DoRWA3 from Dendrobium officinale Plays an Essential Role in Acetylation of Polysaccharides

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Isolation and Sequence Analysis of the DoRWA Genes
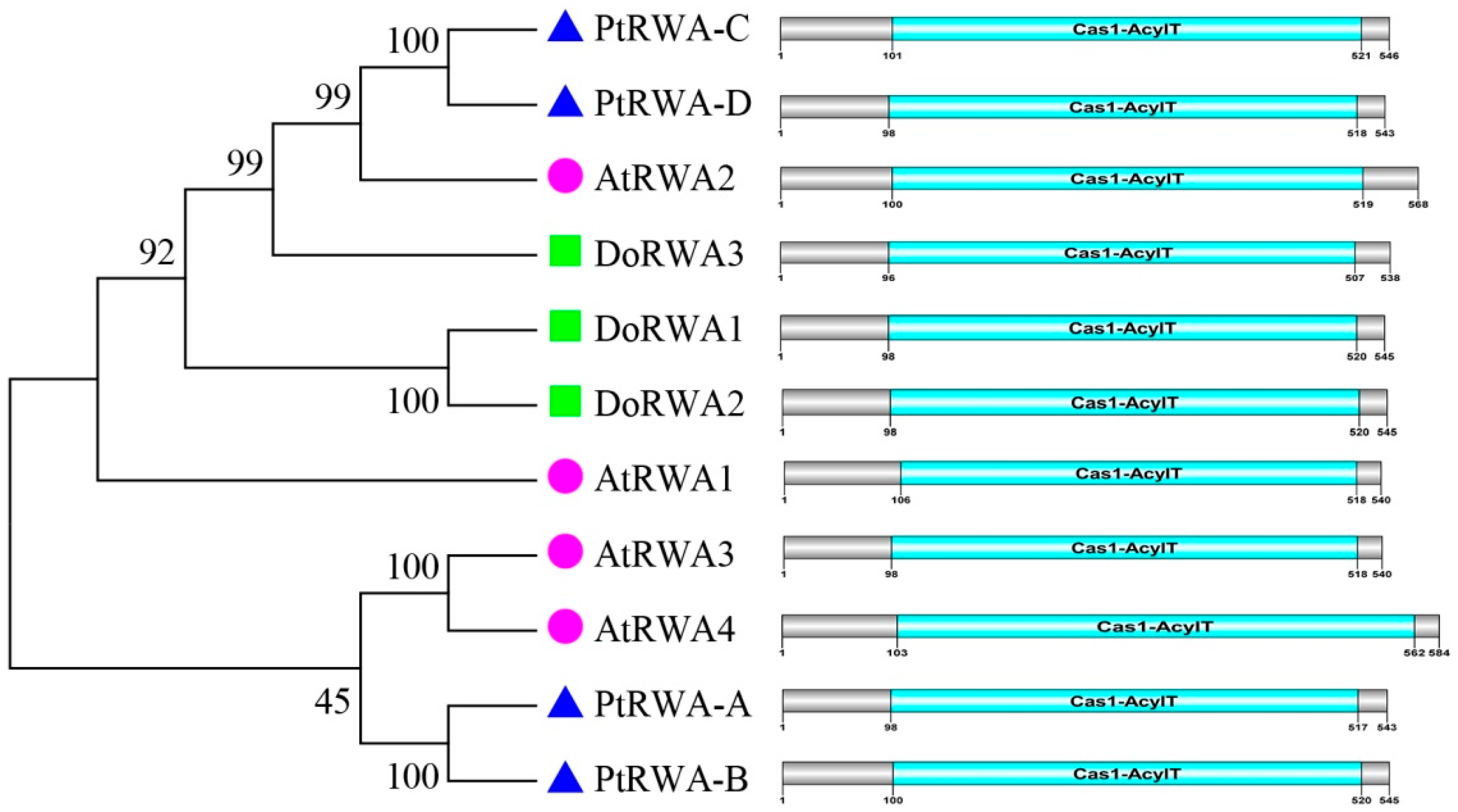
2.2. Bioinformatics of the DoRWA Genes
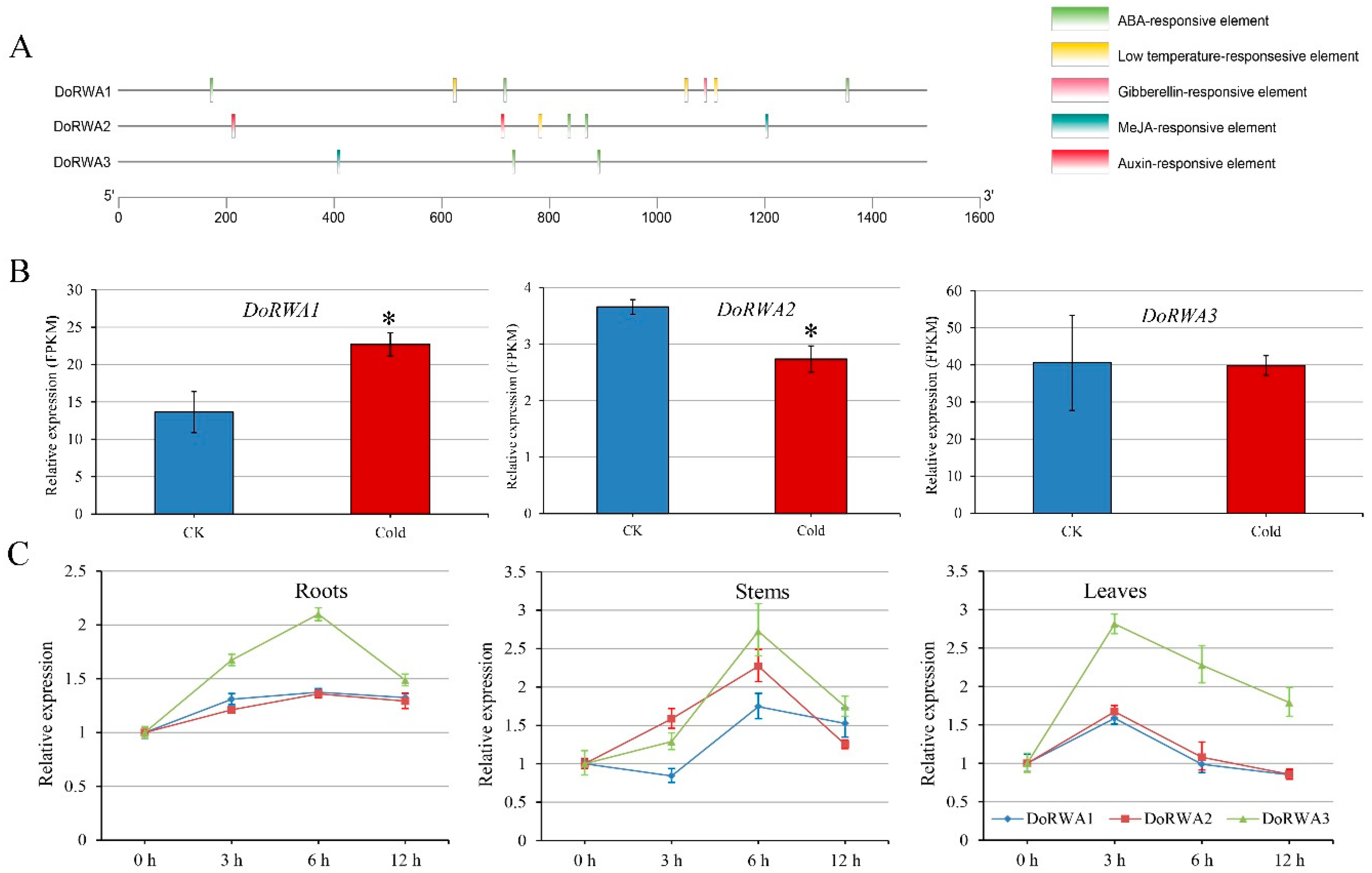
2.3. Analysis of cis-Elements, and Expression Patterns of DoRWA Genes under Cold and ABA Treatments
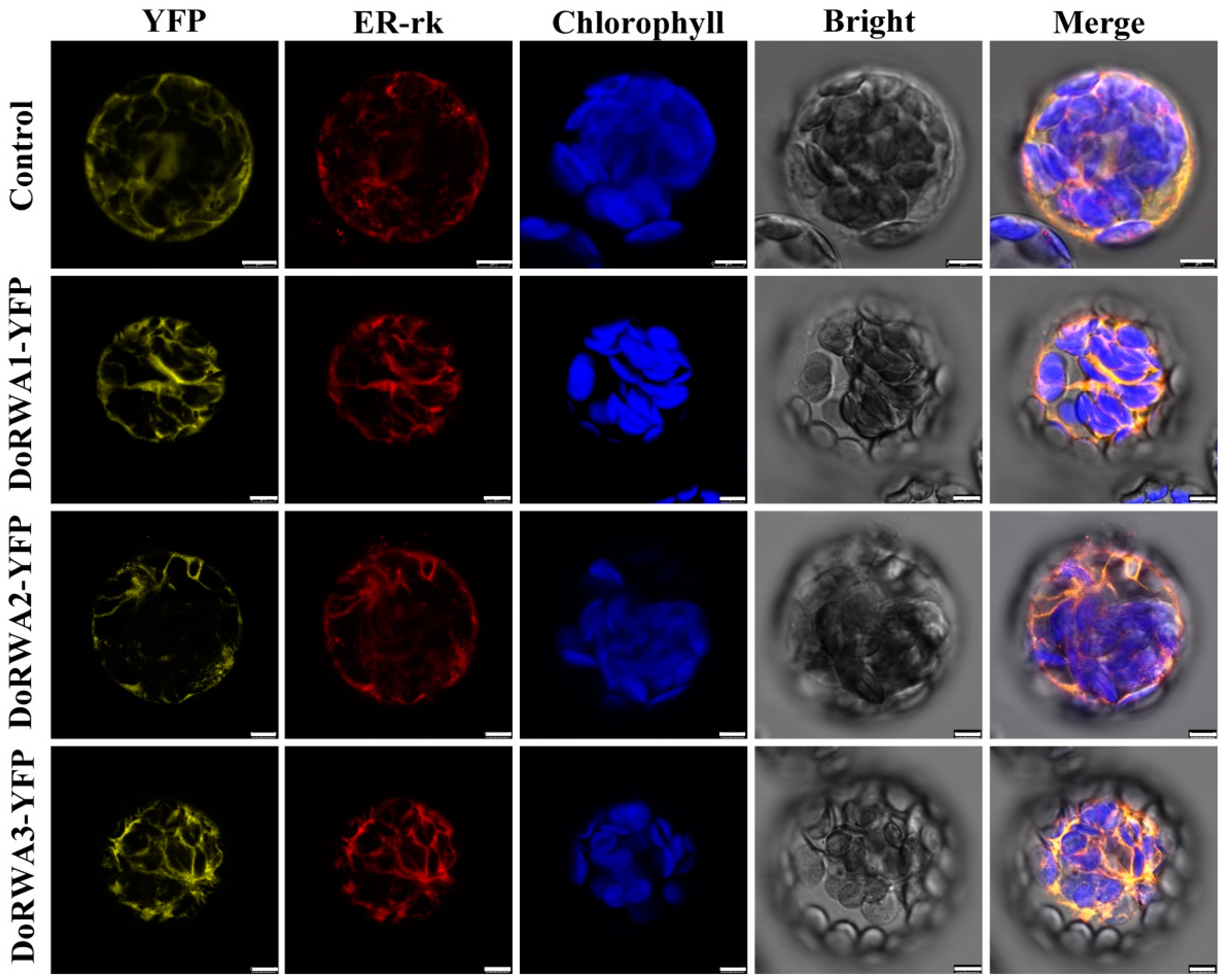
2.4. Cellular Localization of DoRWA Proteins
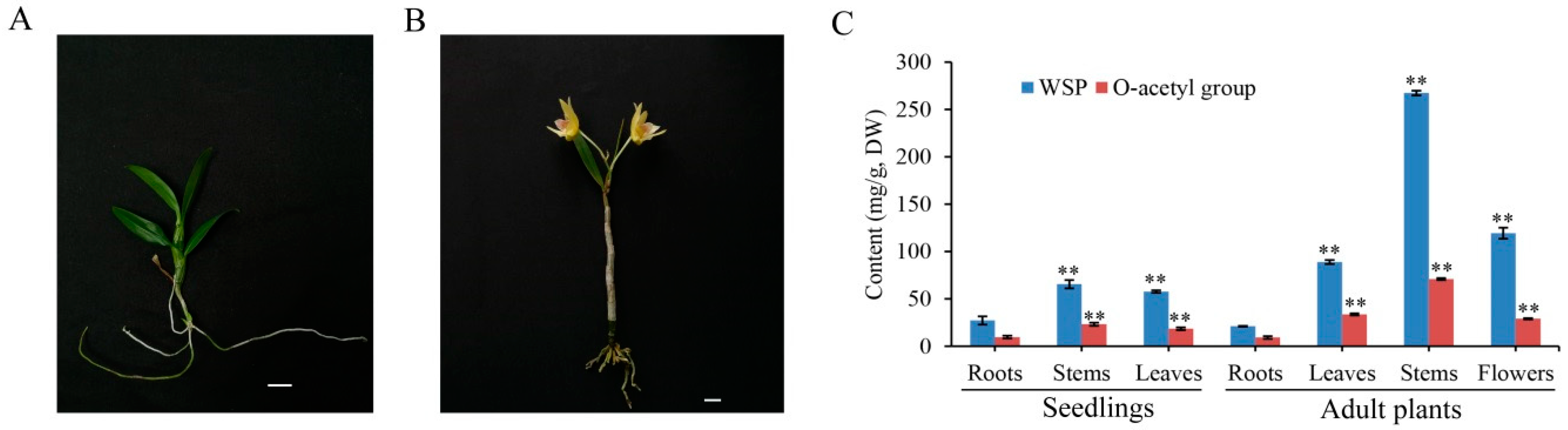
2.5. WSPs and O-Acetyl Groups Mainly Accumulated in the Stems of D. officinale
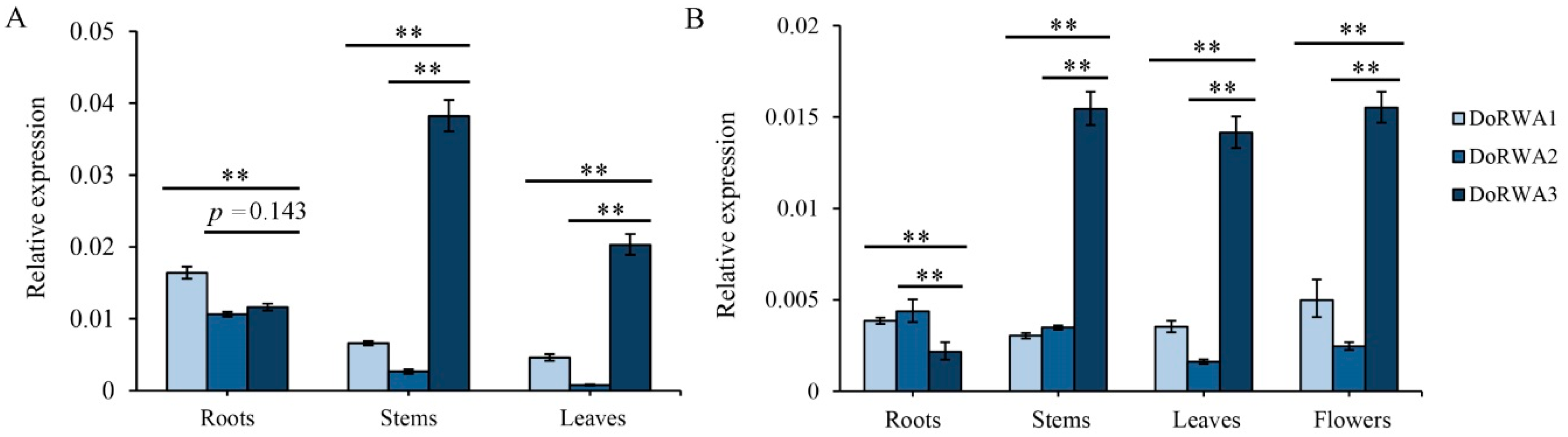
2.6. Expression Patterns of DoRWA Genes in Different Organs of Seedlings and Adult Plants
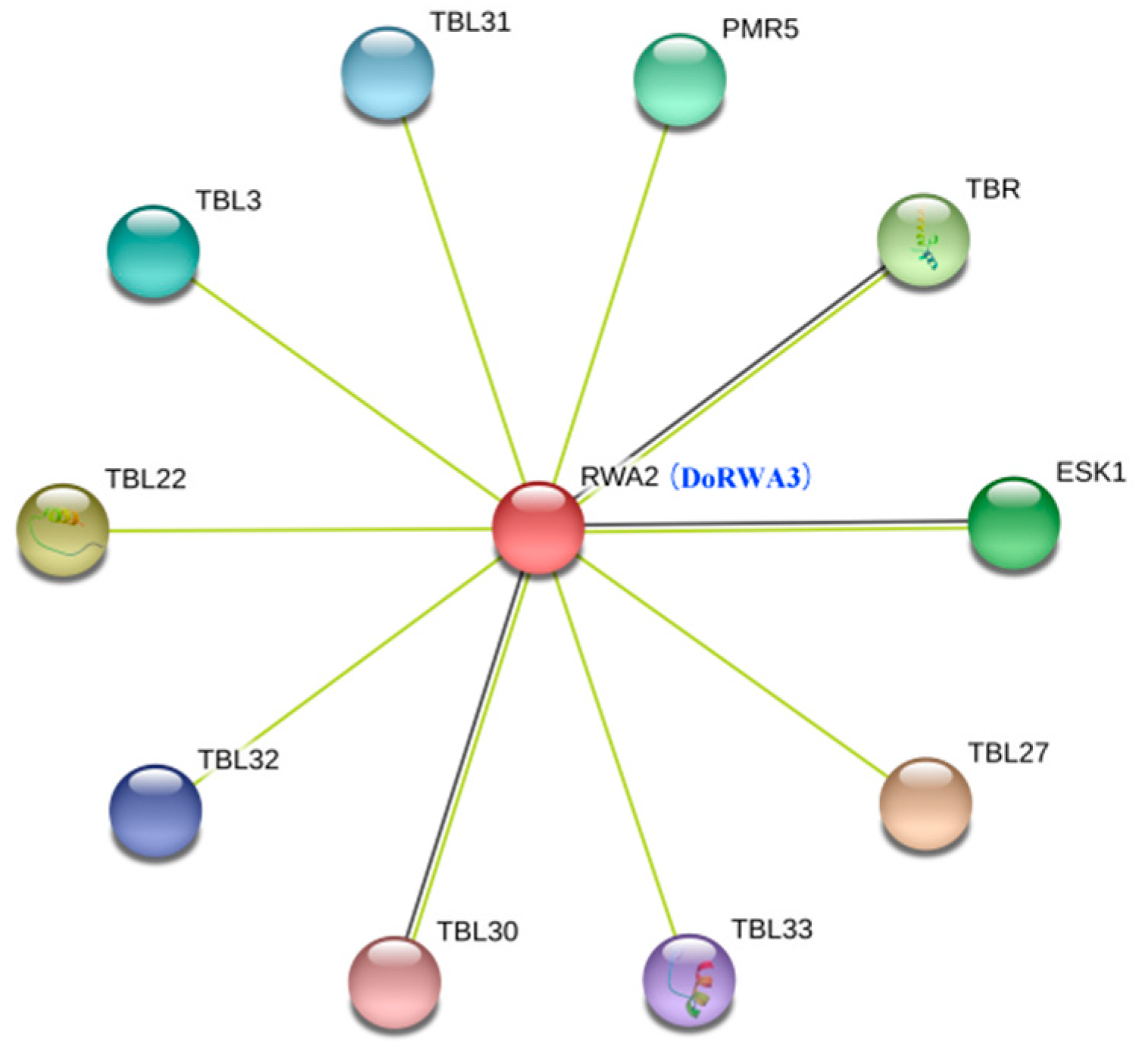
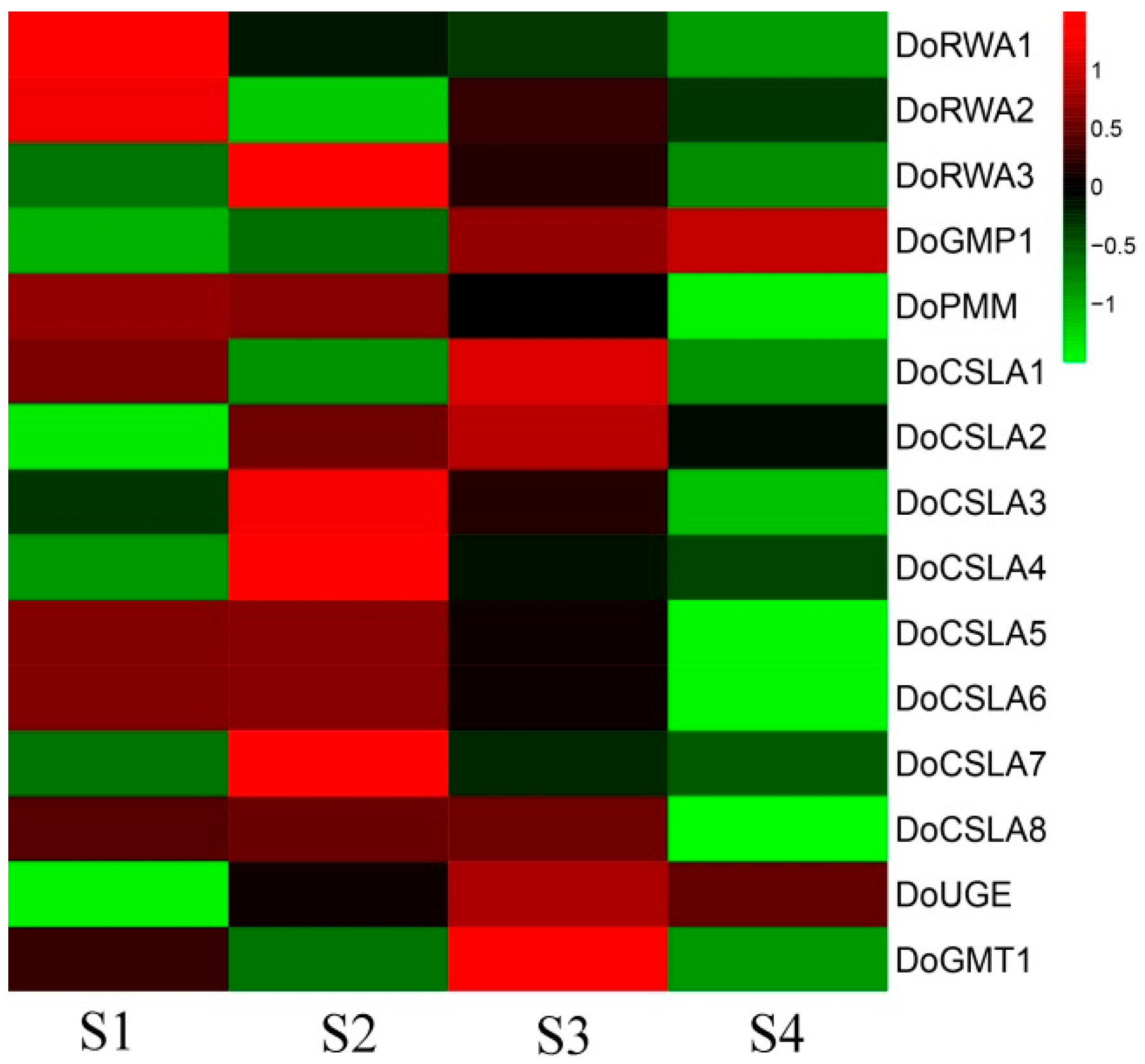
2.7. Co-Expression of DoRWA3 with Synthetic Genes of Polysaccharides
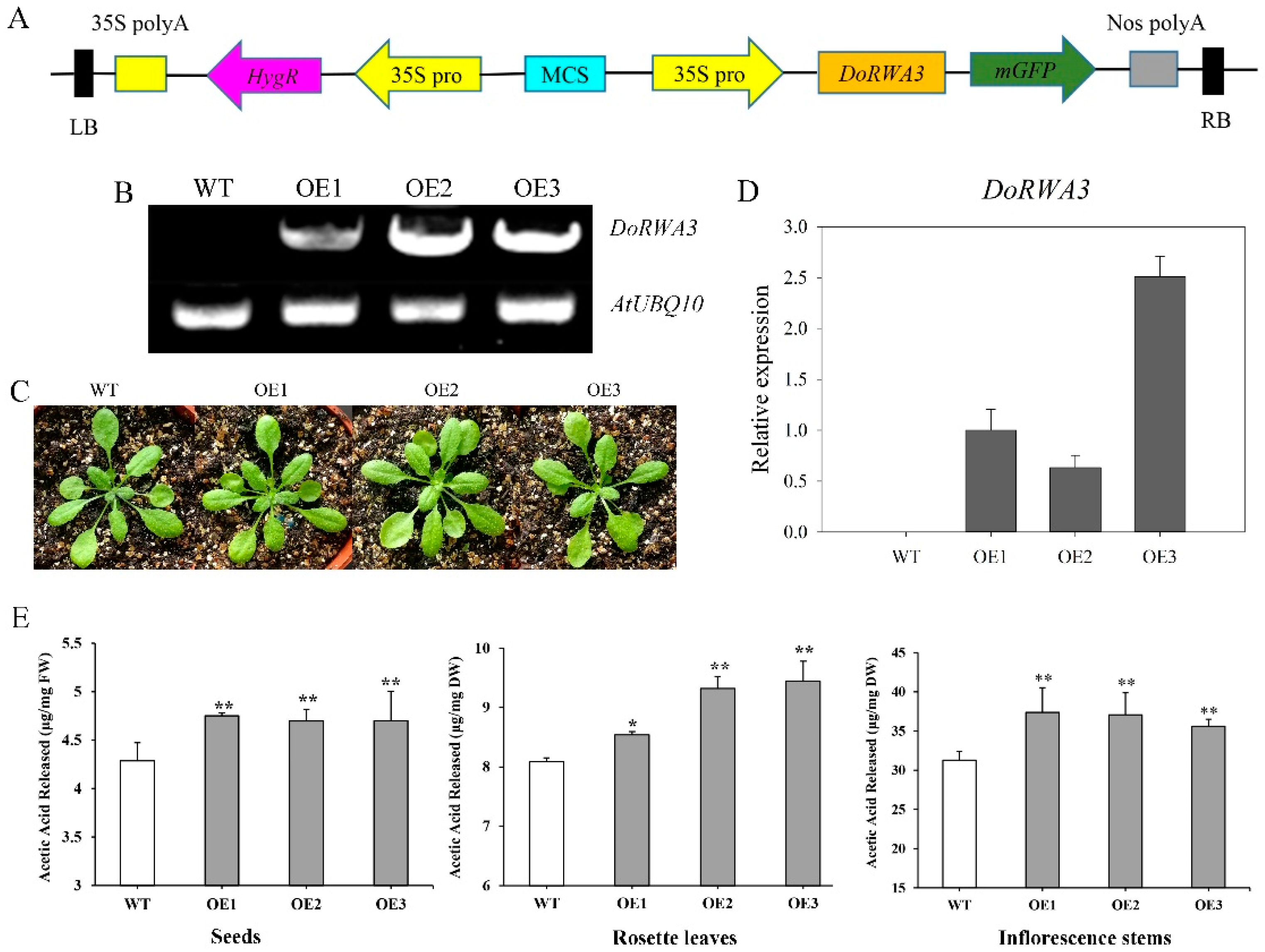
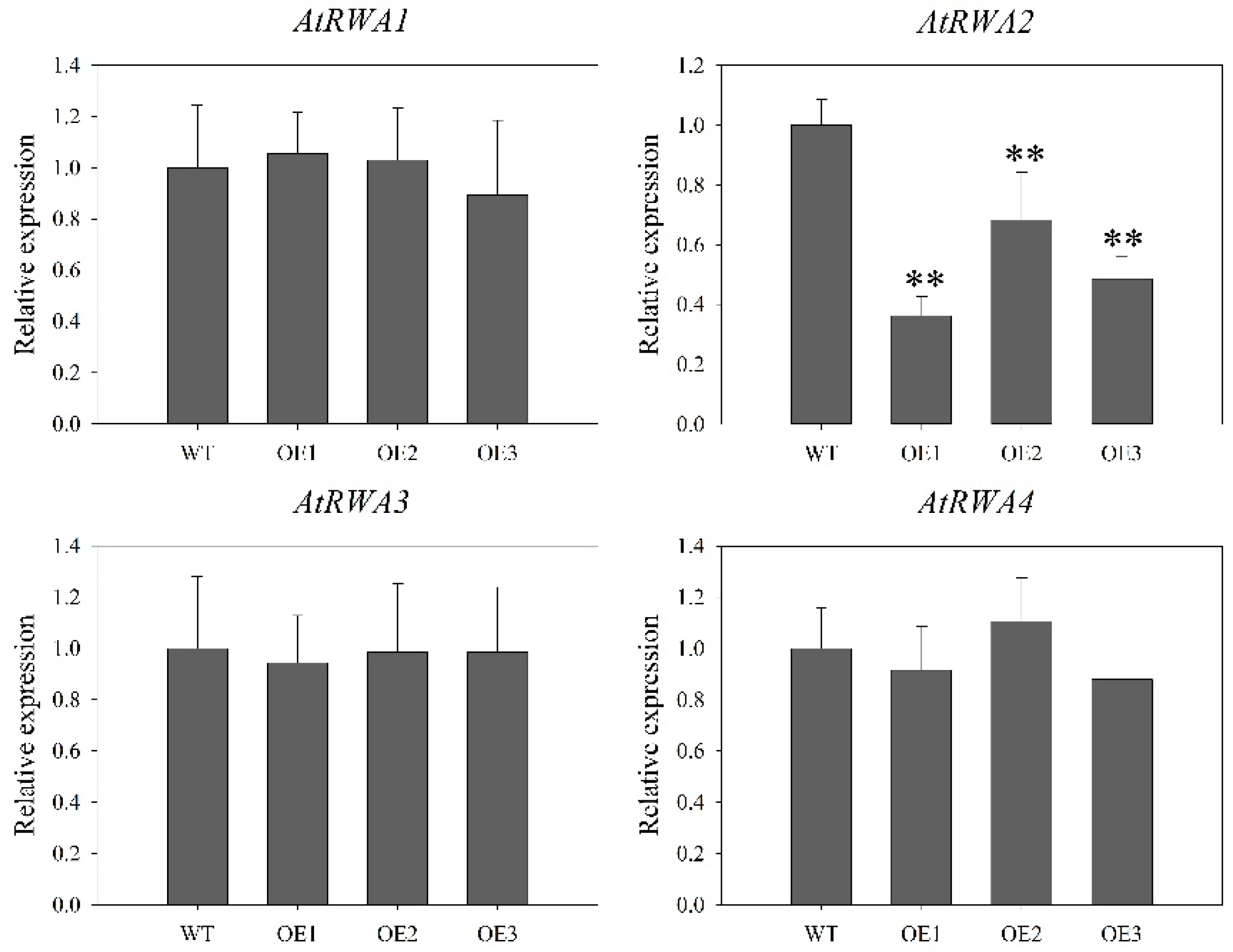
2.8. DoRWA3 Overexpression Increased the Acetylation Level of Polysaccharides in A. thaliana
2.9. ABA Sensitivity Was Not Affected by Constitutive Expression of DoRWA3
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Hormone Treatment
4.2. RNA Extraction, cDNA Synthesis and qRT-PCR Analysis
4.3. Identification and Cloning of DoRWA Genes
4.4. Bioinformatics Analysis
4.5. Protoplast Isolation and Subcellular Localization of DoRWA Proteins
4.6. Content of Water-Soluble Polysaccharides and O-Acetyl Groups in Different Organs
4.7. RNA-Seq Expression Analysis at Four Developmental Stages and under Cold Stress
4.8. Generation of 35S::DoRWA3 Transgenic Lines
4.9. Semi-Quantitative RT-PCR
4.10. Cell Wall Preparation and Determination of Acetyl Esters
4.11. ABA Treatment and Phenotype Assay
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| AIR | Alcohol-insoluble residue |
| Axy9 | Altered xyloglucan9 |
| CSL | Cellulose synthesis-like |
| CK | Cytokinin |
| DA | Degree of acetylation |
| DMRT | Duncan’s multiple range test |
| ER | Endoplasmic reticulum |
| FPKM | Fragments per kilobase per million |
| GMP | GDP-mannose pyrophosphorylation |
