Approaches and Technologies in Male Fertility Preservation


Abstract
:1. Spermatogenesis
2. Cancer, Chemotherapy, and Male Infertility
2.1. Cancer and Male Infertility
2.2. Cancer Patients and Chemotherapy
2.3. Gonadotoxic Agents and Male Infertility
2.4. Recovery of Human Spermatogenesis after Cancer Therapy
3. Options for Male Fertility Preservation
3.1. Cryopreservation of Sperm
3.2. Cryopreservation of Human Testicular Tissue
3.3. Germ Cell Transplantation
3.4. Testicular Tissue Autograft/Xenograft
3.5. Testicular Cell Xenograft
3.6. Organ Culture
3.7. In-Vitro Cultures of SSCs
3.7.1. Two- and Three-Dimensional Culture Systems
3.7.2. 3D Bioprinted Scaffold
3.7.3. Testicular Organoids
3.7.4. Microfluid System and Organ-on-Chip Technology
3.8. Induced Pluripotent Stem Cells
3.9. Protection of Spermatogenesis In Vivo from Harmful Gonadotoxic Therapy
3.10. Pharmacological Agents
3.10.1. AS101
3.10.2. Growth Factors/Cytokines
4. Conclusions and Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Huleihel, M.; Abu Elhija, M.; Lunenfeld, E. In vitro culture of testicular germ cells: Regulatory factors and limitations. Growth Factors 2007, 25, 236–252. [Google Scholar]
- De Kretser, D.M.; de Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8. [Google Scholar]
- Ogawa, T. Spermatogonial transplantation: The principle and possible applications. J. Mol. Med. 2001, 79, 368–374. [Google Scholar]
- Gassei, K.; Orwig, K.E. Experimental methods to preserve male fertility and treat male factor infertility. Fertil. Steril. 2016, 105, 256–266. [Google Scholar]
- Auharek, S.A.; de França, L.R. Postnatal testis development, Sertoli cell proliferation and number of different spermatogonial types in C57BL/6J mice made transiently hypo- and hyperthyroidic during the neonatal period. J. Anat. 2010, 216, 577–588. [Google Scholar]
- Ehmcke, J.; Wistuba, J.; Schlatt, S. Spermatogonial stem cells. Questions, models and perspectives. Hum. Reprod. Update 2006, 12, 275–282. [Google Scholar]
- Michailov, Y.; Lunenfeld, E.; Kapelushnik, J.; Huleihel, M. Leukemia and male infertility: Past, present, and future. Leuk. Lymphoma 2019, 60, 1126–1135. [Google Scholar]
- Williams, D.H.; Karpman, E.; Sander, J.C.; Spiess, P.E.; Pisters, L.L.; Lipshultz, L.I. Pretreatment semen parameters in men with cancer. J. Urol. 2009, 181, 736–740. [Google Scholar]
- Michailov, Y.; Lunenfeld, E.; Kapilushnik, J.; Friedler, S.; Meese, E.; Huleihel, M. Acute Myeloid Leukemia Affects Mouse Sperm Parameters, Spontaneous Acrosome Reaction, and Fertility Capacity. Int. J. Mol. Sci. 2019, 20, 219. [Google Scholar]
- Dohle, G.R. Male infertility in cancer patients: Review of the literature. Int. J. Urol. 2010, 17, 327–331. [Google Scholar]
- Schover, L.R. Motivation for parenthood after cancer: A review. J. Natl. Cancer Inst. Monogr. 2005, 34, 2–5. [Google Scholar]
- Delessard, M.; Saulnier, J.; Rives, A.; Dumont, L.; Rondanino, C.; Rives, N. Exposure to chemotherapy during childhood or adulthood and consequences on spermatogenesis ad male fertility. Int. J. Mol. Sci. 2020, 21, 1454. [Google Scholar]
- Meistrich, M.L. Effects of chemotherapy and radiotherapy on spermatogenesis in humans. Fertil. Steril. 2013, 100, 1180–1186. [Google Scholar]
- Jahnukainen, K.; Ehmcke, J.; Hou, M.; Schlatt, S. Testicular function and fertility preservation in male cancer patients. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 287–302. [Google Scholar]
- Drumond, A.L.; Weng, C.C.; Wang, G.; Chiarini-Garcia, H.; Eras-Garcia, L.; Meistrich, M.L. Effects of multiple doses of cyclophosphamide on mouse testes: Accessing the germ cells lost, and the functional damage of stem cells. Reprod. Toxicol. 2011, 32, 395–406. [Google Scholar]
- Dym, M.; Clermont, Y. Role of spermatogonia in the repair of the seminiferous epithelium following x-irradiation of the rat testis. Am. J. Anat. 1970, 128, 265–282. [Google Scholar]
- Lu, C.C.; Meistrich, M.L. Cytotoxic effects of chemotherapeutic drugs on mouse testis cell. Cancer Res. 1979, 39, 3575–3582. [Google Scholar]
- Kangasniemi, M.; Veromaa, T.I.; Kulmala, J.; Kaipia, A.; Parvinen, M.; Toppari, I. DNA flow cytometry of defined stages of rat seminiferous epithelium: Effects of 3 Gv of high-energy X-irradiation. J. Androl. 1990, 11, 312–317. [Google Scholar]
- Meistrich, M.L.; Hunter, N.R.; Suzuki, N.; Trostle, P.K.; Withers, H.R. Gradual regeneration of mouse testicular stem cells after exposure to ionizing radiation. Radiat. Res. 1978, 74, 349–362. [Google Scholar]
- Meistrich, M.L. Quantitative relation between testicular stem cell survival, sperm production, and fertility in the mouse after treatment with different cytotoxic agents. J. Androl. 1982, 3, 58–68. [Google Scholar]
- Kangasniemi, M.; Huhtaniemi, I.; Meistrich, M.L. Failure of spermatogenesis to recover despite the presence of a spermatogonia in the irradiated LBNF1 rat. Biol. Reprod. 1996, 54, 1200–1208. [Google Scholar]
- Zhang, Z.; Shao, S.; Meistrich, M.L. The radiation-induced block in spermatogonial differentiation is due to damage to the somatic environment, not the germ cells. J. Cell Physiol. 2007, 211, 149–158. [Google Scholar]
