on stressors. Indian J. Exp. Biol. 2012, 50, 447–454. [Google Scholar]
- Bossert, J.M.; Stern, A.L.; Theberge, F.R.M.; Marchant, N.J.; Wang, H.-L.; Morales, M.; Shaham, Y. Role of Projections from Ventral Medial Prefrontal Cortex to Nucleus Accumbens Shell in Context-Induced Reinstatement of Heroin Seeking. J. Neurosci. 2012, 32, 4982–4991. [Google Scholar] [CrossRef]
- Brog, J.S.; Salyapongse, A.; Deutch, A.Y.; Zahm, D.S. The patterns of afferent innervation of the core and shell in the ?Accumbens? part of the rat ventral striatum: Immunohistochemical detection of retrogradely transported fluoro-gold. J. Comp. Neurol. 1993, 338, 255–278. [Google Scholar] [CrossRef]
- Mcdonald, A.J. Organization of amygdaloid projections to the prefrontal cortex and associated striatum in the rat. Neuroscience 1991, 44, 1–14. [Google Scholar] [CrossRef]
- Thomas, K.L.; Hall, J.; Everitt, B.J. Cellular imaging with zif268 expression in the rat nucleus accumbens and frontal cortex further dissociates the neural pathways activated following the retrieval of contextual and cued fear memory: Fear CS-induced zif268 expression. Eur. J. Neurosci. 2002, 16, 1789–1796. [Google Scholar] [CrossRef]
- Lanius, R.A.; Brand, B.; Vermetten, E.; Frewen, P.A.; Spiegel, D. The Dissociative Subtype of Posttraumatic Stress Disorder: Rationale, Clinical and Neurobiological Evidence, and Implications: Dissociative Subtype of PTSD. Depress. Anxiety 2012, 29, 701–708. [Google Scholar] [CrossRef]
- Lanius Ruth, A.; Frewen Paul, A.; Vermetten, E.; Yehuda, R. Fear conditioning and early life vulnerabilities: Two distinct pathways of emotional dysregulation and brain dysfunction in PTSD. Eur. J. Psychotraumatol. 2010, 1, 5467. [Google Scholar] [CrossRef] [PubMed]
- Nuss, P. Anxiety disorders and GABA neurotransmission: A disturbance of modulation. Neuropsychiatr. Dis. Treat. 2015, 11, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalueff, A.V.; Nutt, D.J. Role of GABA in anxiety and depression. Depress. Anxiety 2007, 24, 495–517. [Google Scholar] [CrossRef] [PubMed]
- Olivier, J.D.A.; Vinkers, C.H.; Olivier, B. The role of the serotonergic and GABA system in translational approaches in drug discovery for anxiety disorders. Front. Pharmacol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.S.; Rudolph, U. Anxiety and depression: Mouse genetics and pharmacological approaches to the role of GABA(A) receptor subtypes. Neuropharmacology 2012, 62, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Vinkers, C.H.; van Oorschot, R.; Nielsen, E.Ø.; Cook, J.M.; Hansen, H.H.; Groenink, L.; Olivier, B.; Mirza, N.R. GABAA Receptor α Subunits Differentially Contribute to Diazepam Tolerance after Chronic Treatment. PLoS ONE 2012, 7, e43054. [Google Scholar] [CrossRef]
- Sigel, E.; Steinmann, M.E. Structure, Function, and Modulation of GABA A Receptors. J. Biol. Chem. 2012, 287, 40224–40231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKernan, R.M.; Whiting, P.J. Which GABAA-receptor subtypes really occur in the brain? Trends Neurosci. 1996, 19, 139–143. [Google Scholar] [CrossRef]
- Jacobson-Pick, S.; Elkobi, A.; Vander, S.; Rosenblum, K.; Richter-Levin, G. Juvenile stress-induced alteration of maturation of the GABAA receptor α subunit in the rat. Int. J. Neuropsychopharmacol. 2008, 11. [Google Scholar] [CrossRef] [Green Version]
- Puryear, C.B.; Brooks, J.; Tan, L.; Smith, K.; Li, Y.; Cunningham, J.; Todtenkopf, M.S.; Dean, R.L.; Sanchez, C. Opioid receptor modulation of neural circuits in depression: What can be learned from preclinical data? Neurosci. Biobehav. Rev. 2020, 108, 658–678. [Google Scholar] [CrossRef]