| GMT | GDP-mannose transporter |
| GA | Golgi apparatus |
| JA | Jasmonic acid |
| KGM | Konjac glucomannan |
| MW | Molecular weight |
| MS | Murashige and Skoog |
| N-J | Neighbor-Joining |
| ORF | Open reading frame |
| OE | Overexpression |
| PMM | Phosphomannomutase |
| pI | Isoelectric Points |
| RPKM | Reads per kilobase per million |
| RT | Room temperature |
| TCM | Traditional Chinese medicine |
| qRT-PCR | Quantitative real time polymerase chain reaction |
| RWA | Reduced wall acetylation |
| TBL | Trichome birefringence-like |
| UGE | UDP glucose 4-epimerase |
| WSP | Water-soluble polysaccharide |
References
- Zhang, Z.S.; Wang, X.W.; Yu, S.C.; Yin, L.; Zhao, M.X.; Han, Z.P. Synthesized oversulfated and acetylated derivatives of polysaccharide extracted from Enteromorpha linza and their potential antioxidant activity. Int. J. Biol. Macromol. 2011, 49, 1012–1015. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yang, Y.; Zhang, Y.Y.; Duan, L.S.; Zhou, C.L.; Ni, Y.Y.; Liao, X.J.; Li, Q.H.; Hu, X.S. Effect of acetylation on antioxidant and cytoprotective activity of polysaccharides isolated from pumpkin (Cucurbita pepo, lady godiva). Carbohyd. Polym. 2013, 98, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, H.; Wang, Y.X.; Nie, S.P.; Li, C.; Xie, M.Y. Acetylation and carboxymethylation of the polysaccharide from Ganoderma atrum and their antioxidant and immunomodulating activities. Food Chem. 2014, 156, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Huang, G.L. Preparation and immunological activity of polysaccharides and their derivatives. Int. J. Biol. Macromol. 2018, 12, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Chokboribal, J.; Tachaboonyakiat, W.; Sangvanich, P.; Ruangpornvisuti, V.; Jettanacheawchankit, S.; Thunyakitposal, P. Deacetylation affects the physical properties and bioactivity of acemannan, an extracted polysaccharide from Aloe vera. Carbohyd. Polym. 2015, 133, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Du, X.Z.; Li, J.; Chen, J.; Li, B. Effect of degree of deacetylation on physicochemical and gelation properties of konjac glucomannan. Food Res. Int. 2012, 46, 270–278. [Google Scholar] [CrossRef]