- de Rooij, D.G.; van de Kant, H.J.; Dol, R.; Wagemaker, G.; van Buul, P.P.; van Duijn-Goedhart, A.; de Jong, F.H.; Broerse, J.J. Long-term effects of irradiation before adulthood on reproductive function in the male rhesus monkey. Biol. Reprod. 2002, 66, 486–494. [Google Scholar]
- Schover, L.R.; Brey, K.; Lichtin, A.; Lipshultz, L.I.; Jeha, S. Knowledge and experience regarding cancer, infertility, and sperm banking in younger male survivors. J. Clin Oncol. 2002, 20, 1880–1889. [Google Scholar]
- Holoch, P.; Wald, M. Current options for preservation of fertility in the male. Fertil. Steril. 2011, 96, 286–290. [Google Scholar]
- Garcia, A.; Herrero, M.B.; Holzer, H.; Tulandi, T.; Chan, P. Assisted reproductive outcomes of male cancer survivors. J. Cancer Surviv. 2015, 9, 208–214. [Google Scholar]
- Pelzman, D.L.; Orwig, K.E.; Hwang, K. Progress in Translational Reproductive Science: Testicular Tissue Transplantation and in Vitro Spermatogenesis. Fertil. Steril. 2020, 113, 500–509. [Google Scholar] [CrossRef]
- Sharma, S.; Wistuba, J.; Pock, T.; Schlatt, S.; Neuhaus, N. Spermatogonial Stem Cells: Updates from Specification to Clinical Relevance. Hum. Reprod. Update 2019, 25, 275–297. [Google Scholar] [CrossRef]
- Komeya, M.; Sato, T.; Ogawa, T. In Vitro Spermatogenesis: A Century-Long Research Journey, Still Half Way Around. Reprod. Med. Biol. 2018, 17, 407–420. [Google Scholar] [CrossRef]
- Zarandi, N.P.; Galdon, G.; Kogan, S.; Atala, A.; Sadri-Ardekani, H. Cryostorage of immature and mature human testis tissue to preserve spermatogonial stem cells (SSCs): A systematic review of current experiences toward clinical applications. Stem. Cells Cloning 2018, 11, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Wyns, C.; Curaba, M.; Petit, S.; Vanabelle, B.; Laurent, P.; Wese, J.F.; Donnez, J. Management of Fertility Preservation in Prepubertal Patients: 5 Years’ Experience at the Catholic University of Louvain. Hum. Reprod. 2011, 26, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Uijldert, M.; Meißner, A.; De Melker, A.A.; Van Pelt, A.M.M.; Van De Wetering, M.D.; Van Rijn, R.R.; Van Wely, M.; Van Der Veen, F.; Repping, S. Development of the Testis in Pre-Pubertal Boys With Cancer After Biopsy for Fertility Preservation. Hum. Reprod. 2017, 32, 2366–2372. [Google Scholar] [CrossRef]
- Mulder, C.L.; Catsburg, L.A.; Zheng, Y.; de Winter-Korver, C.M.; Van Daalen, S.K.; Van Wely, M.; Pals, S.; Repping, S.; Van Pelt, A.M. Long-term Health in Recipients of Transplanted in Vitro Propagated Spermatogonial. Stem. Cells Hum. Reprod. 2018, 33, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Brinster, R.L.; Avarbock, M.R. Germline transmission of donor haplotype following spermatogonial transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11303–11307. [Google Scholar]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis following male germ-cell transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar]
- Honaramooz, A.; Megee, S.O.; Dobrinski, I. Germ cell transplantation in pigs. Biol. Reprod. 2002, 66, 21–28. [Google Scholar]
- Honaramooz, A.; Li, M.; Penedo, C.T.; Meyers, S.; Dobrinski, I. Accelerated maturation of primate testis by xenografting into mice. Biol. Reprod. 2004, 70, 1500–1503. [Google Scholar]
- Schlatt, S.; Foppiani, L.; Rolf, C.; Weinbauer, G.F.; Nieschlag, E. Germ cell transplantation into X-irradiated monkey testes. Hum. Reprod. 2002, 17, 55–62. [Google Scholar]
- Shinohara, T.; Inoue, K.; Ogonuki, N.; Kanatsu-Shinohara, M.; Miki, H.; Nakata, K.; Kurome, M.; Nagashima, H.; Toyokuni, S.; Kogishi, K.; et al. Birth of offspring following transplantation of cryopreserved immature testicular pieces and in vitro microinsemination. Hum. Reprod. 2002, 17, 3039–3045. [Google Scholar]
- Oatley, J.M.; de Avila, D.M.; Reeves, J.J.; McLean, D.J. Spermatogenesis and germ cell transgene expression in xenografted bovine testicular tissue. Biol. Reprod. 2004, 71, 494–501. [Google Scholar]
- Oatley, J.M.; Reeves, J.J.; McLean, D.J. Establishment of spermatogenesis in neonatal bovine testicular tissue following ectopic xenografting varies with donor age. Biol. Reprod. 2005, 72, 358–364. [Google Scholar]
- Snedaker, A.K.; Honaramooz, A.; Dobrinski, I. A game of cat and mouse: Xenografting of testis tissue from domestic kittens results in complete cat spermatogenesis in a mouse host. J. Androl. 2004, 25, 926–930. [Google Scholar]
- Rathi, R.; Honaramooz, A.; Zeng, W.; Turner, R.; Dobrinski, I. Germ cell development in equine testis tissue xenografted into mice. Reproduction 2006, 131, 1091–1098. [Google Scholar]
- Shinohara, T.; Orwig, K.E.; Avarbock, M.R.; Brinster, R. Remodeling of the postnatal mouse testis is accompanied by dramatic changes in stem cell number and niche accessibility. Proc. Natl. Acad. Sci. USA 2001, 98, 6186–6191. [Google Scholar]
- HHermann, B.P.; Sukhwani, M.; Winkler, F.; Pascarella, J.N.; Peters, K.A.; Sheng, Y.; Valli, H.; Rodriguez, M.; Ezzelarab, M.; Dargo, G.; et al. Spermatogonial stem cell transplantation into rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell 2012, 11, 715–726. [Google Scholar]
