Fusion Potential of Human Osteoclasts In Vitro Reflects Age, Menopause, and In Vivo Bone Resorption Levels of Their Donors—A Possible Involvement of DC-STAMP


 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Bone Biomarkers, CTX, and PINP Increase with Age
2.2. OCs Generated In Vitro Gain More Nuclei as Donor Age Increases
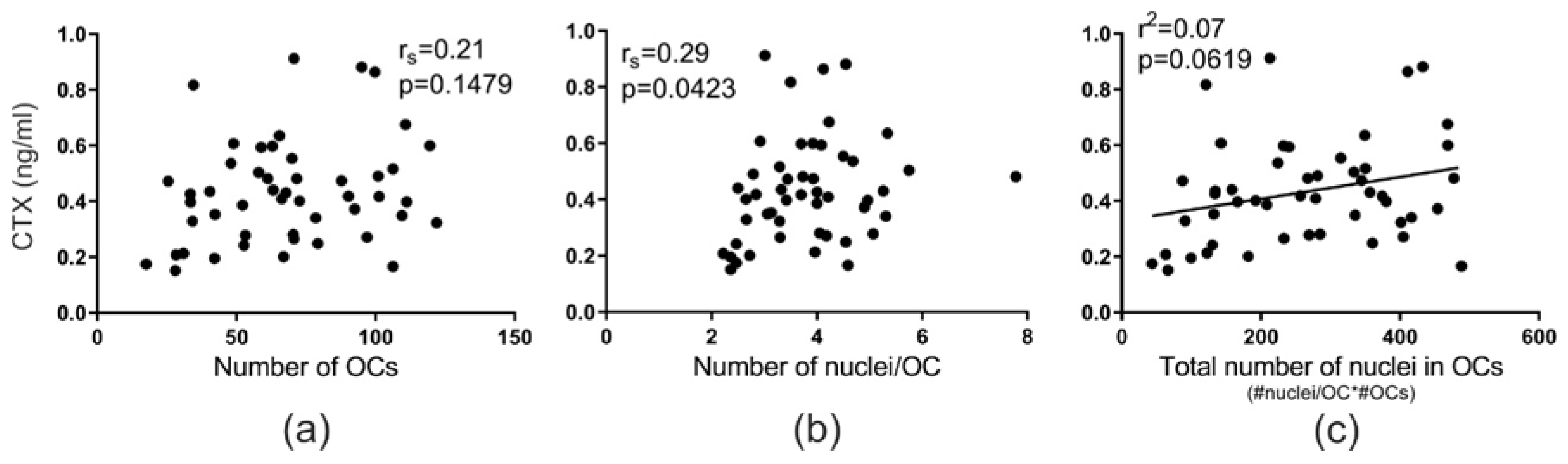
2.3. Fusion Potential of OCs In Vitro Reflects In Vivo Bone Resorption Levels of Their Donor
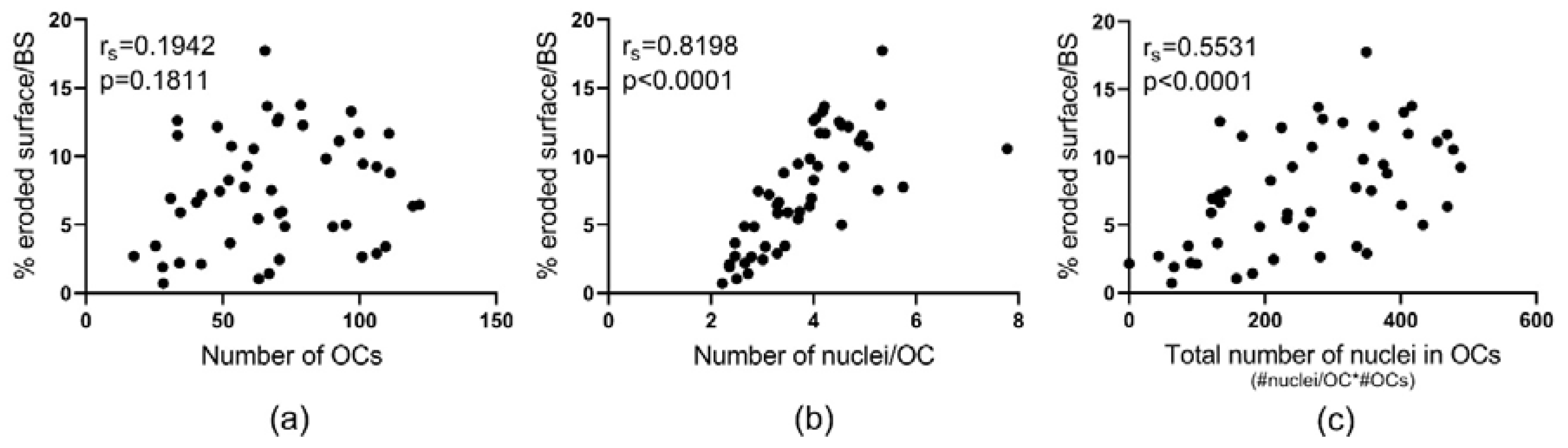
2.4. Fusion Potential of OCs Determines Their Bone Resorptive Activity In Vitro
2.5. DCSTAMP Expression Parallels Age, CTX Levels In Vivo, and Number of Nuclei Per OC In Vitro
2.6. DNA Methylation Levels of Individual Sites in the DCSTAMP Gene Promoter Correlate with Age, CTX In Vivo, and the Number of OCs In Vitro
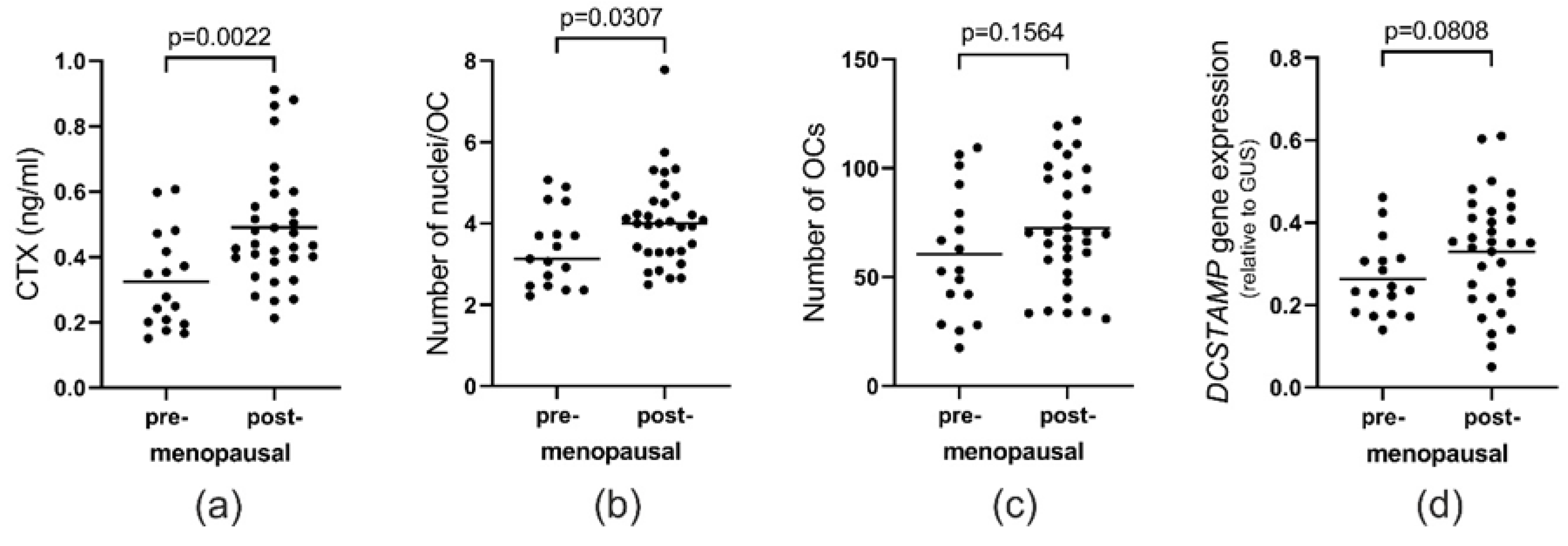
2.7. Menopause Status Also Has an Effect on the Fusion Potential of OCs
3. Discussion
4. Materials and Methods
4.1. Participants, Sample Collection, and Demographics
4.2. CTX-I and PINP Measurements
4.3. In Vitro Generation of Human OCs
4.4. Bone Resorption Assays and Quantification of Bone Resorption
4.5. Droplet Digital RT-PCR
4.6. DNA Methylation Analyses Using Pyrosequencing
4.7. Statistics
4.8. Study Approval
4.9. Data Availability