- Salah, F.; Ghoul, Y.E.; Mahdhi, A.; Majdoub, H.; Jarroux, N.; Sakli, F. Effect of the deacetylation degree on the antibacterial and antibiofilm activity of acemannan from Aloe vera. Ind. Crop. Prod. 2017, 103, 13–18. [Google Scholar] [CrossRef]
- Huang, S.Y.; Chen, F.; Cheng, H.; Huang, G.L. Modification and application of polysaccharide from traditional Chinese medicine such as Dendrobium officinale. Int. J. Biol. Macromol. 2020, 157, 385–393. [Google Scholar] [CrossRef]
- Lee, C.H.; Teng, Q.H.; Zhong, R.Q.; Ye, Z.H. The four Arabidopsis REDUCED WALL ACETYLATION genes are expressed in secondary wall-containing cells and required for the acetylation of xylan. Plant Cell Physiol. 2011, 52, 1289–1301. [Google Scholar] [CrossRef]
- Manabe, Y.; Nafisi, M.; Verhertbruggen, Y.; Orfila, C.; Gille, S.; Rautengarten, C.; Cherk, C.; Marcus, S.E.; Somerville, S.; Pauly, M.; et al. Loss-of-function mutation of REDUCED WALL ACETYLATION2 in Arabidopsis leads to reduced cell wall acetylation and increased resistance to Botrytis cinerea. Plant Physiol. 2011, 155, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Schultink, A.; Naylor, D.; Dama, M.; Pauly, M. The role of the plant-specific ALTERED XYLOGLUCAN9 protein in Arabidopsis cell wall polysaccharide O-acetylation. Plant Physiol. 2015, 167, 1271–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.; Souza, A.D.; Xiong, G.Y.; Benz, M.; Cheng, K.; Schultink, A.; Reca, I.B.; Pauly, M. O-acetylation of Arabidopsis hemicellulose xyloglucan requires AXY4 or AXY4L, proteins with a TBL and DUF231 Domain. Plant Cell 2011, 23, 4041–4053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, V.; Nita, S.; Neumetzler, L.; Schindelasch, D.; Urbain, A.; Eshed, R.; Persson, S.; Delmer, D.; Scheible, W.R. TRICHOME BIREFRINGENCE and its homolog AT5G01360 encode plant-specific DUF231 proteins required for cellulose biosynthesis in Arabidopsis. Plant Physiol. 