- Ryu, B.Y.; Orwig, K.E.; Avarbock, M.R.; Brinster, R.L. Stem cell and niche development in the postnatal rat testis. Dev. Biol. 2003, 263, 253–263. [Google Scholar]
- Dobrinski, I.; Avarbock, M.R.; Brinster, R.L. Transplantation of germ cells from rabbits and dogs into mouse testes. Biol. Reprod. 1999, 61, 1331–1339. [Google Scholar]
- Dobrinski, I.; Avarbock, M.R.; Brinster, R.L. Germ cell transplantation from large domestic animals into mouse testes. Mol. Reprod. Dev. 2000, 57, 270–279. [Google Scholar]
- Hermann, B.P.; Sukhwani, M.; Lin, C.C.; Sheng, Y.; Tomko, J.; Rodriguez, M.; Shuttleworth, J.J.; McFarland, D.; Hobbs, R.M.; Pandolfi, P.P.; et al. Characterization, cryopreservation, and ablation of spermatogonial stem cells in adult rhesus macaques. Stem. Cells 2007, 25, 2330–2338. [Google Scholar]
- Dovey, S.L.; Valli, H.; Hermann, B.P.; Sukhwani, M.; Donohue, J.; Castro, C.A.; Chu, T.; Sanfilippo, J.S.; Orwig, K.E. Eliminating malignant contamination from therapeutic human spermatogonial stem cells. J. Clin. Invest. 2013, 123, 1833–1843. [Google Scholar]
- Valli, H.; Sukhwani, M.; Dovey, S.L.; Peters, K.A.; Donohue, J.; Castro, C.A.; Chu, T.; Marshall, G.R.; Orwig, K.E. Fluorescence- and magnetic-activated cell sorting strategies to isolate and enrich human spermatogonial stem cells. Fertil. Steril. 2014, 102, 566–580. [Google Scholar]
- Wu, X.; Goodyear, S.M.; Abramowitz, L.K.; Bartolomei, M.S.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. Fertile offspring derived from mouse spermatogonial stem cells cryopreserved for more than 14 years. Hum. Reprod. 2012, 27, 1249–1259. [Google Scholar]
- Kubota, H.; Brinster, R.L. Spermatogonial stem cells. Biol. Reprod. 2018, 99, 52–74. [Google Scholar]
- Shetty, G.; Mitchell, J.M.; Meyer, J.M.; Wu, Z.; Lam, T.N.; Phan, T.T.; Zhang, J.; Hill, L.; Tailor, R.C.; Peters, K.A.; et al. Restoration of Functional Sperm Production in Irradiated Pubertal Rhesus Monkeys by Spermatogonial Stem Cell Transplantation. Andrology 2020. [Google Scholar] [CrossRef]
- Brook, P.F.; Radford, J.A.; Shalet, S.M.; Joyce, A.D.; Gosden, R.G. solation of germ cells from human testicular tissue for low temperature storage and autotransplantation. Fertil. Steril. 2001, I75, 269–274. [Google Scholar]
- Radford, J. Restoration of fertility after treatment for cancer. Horm. Res. 2003, 59 (Suppl. 1), 21–23. [Google Scholar]
- Nagano, M.; Avarbock, M.R.; Brinster, R.L. Pattern and kinetics of mouse donor spermatogonial stem cell colonization in recipient testes. Biol. Reprod. 1999, 60, 1429–1436. [Google Scholar]
- Nagano, M.; Ryu, B.Y.; Brinster, C.J.; Avarbock, M.R.; Brinster, R.L. Maintenance of mouse male germ line stem cells in vitro. Biol. Reprod. 2003, 68, 2207–2214. [Google Scholar]
- Wu, X.; Schmidt, J.A.; Avarbock, M.R.; Tobias, J.W.; Carlson, C.A.; Kolon, T.F.; Ginsberg, J.P.; Brinster, R.L. Prepubertal human spermatogonia and mouse gonocytes share conserved gene expression of germline stem cell regulatory molecules. Proc. Natl. Acad. Sci. USA 2009, 106, 21672–21677. [Google Scholar]
- Takashima, S.; Shinohara, T. Culture and transplantation of spermatogonial stem cells. Stem Cell Res. 2018, 29, 46–55. [Google Scholar]
- Ogawa, T.; Dobrinski, I.; Avarbock, M.R.; Brinster, R.L. Transplantation of male germ line stem cells restores fertility in infertile mice. Nat. Med. 2000, 6, 29–34. [Google Scholar]
- Fujita, K.; Tsujimura, A.; Miyagawa, Y.; Kiuchi, H.; Matsuoka, Y.; Takao, T.; Takada, S.; Nonomura, N.; Okuyama, A. Isolation of germ cells from leukemia and lymphoma cells in a human in vitro model: Potential clinical application for restoring human fertility after anticancer therapy. Cancer Res. 2006, 66, 11166–11171. [Google Scholar]
- Geens, M.; Goossens, E.; Tournaye, H. Cell selection by selective matrix adhesion is not sufficiently efficient for complete malignant cell depletion from contaminated human testicular cell suspensions. Fertil. Steril. 2011, 787, 91–95. [Google Scholar]
- Rodriguez-Sos, J.R.; Dobrinski, I. Recent development in testis tissue xenografting. Reproduction 2009, 138, 187–194. [Google Scholar]
- Galdon, G.; Atala, A.; Sadri-Ardekani, H. In Vitro Spermatogenesis: How Far from Clinical Application? Curr Urol Rep. 2016, 17, 49. [Google Scholar] [CrossRef]
- Honaramooz, A.; Snedaker, A.; Boiani, M.; Scholer, H.; Dobrinski, I.; Schlatt, S. Sperm from neonatal mammalian testes grafted in mice. Nature 2002, 418, 778–781. [Google Scholar]
- Luetjens, C.M.; Stukenborg, J.B.; Nieschlag, E.; Simoni, M.; Wistuba, J. Complete spermatogenesis in orthotopic but not in ectopic transplants of autologously grafted marmoset testicular tissue. Endocrinology 2008, 149, 1736–1747. [Google Scholar]
- Fayomi, A.P.; Peters, K.; Sukhwani, M.; Valli-Pulaski, H.; Shetty, G.; Meistrich, M.L.; Houser, L.; Robertson, N.; Roberts, V.; Ramsey, C.; et al. Autologous grafting of cryopreserved prepubertal rhesus testis produces sperm and offspring. Science 2019, 363, 1314–1319. [Google Scholar]