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CTX | C-terminal telopeptide of type I collagen |
| DC-STAMP | Dendritic cell-specific transmembrane protein |
| DCSTAMP | Gene encoding “Dendritic cell-specific transmembrane protein” |
| M-CSF | Macrophage colony-stimulating factor |
| OC | Osteoclast |
| PINP | Procollagen type I N-terminal propeptide |
| RANKL | Receptor activator of nuclear factor kappa-B ligand |
References
- Parfitt, A.M.; Mathews, C.H.E.; Villanueva, A.B.; Kleerekoper, M.; Frame, B.; Rao, D.S. Relationships between surface, volume, and thickness of iliac trabecular bone in aging and in osteoporosis. Implications for the microanatomic and cellular mechanisms of bone loss. J. Clin. Investig. 1983, 72, 1396–1409. [Google Scholar] [CrossRef] [PubMed]
- Riggs, B.L.; Khosla, S.; Melton, L.J. Sex steroids and the construction and conservation of the adult skeleton. Endocr. Rev. 2002, 23, 279–302. [Google Scholar] [CrossRef] [PubMed]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Katsimbri, P. The biology of normal bone remodelling. Eur. J. Cancer Care 2017, 26, e12740. [Google Scholar] [CrossRef]
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporos. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Eastell, R.; Hannon, R.A. Biomarkers of bone health and osteoporosis risk. Proc. Nutr. Soc. 2008, 67, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Garnero, P.; Sornay-Rendu, E.; Chapuy, M.