2010, 153, 590–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manabe, Y.; Verhertbruggen, Y.; Gill, S.; Harholt, J.; Chong, S.L.; Pawar, P.M.A.; Mellerowicz, E.J.; Tenkanen, M.; Cheng, K.; Pauly, M.; et al. Reduced wall acetylation proteins play vital and distinct roles in cell wall O-acetylation in Arabidopsis. Plant Physiol. 2013, 163, 1107–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawar, P.M.A.; Ratke, C.; Balasubramanian, V.K.; Chong, S.L.; Gandla, M.L.; Adriasola, M.; Sparrman, T.; Hedenström, M.; Szwaj, K.; Derba-Maceluch, M.; et al. Downregulation of RWA genes in hybrid aspen affects xylan acetylation and wood saccharification. New Phytol. 2017, 214, 1491–1505. [Google Scholar] [CrossRef] [Green Version]
- Teixeira da Silva, J.A.; Ng, T.B. The medicinal and pharmaceutical importance of Dendrobium species. Appl. Microbiol. Biotechnol. 2017, 101, 2227–2239. [Google Scholar] [CrossRef]
- Xing, X.H.; Cui, S.W.; Nie, S.P.; Phillips, G.O. Study on Dendrobium officinale O-acetyl-glucomannan (Dendronan®): Part II. Fine structure of O-acetylated residues. Carbohyd. Polym. 2015, 117, 422–433. [Google Scholar] [CrossRef]
- Huang, X.J.; Nie, S.P.; Cai, H.L.; Zhang, G.Y.; Cui, S.W.; Xie, M.Y.; Phillips, G.O. Study on Dendrobium officinale O-acetyl-glucomannan (Dendronan®): Part IV. Immunomogulatory activity in vivo. J. Funct. Foods 2015, 15, 525–532. [Google Scholar] [CrossRef]
- Wei, W.; Feng, L.; Bao, W.R.; Ma, D.L.; Leung, C.H.; Nie, S.P.; Han, Q.B. (2016) Structure characterization and immunomodulating effects of polysaccharides isolated from Dendrobium officinale. J. Agric. Food Chem. 2016, 64, 881–889. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Xu, Q.; Bian, C.; Tsai, W.C.; Yeh, C.M.; Liu, K.W.; Youshida, K.; Zhang, L.S.; Chang, S.B.; Chen, F.; et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 19029. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, S.A.; Larue, C.; Bonk, L.; Khan, A.; Michel, H.C.; Stein, R.J.; Grolimund, D.; Begerow, D.; Neumann, U.; Haydon, M.J.; et al. Etiolated seedlings development requires repression of photomorphogenesis by a small cell-wall-derived dark signal. Curr. Biol. 2017, 27, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.X.; Teng, Q.; Zhong, R.Q.; Ye, Z.H. TBL3 and TBL31, two Arabidopsis DUF231 domain proteins, are required for 3-O-monoacetylation of xylan. Plant Cell Physiol. 