- Wyns, C.; Van Langendonckt, A.; Wese, F.X.; Donnez, J.; Curaba, M. Long-term spermatogonial survival in cryopreserved and xenografted immature human testicular tissue. Hum. Reprod. 2008, 23, 2402–2414. [Google Scholar]
- Geens, M.; De Block, G.; Goossens, E.; Frederickx, V.; Van Steirteghem, A.; Tournaye, H. Spermatogonial survival after grafting human testicular tissue to immunodeficient mice. Hum. Reprod. 2006, 21, 390–396. [Google Scholar]
- Schlatt, S.; Honaramooz, A.; Ehmcke, J.; Goebell, P.J.; Rübben, H.; Dhir, R.; Dobrinski, I.; Patrizio, P. Limited survival of adult human testicular tissue as ectopic xenograft. Hum. Reprod. 2006, 21, 384–389. [Google Scholar]
- Goossens, E.; Geens, M.; De Block, G.; Tournaye, H. Spermatogonial survival in long-term human prepubertal xenografts. Fertil. Steril. 2008, 90, 2019–2022. [Google Scholar]
- Honaramooz, A.; Megee, S.O.; Rathi, R.; Dobrinski, I. Building a testis: Formation of functional testis tissue after transplantation of isolated porcine (Sus scrofa) testis cells. Biol. Reprod. 2007, 76, 43–47. [Google Scholar]
- Kita, K.; Watanabe, T.; Ohsaka, K.; Hayashi, H.; Kubota, Y.; Nagashima, Y.; Aoki, I.; Taniguchi, H.; Noce, T.; Inoue, K.; et al. Production of functional spermatids from mouse germline stem cells in ectopically reconstituted seminiferous tubules. Biol. Reprod. 2007, 76, 211–217. [Google Scholar]
- Arregui, L.; Rathi, R.; Megee, S.O.; Honaramooz, A.; Gomendio, M.; Roldan, E.R.; Dobrinski, I. Xenografting of sheep testis tissue and isolated cells as a model for preservation of genetic material from endangered ungulates. Reproduction 2008, 136, 85–93. [Google Scholar]
- Shetty, G.; Mitchell, J.M.; Lam, T.N.A.; Wu, Z.; Zhang, J.; Hill, L.; Tailor, R.C.; Peters, K.A.; Penedo, M.C.; Orwig, K.E.; et al. Donor spermatogenesis in de novo formed seminiferous tubules from transplanted testicular cells in rhesus monkey testis. Hum. Reprod. 2018, 33, 2249–2255. [Google Scholar]
- Mahmoud, H. Spermatogenesis in artificial three-dimensional system. Stem. Cells 2012, 30, 2355–2360. [Google Scholar]
- Steinberger, A.; Steinberger, E.; Perloff, W.H. Mammalian testes in organ culture. Exp. Cell Res. 1964, 36, 19–27. [Google Scholar]
- Steinberger, A.; Steinberger, E. Factors affecting spermatogenesis in organ cultures of mammalian testes. J. Reprod. Fertil. 1967, 2, 117–124. [Google Scholar]
- Ogawa, T. In vitro spermatogenesis: The dawn of a new era in the study of male infertility. Int. J. Urol. 2012, 19, 282–283. [Google Scholar]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y. and Ogawa, T. In vitro production of functional sperm in cultured neonatal mouse testes. Nature 2011, 471, 504–508. [Google Scholar]
- Dumont, L.; Arkoun, B.; Jumeau, F.; Milazzo, J.P.; Bironneau, A.; Liot, D.; Wils, J.; Rondanino, C.; Rives, N. Assessment of the optimal vitrification protocol for pre-pubertal mice testes leading to successful in vitro production of flagellated spermatozoa. Andrology 2015, 3, 611–625. [Google Scholar]
- Medrano, J.V.; Vilanova-Perez, T.; Fornes-Ferrer, V.; Navarro-Gomezlechon, A.; Martínez-Triguero, M.L.; García, S.; Gómez-Chacón, J.; Povo, I.; Pellicer, A.; Andrés, M.M.; et al. Influence of temperature, serum, and gonadotropin supplementation in short- and long-term organotypic culture of human immature testicular tissue. Fertil. Steril. 2018, 110, 1045–1057.e3. [Google Scholar]
- Yang, S.; Ping, P.; Ma, M.; Li, P.; Tian, R.; Yang, H.; Liu, Y.; Gong, Y.; Zhang, Z.; Li, Z.; et al. Generation of haploid spermatids with fertilization and development capacity from human spermatogonial stem cells of cryptorchid patients. Stem Cell Rep. 2014, 3, 663–675. [Google Scholar]
- De Michele, F.; Poels, J.; Weerens, L.; Petit, C.; Evrard, Z.; Ambroise, J.; Gruson, D.; Wyns, C. Preserved seminiferous tubule integrity with spermatogonial survival and induction of Sertoli and Leydig cell maturation after long-term organotypic culture of prepubertal human testicular tissue. Hum. Reprod. 2017, 32, 32–45. [Google Scholar]
- Lee, J.H.; Kim, H.J.; Kim, H.; Lee, S.J.; Gye, M.C. In vitro spermatogenesis by threedimensional culture of rat testicular cells in collagen gel matrix. Biomaterials 2006, 27, 2845–2853. [Google Scholar]
- Stukenborg, J.B.; Wistuba, J.; Luetjens, C.M.; Elhija, M.A.; Huleihel, M.; Lunenfeld, E.; Gromoll, J.; Nieschlag, E.; Schlatt, S. Coculture of spermatogonia with somatic cells in a novel three dimensional soft-agar-culture-system. J. Androl. 2008, 29, 312–329. [Google Scholar]
- Sadri-Ardekani, H.; Mizrak, S.C.; van Daalen, S.K.; Korver, C.M.; Roepers-Gajadien, H.L.; Koruji, M.; Hovingh, S.; de Reijke, T.M.; de la Rosette, J.J.; van der Veen, F.; et al. Propagation of human spermatogonial stem cells in vitro. JAMA 2009, 302, 2127–2134. [Google Scholar]
- Sadri-Ardekani, H.; Akhondi, M.A.; van der Veen, F.; Repping, S.; van Pelt, A.M. In Vitro Propagation of Human Prepubertal Spermatogonial Stem. Cells. JAMA 2011, 305, 2416–2418. [Google Scholar]