- Van’t Veer, A.; Carlezon, W.A. Role of kappa-opioid receptors in stress and anxiety-related behavior. Psychopharmacology (Berl.) 2013, 229, 435–452. [Google Scholar] [CrossRef] [PubMed]
- Lutz, P.-E.; Gross, J.A.; Dhir, S.K.; Maussion, G.; Yang, J.; Bramoulle, A.; Meaney, M.J.; Turecki, G. Epigenetic Regulation of the Kappa Opioid Receptor by Child Abuse. Biol. Psychiatry 2018, 84, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Karkhanis, A.N.; Rose, J.H.; Weiner, J.L.; Jones, S.R. Early-Life Social Isolation Stress Increases Kappa Opioid Receptor Responsiveness and Downregulates the Dopamine System. Neuropsychopharmacology 2016, 41, 2263–2274. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.J.; Husbands, S.M. Targeting opioid receptor signaling in depression: Do we need selective κ opioid receptor antagonists? Neuronal Signal. 2018, 2, NS20170145. [Google Scholar] [CrossRef] [Green Version]
- Carlezon, W.A.; Béguin, C.; DiNieri, J.A.; Baumann, M.H.; Richards, M.R.; Todtenkopf, M.S.; Rothman, R.B.; Ma, Z.; Lee, D.Y.-W.; Cohen, B.M. Depressive-Like Effects of the κ-Opioid Receptor Agonist Salvinorin A on Behavior and Neurochemistry in Rats. J. Pharmacol. Exp. Ther. 2006, 316, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Knoll, A.T.; Meloni, E.G.; Thomas, J.B.; Carroll, F.I.; Carlezon, W.A. Anxiolytic-Like Effects of κ-Opioid Receptor Antagonists in Models of Unlearned and Learned Fear in Rats. J. Pharmacol. Exp. Ther. 2007, 323, 838–845. [Google Scholar] [CrossRef]
- Horovitz, O.; Tsoory, M.M.; Yovell, Y.; Richter-Levin, G. A rat model of pre-puberty (Juvenile) stress-induced predisposition to stress-related disorders: Sex similarities and sex differences in effects and symptoms. World J. Biol. Psychiatry 2014, 15, 36–48. [Google Scholar] [CrossRef]
- Savonenko, A.V.; Brush, F.R.; Zieliński, K. How do rats cope with the two-way escape problem in a homogeneous shuttle box? Acta Neurobiol. Exp. (Warsz.) 1999, 59, 145–157. [Google Scholar]
- Brydges, N.M.; Hall, L.; Nicolson, R.; Holmes, M.C.; Hall, J. The Effects of Juvenile Stress on Anxiety, Cognitive Bias and Decision Making in Adulthood: A Rat Model. PLoS ONE 2012, 7, e48143. [Google Scholar] [CrossRef] [Green Version]
- Bondar, N.P.; Lepeshko, A.A.; Reshetnikov, V.V. Effects of Early-Life Stress on Social and Anxiety-Like Behaviors in Adult Mice: Sex-Specific Effects. Behav. Neurol. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, R.J.; McKittrick, C.R.; Blanchard, D.C. Animal models of social stress: Effects on behavior and brain neurochemical systems. Physiol. Behav. 2001, 73, 261–271. [Google Scholar] [CrossRef]
- Elharrar, E.; Warhaftig, G.; Issler, O.; Sztainberg, Y.; Dikshtein, Y.; Zahut, R.; Redlus, L.; Chen, A.; Yadid, G. Overexpression of Corticotropin-Releasing Factor Receptor Type 2 in the Bed Nucleus of Stria Terminalis Improves Posttraumatic Stress Disorder-like Symptoms in a Model of Incubation of Fear. Biol. Psychiatry 2013, 74, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Panksepp, J. Affective consciousness: Core emotional feelings in animals and humans. Conscious. Cogn. 2005, 14, 30–80. [Google Scholar] [CrossRef] [PubMed]