2016, 57, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.F.; Sun, Y.; Zhang, B.C.; Mansoori, N.; Wan, J.X.; Liu, Y.; Wang, Z.W.; Shi, Y.Z.; Zhou, Y.H.; Zheng, S.J. TRICHOME BIREFRINGENCE-LIKE27 affects aluminum sensitivity by modulating the O-acetylation of xyloglucan and aluminum-binding capacity in Arabidopsis. Plant Physiol. 2014, 166, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, B.K.; Cai, X.; Nebenführ, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef]
- He, C.M.; Zhang, J.X.; Liu, X.C.; Zeng, S.J.; Wu, K.L.; Yu, Z.M.; Wang, X.J.; Teixeira da Silva, J.A.; Lin, Z.J.; Duan, J. Identification of genes involved in biosynthesis of mannan polysaccharides in Dendrobium officinale by RNA-seq analysis. Plant Mol. Biol. 2015, 88, 219–231. [Google Scholar] [CrossRef]
- He, C.M.; Yu, Z.M.; Teixeira da Silva, J.A.; Zhang, J.X.; Liu, X.C.; Wang, X.J.; Zhang, X.H.; Zeng, S.J.; Wu, K.L.; Tan, J.W.; et al. DoGMP1 from Dendrobium officinale contributes to mannose content of water-soluble polysaccharides and plays a role in salt stress response. Sci. Rep. 2017, 7, 41040. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.M.; He, C.M.; Teixeira da Silva, J.A.; Zhang, G.H.; Dong, W.; Luo, J.P.; Duan, J. Molecular cloning and functional analysis of DoUGE related to water-soluble polysaccharides from Dendrobium officinale with enhanced abiotic stress tolerance. Plant Cell Tissue Organ Cult. 2017, 131, 579–599. [Google Scholar] [CrossRef]
- Yu, Z.M.; He, C.M.; Teixeira da Silva, J.A.; Luo, J.P.; Yang, Z.Y.; Duan, J. The GDP-mannose transporter gene (DoGMT) from Dendrobium officinale is critical for mannan biosynthesis in plant growth and development. Plant Sci. 2018, 277, 43–54. [Google Scholar] [CrossRef]
- Liu, X.; Xie, J.H.; Jia, S.; Huang, L.X.; Wang, Z.J.; Li, C.; Xie, M.Y. Immunomodulatory effects of an acetylated Cyclocarya paliurus polysaccharide on murine macrophages RAW264.7. Int. J. Biol. Macromol. 2017, 98, 576–581. [Google Scholar] [CrossRef]