- Stukenborg, J.B.; Schlatt, S.; Simoni, M.; Yeung, C.H.; Elhija, M.A.; Luetjens, C.M.; Huleihel, M.; Wistuba, J. New horizons for in vitro spermatogenesis? An update on novel three-dimensional culture systems as tools for meiotic and postmeiotic differentiation of testicular germ cells. Mol. Hum. Reprod. 2009, 15, 971–980. [Google Scholar]
- Huleihel, M.; Nourashrafeddin, S.; Plant, T.M. Application of three dimensional culture systems to study mammalian spermatogenesis, with an emphasis on the rhesus monkey (Macaca mulatta). Asian J. Androl. 2015, 17, 972–980. [Google Scholar]
- Abu Elhija, M.; Lunenfeld, E.; Schlatt, S.; Huleihel, M. Differentiation of murine male germ cells to spermatozoa in a soft agar culture system. Asian J. Androl. 2012, 14, 285–293. [Google Scholar]
- AbuMadighem, A.; Solomon, R.; Stepanovsky, A.; Kapelushnik, J.; Shi, Q.; Meese, E.; Lunenfeld, E.; Huleihel, M. Development of spermatogenesis in vitro in three-dimensional culture from spermatogonial cells of busulfan-treated immature mice. Int. J. Mol. Sci. 2018, 19, 3804–3820. [Google Scholar]
- Abofoul-Azab, M.; Lunenfeld, E.; Levitas, E.; Zeadna, A.; Younis, J.; Bar-Ami, S.; Huleihel, M. Identification of Premeiotic, Meiotic, and Postmeiotic Cells in Testicular Biopsies Without Sperm from Sertoli Cell-Only Syndrome Patients. Int. J. Mol. Sci. 2019, 20, 470. [Google Scholar] [CrossRef] [Green Version]
- Huleihel, N.; Azab, M.; Levitas, E.; Fisch, B.; Pinkas, H.; Stein, A.; Younis, J.S.; Bar-Ami, S.; Orvieto, R.; Ziadna, A.; et al. The capacity to induce generation of premeiotic and meiotic cells from biopsies without sperm from non-obstructive azoospermic and Klinefelter syndrome patients were different under in vitro culture conditions. Fertil. Steril. 2015, 104, e91. [Google Scholar]
- Azab, M.; Lunenfeld, E.; Kapelushnik, J.; Huleihel, M. Fertility reservation of pre-pubertal cancer patient boys before aggressive chemotherapy. Preliminary results from in vitro cultures of fresh testicular tissue from three pre-pubertal patients. Fertil. Steril. 2013, 100, S63. [Google Scholar]
- Valli-Pulaski, H.; Peters, K.A.; Gassei, K.; Steime, S.R.; Sukhwani, M.; Hermann, B.P.; Dwomor, L.; David, S.; Fayomi, A.P.; Munyoki, S.K.; et al. Testicular tissue cryopreservation: 8 years of experience from a coordinated network of academic centers. Hum. Reprod. 2019, 2019, 1–12. [Google Scholar] [CrossRef]
- Baert, Y.; Dvorakova-Hortova, K.; Margaryan, H.; Goossens, E. Mouse in Vitro Spermatogenesis on Alginate-Based 3D Bioprinted Scaffolds. Biofabrication 2019, 11, 035011. [Google Scholar] [CrossRef]
- Alves-Lopes, J.P.; Stukenborg, J.B. Testicular organoids: A new model to study the testicular microenvironment in vitro? Hum. Reprod. Update 2018, 24, 176–191. [Google Scholar]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a dish: Modeling development and disease using organoid technologies. Science 2014, 345, 1247125. [Google Scholar]
- Clevers, H. Modeling development and disease with organoids. Cell 2016, 165, 1586–1597. [Google Scholar]
- Huch, M.; Knoblich, J.A.; Lutolf, M.P.; Martinez-Arias, A. The hope and the hype of organoid research. Development 2017, 144, 938–941. [Google Scholar]
- Pendergraft, S.S.; Sadri-Ardekani, H.; Atala, A.; Bishop, C.E. Three-dimensional testicular organoid: A novel tool for the study of human spermatogenesis and gonadotoxicity in vitro dagger. Biol. Reprod. 2017, 96, 720–732. [Google Scholar]
- Baert, Y.; De Kock, J.; Alves-Lopes, J.P.; Söder, O.; Stukenborg, J.-B.; Goossens, E. Primary human testicular cells self-organize into organoids with testicular properties. Stem Cell Rep. 2017, 1, 30–38. [Google Scholar]
- Alves-Lopes, J.P.; Soder, O.; Stukenborg, J.B. Testicular organoid generation by a novel in vitro three-layer gradient system. Biomaterials 2017, 130, 76–89. [Google Scholar]
- Topraggaleh, T.R.; Valojerdi, M.R.; Montazeri, L.; Baharvand, H. A testis-derived macroporous 3D scaffold as a platform for the generation of mouse testicular organoids. Biomater. Sci. 2019, 7, 1422–1436. [Google Scholar]
- Komeya, M.; Kimura, H.; Nakamura, H.; Yokonishi, T.; Sato, T.; Kojima, K.; Hayashi, K.; Katagiri, K.; Yamanaka, H.; Sanjo, H.; et al. Long-term ex vivo maintenance of testis tissues producing fertile sperm in a microfluidic device. Sci. Rep. 2016, 6, 21472. [Google Scholar]
- Komeya, M.; Hayashi, K.; Nakamura, H.; Yamanaka, H.; Sanjo, H.; Kojima, K.; Sato, T.; Yao, M.; Kimura, H.; Fujii, T.; et al. Pumpless Microfluidic System Driven by Hydrostatic Pressure Induces and Maintains Mouse Spermatogenesis in Vitro. Sci. Rep. 2017, 7, 15459. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, H.; Komeya, M.; Nakamura, H.; Sanjo, H.; Sato, T.; Yao, M.; Kimura, H.; Fujii, T.; Ogawa, T. A monolayer microfluidic device supporting mouse spermatogenesis with improved visibility. Biochem. Biophys. Res. Commun. 2018, 500, 885–891. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar]
- Hou, J.; Yang, S.; Yang, H.; Liu, Y.; Hai, Y.; Chen, Z.; Guo, Y.; Gong, Y.; Gao, W.-Q.; Li, Z.; et al. Generation of male differentiated germ cells from various types of stem cells. Reproduction 2014, 147, R179–R188. [Google Scholar]