- Baptista-de-Souza, D.; Pelarin, V.; Canto-de-Souza, L.; Nunes-de-Souza, R.L.; Canto-de-Souza, A. Interplay between 5-HT2C and 5-HT1A receptors in the dorsal periaqueductal gray in the modulation of fear-induced antinociception in mice. Neuropharmacology 2018, 140, 100–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, R.; de Oliveira, R.C.; Falconi-Sobrinho, L.L.; da Silva Soares, R.; Coimbra, N.C. 5-Hydroxytryptamine 2A/2C receptors of nucleus raphe magnus and gigantocellularis/paragigantocellularis pars α reticular nuclei modulate the unconditioned fear-induced antinociception evoked by electrical stimulation of deep layers of the superior colliculus and dorsal periaqueductal grey matter. Behav. Brain Res. 2017, 316, 294–304. [Google Scholar] [CrossRef]
- Chen, Q.; Shao, C.; Zhou, H.; Ma, R.; Jiang, P.; Yang, K. Differential sensitivity of presynaptic and postsynaptic GABAB receptors in rat ventrolateral periaqueductal gray. Neuroreport 2017, 28, 1221–1224. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: Boston, MA, USA; Elsevier: Amsterdam, The Netherlands, 2007; ISBN 978-0-12-547612-6. [Google Scholar]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]






| Observed | Predicted | ||||
|---|---|---|---|---|---|
| Control | Juvenile | Adult | Juvenile + Adult | % Correct | |
| Control | 13 | 0 | 0 | 0 | 100% |
| Juv stress | 0 | 13 | 1 | 0 | 92.9% |
| Adult stress | 0 | 1 | 10 | 2 | 76.9% |
| J+A stress | 0 | 0 | 1 | 13 | 92.9% |
| Overall % | 24.1% | 25.9% | 22.2% | 27.8% | 90.7% |
| χ2(30) = 113, p < 0.001 | |||||
| Explanatory Variables | Juv Stress | Adult Stress | J+A Stress | ||||
|---|---|---|---|---|---|---|---|
| Region+ Receptor | Sig. | Odds Ratio | 95% CI | Odds Ratio | 95% CI | Odds Ratio | 95% C |
| NuAc core GABAAα1 | 0.000 | 1.047 | 0.824–1.331 | 1.121 | 0.873–1.438 | 1.158 | 0.896–1.496 |
| dPAG GABAAα1 | 0.000 | 0.874 | 0.780–0.978 | 0.970 | 0.885–1.063 | 1.020 | 0.956–1.087 |
| vPAG GABAAα1 | 0.052 | 1.042 | 0.884–1.228 | 1.103 | 0.927–1.312 | 1.104 | 0.929–1.314 |
| PRL GABAAα2 | 0.005 | 0.897 | 0.715–1.124 | 0.840 | 0.663–1.065 | 0.875 | 0.695–1.102 |
| NuAc shell GABAAα2 | 0.049 | 0.987 | 0.877–1.112 | 1.056 | 0.953–1.170 | 1.043 | 0.941–1.155 |
| BLA GABAAα2 | 0.000 | 1.146 | 0.909–1.445 | 1.176 | 0.929–1.488 | 1.187 | 0.936–1.504 |
| vPAG GABAAα2 | 0.000 | 1.240 | 0.894–1.720 | 1.073 | 0.775–1.486 | 1.011 | 0.734–1.394 |
| IL KOR | 0.190 | 0.976 | 0.896–1.063 | 0.971 | 0.887–1.062 | 1.000 | 0.908–1.101 |
| BLA KOR | 0.071 | 1.160 | 0.867–1.552 | 1.149 | 0.862–1.532 | 1.140 | 0.854–1.523 |
| CeA KOR | 0.063 | 0.808 | 0.530–1.231 | 0.793 | 0.515–1.219 | 0.779 | 0.502–1.210 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horovitz, O.; Ardi, Z.; Ashkenazi, S.K.; Ritov, G.; Anunu, R.; Richter-Levin, G. Network Neuromodulation of Opioid and GABAergic Receptors Following a Combination of “Juvenile” and “Adult Stress” in Rats. Int. J. Mol. Sci. 2020, 21, 5422. https://doi.org/10.3390/ijms21155422
Horovitz O, Ardi Z, Ashkenazi SK, Ritov G, Anunu R, Richter-Levin G. Network Neuromodulation of Opioid and GABAergic Receptors Following a Combination of “Juvenile” and “Adult Stress” in Rats. International Journal of Molecular Sciences. 2020; 21(15):5422. https://doi.org/10.3390/ijms21155422
Chicago/Turabian StyleHorovitz, Omer, Ziv Ardi, Shiri Karni Ashkenazi, Gilad Ritov, Rachel Anunu, and Gal Richter-Levin. 2020. "Network Neuromodulation of Opioid and GABAergic Receptors Following a Combination of “Juvenile” and “Adult Stress” in Rats" International Journal of Molecular Sciences 21, no. 15: 5422. https://doi.org/10.3390/ijms21155422
APA StyleHorovitz, O., Ardi, Z., Ashkenazi, S. K., Ritov, G., Anunu, R., & Richter-Levin, G. (2020). Network Neuromodulation of Opioid and GABAergic Receptors Following a Combination of “Juvenile” and “Adult Stress” in Rats. International Journal of Molecular Sciences, 21(15), 5422. https://doi.org/10.3390/ijms21155422