- Hsieh, Y.Y.; Chien, C.; Liao, S.K.S.; Liao, S.F.; Huang, W.T.; Yang, W.B.; Lin, C.C.; Cheng, T.J.R.; Chang, C.C.; Fang, J.M.; et al. Structure and bioactivity of polysaccharides in medicinal plant Dendrobium huoshanense. Bioorgan. Med. Chem. 2008, 16, 6054–6068. [Google Scholar] [CrossRef]
- Tong, W.; Yu, Q.; Li, H.; Cui, W.W.; Nie, S.P. Chemical modification and immunoregulatory activity of polysaccharides from Dendrobium officinale. Food Sci. 2017, 38, 155–160. [Google Scholar]
- Jiang, L.M.; Nie, S.P.; Huang, D.F.; Fu, Z.H.; Xie, M.Y. Acetylation modification improves immunoregulatory effect of polysaccharide from seeds of Plantago asiatica L. J. Chem. 2018, 2018, 3082026. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ye, T.; Wu, X.F.; Chen, J.; Wang, S.S.; Lin, L.F.; Li, B. Preparation and characterization of heterogeneous deacetylated konjac glucomannan. Food Hydrocoll. 2014, 40, 9–15. [Google Scholar] [CrossRef]
- Javed, S.; Rahman, A. Aloe vera gel in food, health products and cosmetics industry. In Studies in Natural Products and Chemistry; Rahman, A.U., Ed.; Elsevier Science & Technology: London, UK, 2014; Volume 41, pp. 261–285. [Google Scholar]
- Nishinari, K.; Williams, P.A.; Phillips, G.O. Review of the physicochemical characteristics and properties of konjac mannan. Food Hydrocoll. 1992, 6, 199–222. [Google Scholar] [CrossRef]
- Li, M.Y.; Feng, G.P.; Wang, H.; Yang, R.L.; Xu, Z.L.; Sun, Y.M. Deacetylated konjac glucomannan is less effective in reducing dietary-induced hyperlipidemia and hepatic steatosis in C57BL/6 mice. J. Agric. Food Chem. 2017, 65, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Humman, J.H. Composition and applications of Aloe vera leaf gel. Molecules 2008, 13, 1599–1616. [Google Scholar] [CrossRef] [Green Version]
- Nafisi, M.; Stranne, M.; Fimognari, L.; Atwell, S.; Martens, H.J.; Pedas, P.R.; Hansen, S.F.; Nawrath, C.; Scheller, H.V.; Kliebenstein, D.J.; et al. Acetylation of cell wall is required for structural integrity of the leaf surface and exerts a global impact on plant stress responses. Front. Plant Sci. 2015, 6, 550. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Chattopadhyay, N.; Kher, R.; Godbole, M. Inexpensive SDS/phenol method for RNA extraction from tissues. Biotechniques 1993, 15, 24–26. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, M.M.; Zhang, D.W.; Guo, S.X. Reference gene selection for real-time quantitative PCR analysis of Dendrobium officinale. Chin. Pharm. J. 2013, 48, 1664–1668. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.H.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Wen, L.P.; Gao, X.J.; Jin, C.J.; Xue, Y.; Yao, X.B. DOG1.0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.V.D.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucl. Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools—An integrative toolkit developed for interactive of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Cepas, J.H.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucl. Acids Res. 2019, 47, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Schapire, A.L.; Lois, L.M. A simplified and rapid method for the isolation and transfection of Arabidopsis leaf mesophyll protoplasts for large-scale applications. In Methods in Molecular Biology; Walker, J.M., Clifton, N.J., Eds.; Humana Press: New York, NY, USA, 2016; pp. 79–88. [Google Scholar]
- Citovsky, V.; Lee, L.Y.; Vyas, S.; Glick, E.; Chen, M.E.; Vainstein, A.; Gafni, Y.; Gelvin, S.B.; Tzfira, T. Subcellular localization of interacting proteins by biomolecular fluorescence complementation in planta. J. Mol. Biol. 2006, 362, 1120–1131. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Gudlavalleti, S.K.; Datta, A.K.; Tzeng, Y.L.; Noble, C.; Carlson, R.W.; Stephens, D.S. The Neisseria meningitides serogroup a capsular polysaccharide O-3 and O-4 acetyltransferase. J. Biol. Chem. 2004, 279, 42765–42773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.X.; He, C.M.; Wu, K.L.; Teixeira da Silva, J.A.; Zeng, S.J.; Zhang, X.H.; Yu, Z.M.; Xia, H.Q.; Duan, J. Transcriptome analysis of Dendrobium officinale and its application to the identification of genes associated with polysaccharide synthesis. Front. Plant. Sci. 2016, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.G.; Jiang, W.; Chen, S.L.; Mantri, N.; Tao, Z.M.; Jiang, C.X. Insights