- Gauthier-Fisher, A.; Kauffman, A.; Librach, C.L. Potential use of stem cells for fertility preservation. Andrology 2020, 8, 862–878. [Google Scholar]
- Mishra, S.; Kacin, E.; Stamatiadis, P.; Franck, S.; Van der Jeught, M.; Mertes, H.; Pennings, G.; De Sutter, P.; Sermon, K.; Heindryckx, B.; et al. The role of the reprogramming method and pluripotency state in gamete differentiation from patient-specific human pluripotent stem cells. Mol. Hum. Reprod. 2018, 24, 173–184. [Google Scholar]
- Nagamatsu, G.; Hayashi, K. Stem cells, in vitro gametogenesis and male fertility. Reproduction 2017, 154, F79–F91. [Google Scholar]
- Rombaut, C.; Mertes, H.; Heindryckx, B.; Goossens, E. Human in vitro spermatogenesis from pluripotent stem cells: In need of a stepwise differentiation protocol? Mol. Hum. Reprod. 2018, 24, 47–54. [Google Scholar]
- Shimizu, T.; Shiohara, M.; Tai, T.; Nagao, K.; Nakajima, K.; Kobayashi, H. Derivation of integration-free iPSCs from a Klinefelter syndrome patient. Reprod. Med. Biol. 2016, 15, 35–43. [Google Scholar]
- Wang, X.; Liao, T.; Wan, C.; Yang, X.; Zhao, J.; Fu, R.; Yao, Z.; Huang, Y.; Shi, Y.; Chang, G.; et al. Efficient generation of human primordial germ cell-like cells from pluripotent stem cells in a methylcellulose-based 3D system at large scale. PeerJ 2019, 6, e6143. [Google Scholar]
- Kee, K.; Angeles, V.T.; Flores, M.; Nguyen, H.N.; Pera, R.A.R. Human dazl, daz and boule genes modulate primordial germ-cell and haploid gamete formation. Nature 2009, 462, 222–225. [Google Scholar]
- Easley, C.A., IV; Phillips, B.T.; McGuire, M.M.; Barringer, J.M.; Valli, H.; Hermann, B.P.; Simerly, C.R.; Rajkovic, A.; Miki, T.; Orwig, K.E.; et al. Direct differentiation of human pluripotent stem cells into haploid spermatogenic cells. Cell Rep. 2012, 2, 440–446. [Google Scholar]
- Kee, K.; Gonsalves, J.M.; Clark, A.T.; Pera, R.A. Bone morphogenetic proteins induce germ cell differentiation from human embryonic stem cells. Stem. Cells Dev. 2006, 15, 831–837. [Google Scholar]
- Teramura, T.; Takehara, T.; Kawata, N.; Fujinami, N.; Mitani, T.; Takenoshita, M.; Matsumoto, K.; Saeki, K.; Iritani, A.; Sagawa, N.; et al. Primate embryonic stem cells proceed to early gametogenesis in vitro. Cloning Stem. Cells 2007, 9, 144–156. [Google Scholar]
- Park, T.S.; Galic, Z.; Conway, A.E.; Lindgren, A.; Van Handel, B.J.; Magnusson, M.; Richter, L.; Teitell, M.A.; Mikkola, H.K.; Lowry, W.E.; et al. Derivation of primordial germ cells from human embryonic and induced pluripotent stem cells is significantly improved by coculture with human fetal gonadal cells. Stem. Cells 2009, 27, 783–795. [Google Scholar]
- Yamauchi, K.; Hasegawa, K.; Chuma, S.; Nakatsuji, N.; Suemori, H. In vitro germ cell differentiation from cynomolgus monkey embryonic stem cells. PLoS ONE 2009, 4, e5338. [Google Scholar]
- Hayashi, K.; Ohta, H.; Kurimoto, K.; Aramaki, S.; Saitou, M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell 2011, 146, 519–532. [Google Scholar]
- Yuan, Y.; Zhou, Q.; Wan, H.; Shen, B.; Wang, X.; Wang, M.; Feng, C.; Xie, M.; Gu, T.; Zhou, T.; et al. Generation of fertile offspring from Kit(w)/Kit(w/v) mice through differentiation of gene corrected nuclear transfer embryonic stem cells. Cell Res. 2015, 25, 851–863. [Google Scholar]
- Sosa, E.; Chen, D.; Rojas, E.J.; Hennebold, J.D.; Peters, K.A.; Wu, Z.; Lam, T.N.; Mitchell, J.M.; Sukhwani, M.; Tailor, R.C.; et al. Differentiation of primate primordial germ cell-like cells following transplantation into the adult gonadal niche. Nat. Commun. 2018, 9, 5339. [Google Scholar]
- Irie, N.; Surani, M.A. Efficient induction and isolation of human primordial germ cell-like cells from competent human pluripotent stem cells. Methods Mol. Biol. 2017, 1463, 217–226. [Google Scholar]
- Panula, S.; Medrano, J.V.; Kee, K.; Bergström, R.; Nguyen, H.N.; Byers, B.; Wilson, K.D.; Wu, J.C.; Simon, C.; Hovatta, O.; et al. Human germ cell differentiation from fetal- and adult-derived induced pluripotent stem cells. Hum. Mol. Genet. 2011, 20, 752–762. [Google Scholar]
- Medrano, J.V.; Ramathal, C.; Nguyen, H.N.; Simon, C.; Reijo Pera, R.A. Divergent RNA-binding proteins, DAZL and VASA, induce meiotic progression in human germ cells derived in vitro. Stem. Cells 2012, 30, 441–451. [Google Scholar]
- Eguizabal, C.; Montserrat, N.; Vassena, R.; Barragan, M.; Garreta, E.; Garcia-Quevedo, L.; Vidal, F.; Giorgetti, A.; Veiga, A. and Belmonte, J.I. Complete meiosis from human induced pluripotent stem cells. Stem. Cells 2011, 29, 1186–1195. [Google Scholar]
- Shetty, G.; Uthamanthil, R.K.; Zhou, W.; Shao, S.H.; Weng, C.C.; Tailor, R.C.; Hermann, B.P.; Orwig, K.E.; Meistrich, M.L. Hormone Suppression with GnRH Antagonist Promotes Spermatogenic Recovery from Transplanted Spermatogonial Stem Cells in Irradiated Cynomolgus Monkeys. Andrology 2013, 1, 886–898. [Google Scholar]
- Masala, A.; Faedda, R.; Alagna, S.; Satta, A.; Chiarelli, G.; Rovasio, P.P.; Ivaldi, R.; Taras, M.S.; Lai, E.; Bartoli, E. Use of testosterone to prevent cyclophosphamide-induced azoospermia. Ann. Intern. Med. 1997, 126, 292–295. [Google Scholar]
- Thomson, A.B.; Critchley, H.O.; Kelnar, C.J.; Wallace, W.H. Late reproductive sequelae following treatment of childhood cancer and options for fertility preservation. Best Pract. Res. Clin. Endocrinol. Metab. 2002, 16, 311–334. [Google Scholar]
- Ogawa, T.; Dobrinski, I.; Brinster, R.L. Recipient preparation is critical for spermatogonial transplantation in the rat. Tissue Cell 1999, 31, 461–472. [Google Scholar]
- Ogawa, T.; Dobrinski, I.; Avarbock, M.R.; Brinster, R.L. Leuprolide, a gonadotropin-releasing hormone agonist, enhances colonization after spermatogonial transplantation into mouse testes. Tissue Cell 1998, 30, 583–588. [Google Scholar]
- Dobrinski, I.; Ogawa, T.; Avarbock, M.R.; Brinster, R.L. Effect of the GnRH-agonist leuprolide on colonization of recipient testes by donor spermatogonial stem cells after transplantation in mice. Tissue Cell 2001, 33, 200–207. [Google Scholar]
- Ohmura, M.; Ogawa, T.; Ono, M.; Dezawa, M.; Hosaka, M.; Kubota, Y.; Sawada, H. Increment of murine spermatogonial cell number by gonadotropin-releasing hormone analogue is independent of stem cell factor c-kit signal. Biol. Reprod. 2003, 68, 2304–2313. [Google Scholar]
- Zhang, Z.; Renfree, M.B.; Short, R.V. Successful intra- and interspecific male germ cell transplantation in the rat. Biol. Reprod. 2003, 68, 961–967. [Google Scholar]
- Wang, G.; Shao, S.H.; Weng, C.C.; Wei, C.; Meistrich, M.L. Hormonal suppression restores fertility in irradiated mice from both endogenous and donor-derived stem spermatogonia. Toxicol. Sci. 2010, 117, 225–237. [Google Scholar]
- Meng, J.; Holdcraft, R.W.; Shima, J.E.; Griswold, M.D.; Braun, R.E. Androgens regulate the permeability of the blood-testis barrier. Proc. Natl. Acad. Sci. USA 2005, 102, 16696–16700. [Google Scholar]
- Shetty, G.; Wu, Z.; Lam, T.N.A.; Phan, T.T.; Orwig, K.E.; Meistrich, M.L. Effect of hormone modulations on donor-derived spermatogenesis or colonization after syngeneic and xenotransplantation in mice. Andrology 2019, 7, 257–265. [Google Scholar]
- Sredni, B.; Caspi, R.R.; Klein, A.; Kalechman, Y.; Danziger, Y.; Ben, Y.M.; Tamari, T.; Shalit, F.; Albeck, M. A new immunomodulating compound (AS-101) with potential therapeutic application. Nature 1987, 330, 173–176. [Google Scholar]
- Sredni, B. Immunomodulating tellurium compounds as anti-cancer agents. Semin. Cancer Biol. 2012, 22, 60–69. [Google Scholar]
- Sredni, B.; Weil, M.; Khomenok, G.; Lebenthal, I.; Teitz, S.; Mardor, Y.; Ram, Z.; Orenstein, A.; Kershenovich, A.; Michowiz, S.; et al. Ammonium trichloro(dioxoethylene-o,o’)tellurate (AS101) sensitizes tumors to chemotherapy by inhibiting the tumor interleukin 10 autocrine loop. Cancer Res. 2004, 64, 1843–1852. [Google Scholar]
- Sredni, B.; Tichler, T.; Shani, A.; Catane, R.; Kaufman, B.; Strassmann, G.; Albeck, M.; Kalechman, Y. Predominance of TH1 response in tumor-bearing mice and cancer patients treated with AS101. J. Natl. Cancer Inst. 1996, 88, 1276–1284. [Google Scholar]
- Kalechman, Y.; Zuloff, A.; Albeck, M.; Strassmann, G.; Sredni, B. Role of endogenous cytokines secretion in radioprotection conferred by the immunomodulator ammonium trichloro(dioxyethylene-O-O0 )tellurate. Blood 1995, 85, 1555–1561. [Google Scholar]
- Kalechman, Y.; Sredni, B.; Weinstein, T.; Freidkin, I.; Tobar, A.; Albeck, M.; Gafter, U. Production of the novel mesangial autocrine growth factors GDNF and IL-10 is regulated by the immunomodulator AS101. J. Am. Soc. Nephrol. 2003, 14, 620–630. [Google Scholar]
- Carmely, A.; Meirow, D.; Peretz, A.; Albeck, M.; Bartoov, B.; Sredni, B. Protective effect of the immunomodulator AS101 against cyclophosphamide-induced testicular damage in mice. Hum. Reprod. 2009, 24, 1322–1329. [Google Scholar]
- Demetri, G.D.; Griffin, J.D. Granulocyte colony-stimulating factor and its receptor. Blood 1991, 78, 2791–2808. [Google Scholar]
- Shimamura, M.; Kobayashi, Y.; Yuo, A.; Urabe, A.; Okabe, T.; Komatsu, Y.; Itoh, S.; Takaku, F. Effect of human recombinant granulocyte colony stimulating factor on hematopoietic injury in mice induced by 5-fluorouracil. Blood 1987, 69, 353–355. [Google Scholar]
- Uckun, F.M.; Souza, L.; Waddick, K.G.; Wick, M.; Song, C.W. In vivo radioprotective effects of recombinant human granulocyte colony-stimulating factor in lethally irradiated mice. Blood 1990, 75, 638–645. [Google Scholar]
- Kim, J.; Lee, S.; Jeon, B.; Jang, W.; Moon, C.; Kim, S. Protection of spermatogenesis against gamma ray-induced damage by granulocyte colony-stimulating factor in mice. Andrologia 2010, 43, 87–93. [Google Scholar]
- Kotzur, T.; Benavides-Garcia, R.; Mecklenburg, J.; Sanchez, J.R.; Reilly, M.; Hermann, B.P. Granulocyte Colony-Stimulating Factor (G-CSF) Promotes Spermatogenic Regeneration From Surviving Spermatogonia After High-Dose Alkylating Chemotherapy. Reprod. Biol. Endocrinol. 2017, 15, 7. [Google Scholar] [CrossRef] [Green Version]
- Khanlarkhani, N.; Pasbakhsh, P.; Mortezaee, K.; Naji, M.; Amidi, F.; Najafi, A.; Sobhani, A.; Zendedel, A. Effect of Human Recombinant Granulocyte Colony-Stimulating Factor on Rat Busulfan-Induced Testis Injury. J. Mol. Histol. 2016, 47, 59–67. [Google Scholar] [CrossRef]