from the cold transcriptome and metabolome of Dendrobium officinale: Global reprogramming of metabolic and gene regulation networks during cold accumulation. Front. Plant Sci. 2016, 7, 1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation for Arabidopsis thaliana. Plant. J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harholt, J.; Jensen, J.K.; Sørensen, S.O.; Orfila, C.; Pauly, M.; Scheller, H.V. ARABINAN DEFICIENT 1 is a putative arabinosyltransferase involved in biosynthesis of pectic arabinan in Arabidopsis. Plant Physiol. 2006, 140, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.; Cheng, K.; Skinner, M.E.; Liepman, A.H.; Wilkerson, C.G.; Pauly, M. Deep sequencing of voodoo lily (Amorphophallus konjac): An approach to identify relevant genes involved in the synthesis of the hemicellulose glucomannan. Planta 2011, 234, 515–526. [Google Scholar] [CrossRef] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Si, C.; Teixeira da Silva, J.A.; He, C.; Yu, Z.; Zhao, C.; Wang, H.; Zhang, M.; Duan, J. DoRWA3 from Dendrobium officinale Plays an Essential Role in Acetylation of Polysaccharides. Int. J. Mol. Sci. 2020, 21, 6250. https://doi.org/10.3390/ijms21176250
Si C, Teixeira da Silva JA, He C, Yu Z, Zhao C, Wang H, Zhang M, Duan J. DoRWA3 from Dendrobium officinale Plays an Essential Role in Acetylation of Polysaccharides. International Journal of Molecular Sciences. 2020; 21(17):6250. https://doi.org/10.3390/ijms21176250
Chicago/Turabian StyleSi, Can, Jaime A. Teixeira da Silva, Chunmei He, Zhenming Yu, Conghui Zhao, Haobin Wang, Mingze Zhang, and Jun Duan. 2020. "DoRWA3 from Dendrobium officinale Plays an Essential Role in Acetylation of Polysaccharides" International Journal of Molecular Sciences 21, no. 17: 6250. https://doi.org/10.3390/ijms21176250
APA StyleSi, C., Teixeira da Silva, J. A., He, C., Yu, Z., Zhao, C., Wang, H., Zhang, M., & Duan, J. (2020). DoRWA3 from Dendrobium officinale Plays an Essential Role in Acetylation of Polysaccharides. International Journal of Molecular Sciences, 21(17), 6250. https://doi.org/10.3390/ijms21176250