- Benavides-Garcia, R.; Joachim, R.; Pina, N.A.; Mutoji, K.N.; Reilly, M.A.; Hermann, B.P. Granulocyte colony-stimulating factor prevents loss of spermatogenesis after sterilizing busulfan chemotherapy. Fertil. Steril. 2015, 103, 270–280. [Google Scholar]
- Zhu, Y.; Yin, Q.; Wei, D.; Yang, Z.; Du, Y.; Ma, Y. Autophagy in male reproduction. Sys Biol. Reprod. Med. 2019, 65, 265–272. [Google Scholar]
- Noguchi, M.; Hirata, N.; Tanaka, T.; Suizu, F.; Nakajima, H.; Chiorini, J.A. Autophagy as a modulator of cell death machinery. Cell Death Dis. 2020, 11, 517. [Google Scholar]
- Zhang, M.; Liu, F.; Zhou, P.; Wang, Q.; Xu, C.; Li, Y.; Bian, L.; Liu, Y.; Zhou, J.; Wang, F.; et al. The MTOR signaling pathway regulates macrophage differentiation from mouse myeloid progenitors by inhibiting autophagy. Autophagy 2019, 15, 1150–1162. [Google Scholar]
- Meng, D.; Frank, A.R.; Jewell, J.L. mTOR signaling in stem and progenitor cells. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Bento, C.F.; Renna, M.; Ghislat, G.; Puri, C.; Ashkenazi, A.; Vicinanza, M.; Menzies, F.M.; Rubinsztein, D.C. Mammalian autophagy: How does it work? Annu. Rev. Biochem. 2016, 85, 685–713. [Google Scholar]
- Phadwal, K.; Watson, A.S.; Simon, A.K. Tightrope act: Autophagy in stem cell renewal, differentiation, proliferation, and aging. Cell Mol. Life Sci. 2013, 70, 89–103. [Google Scholar]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar]
- Liu, S.; Huang, L.; Geng, Y.; He, J.; Chen, X.; Xu, H.; Li, R.; Wang, Y.; Ding, Y.; Liu, X. Rapamycin inhibits spermatogenesis by changing the autophagy status through suppressing mechanistic target of rapamycin-p70S6 kinase in male rats. Mol. Med. Rep. 2017, 16, 4029–4037. [Google Scholar]
- Shemesh, I.I.; Rozen-Zvi, B.; Kalechman, Y.; Gafter, U.; Sredni, B. Prevents Diabetic Nephropathy Progression and Mesangial Cell Dysfunction: Regulation of the AKT Downstream Pathway. PLoS ONE 2014, 9, e114287. [Google Scholar] [CrossRef]
- Li, D.; Yang, H.; Nan, H.; Liu, P.; Pang, S.; Zhao, Q.; Karni, R.; Kamps, M.P.; Xu, Y.; Zhou, J.; et al. Identification of key regulatory pathways of myeloid differentiation using an mESC-based karyotypically normal cell model. Blood 2012, 120, 4712–4719. [Google Scholar]


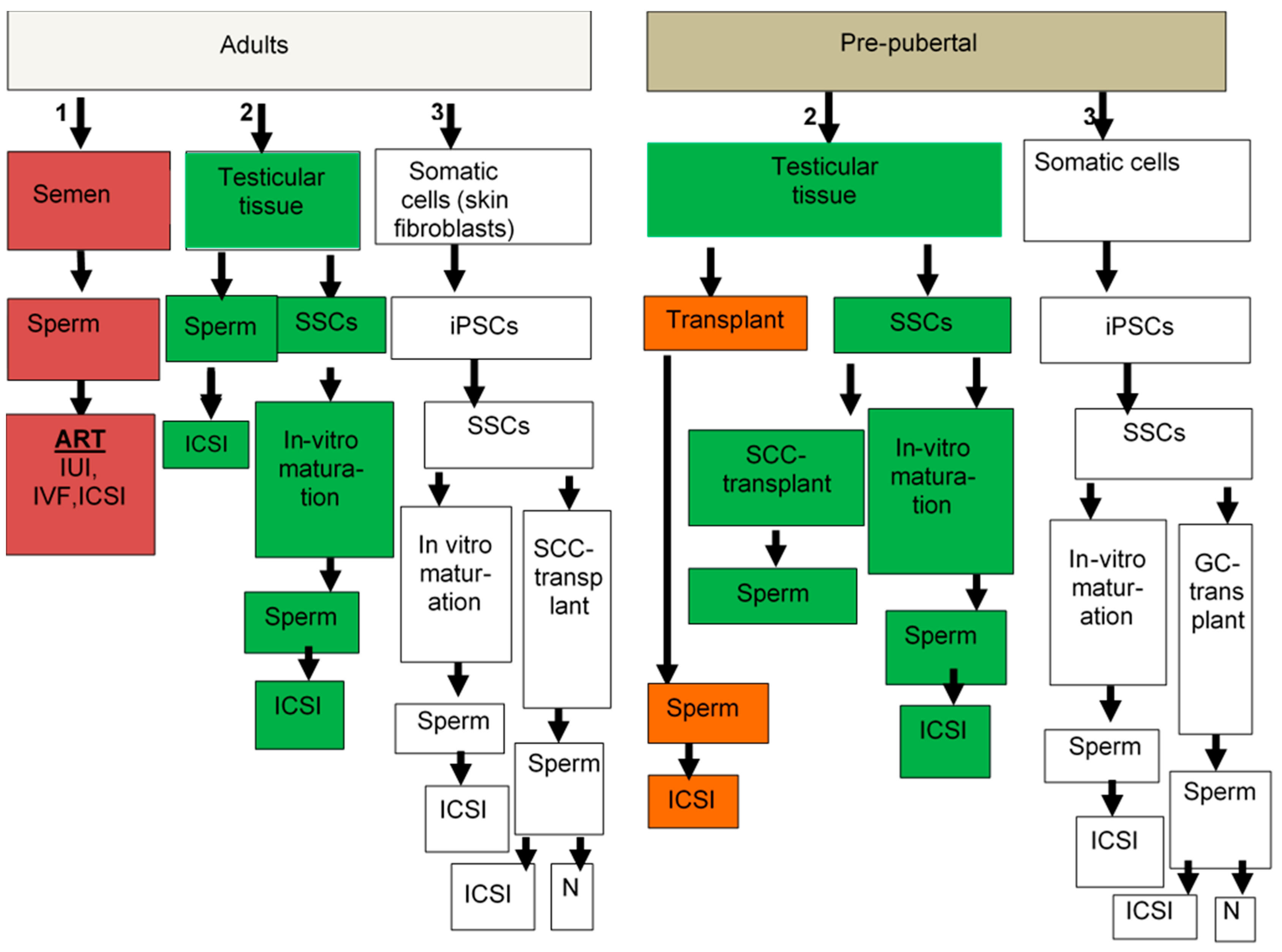{kind=link}
| Approach | Patients | Future Therapeutic Reproductive Strategies | Fertility Options | Advantage | Limitations |
|---|---|---|---|---|---|
| Cryopreservation of sperm | Adults | ART 1–3 | ART | Fertility preservation | (1) Limited quantity and quality of the sperm (2) Not an option for prepubertal patients since they do not yet generate sperm (3) Not recommended after initiation of chemotherapy |
| Testicular biopsy | Pre-pubertal or Adults | Autologous graft 4−6 Autologous germ cell transplantation 4−6 | Intercourse or ART | Fertility preservation and/or restoration | For strategies: (4) Possible contamination with cancer cells * (5) Not yet applicable in human (6) Needs more research for validation, efficiency, and genetic stability (7) Transmissions of mouse virus to human germ line |
| Organ culture 5,6 In-vitro differentiation culture of SSCs to sperm 5,6 Xenograft 5−7 | ART | ||||
| Induced pluripotent stem cells (iPSC) | Adults | In-vitro culture: Differentiation to SSCs 8,9 | Intercourse or ART | Fertility restoration | (8) Needs more research for validation, safety, efficiency, and genetic stability (9) Not yet applicable in humans |
| Differentiation of SSCs to sperm 8,9 | ART | ||||
| Agents to protect spermatogenesis | Pre-pubertal or Adults | Normal fertility 10−12 ART 10−12 | Intercourse or ART | Fertility preservation | (10) Needs more research for possible protective agents (G-CSF, AS101) (11) Needs more research for validation and safety (12) Not yet applicable in humans |
| Gene therapy ** | Adults Or Pubertal | Normal fertility 13−15 In-vitro fertilization 13−15 | Intercourse or ART | Fertility restoration | (13) Needs more research for validation, safety, efficiency, and genetic stability (14) Could be used in testicular somatic cells, but not germ line cells. (15) Not yet applicable in humans |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huleihel, M.; Lunenfeld, E. Approaches and Technologies in Male Fertility Preservation. Int. J. Mol. Sci. 2020, 21, 5471. https://doi.org/10.3390/ijms21155471
Huleihel M, Lunenfeld E. Approaches and Technologies in Male Fertility Preservation. International Journal of Molecular Sciences. 2020; 21(15):5471. https://doi.org/10.3390/ijms21155471
Chicago/Turabian StyleHuleihel, Mahmoud, and Eitan Lunenfeld. 2020. "Approaches and Technologies in Male Fertility Preservation" International Journal of Molecular Sciences 21, no. 15: 5471. https://doi.org/10.3390/ijms21155471
APA StyleHuleihel, M., & Lunenfeld, E. (2020). Approaches and Technologies in Male Fertility Preservation. International Journal of Molecular Sciences, 21(15), 5471. https://doi.org/10.3390/ijms21155471