-C.; Delmas, P.D. Increased bone turnover in late postmenopausal women is a major determinant of osteoporosis. J. Bone Miner. Res. 1996, 11, 337–349. [Google Scholar] [CrossRef]
- Ravn, P.; Fledelius, C.; Rosenquist, C.; Overgaard, K.; Christiansen, C. High bone turnover is associated with low bone mass in both pre- and postmenopausal women. Bone 1996, 19, 291–298. [Google Scholar] [CrossRef]
- Mouritzen, U.; Christgau, S.; Lehmann, H.J.; Tankó, L.B.; Christiansen, C. Cartilage turnover assessed with a newly developed assay measuring collagen type II degradation products: Influence of age, sex, menopause, hormone replacement therapy, and body mass index. Ann. Rheum. Dis. 2003, 62, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Glover, S.J.; Gall, M.; Schoenborn-Kellenberger, O.; Wagener, M.; Garnero, P.; Boonen, S.; Cauley, J.A.; Black, D.M.; Delmas, P.D.; Eastell, R. Establishing a reference interval for bone turnover markers in 637 healthy, young, premenopausal women from the united kingdom, france, belgium, and the united states. J. Bone Miner. Res. 2009, 24, 389–397. [Google Scholar] [CrossRef]
- Michelsen, J.; Wallaschofski, H.; Friedrich, N.; Spielhagen, C.; Rettig, R.; Ittermann, T.; Nauck, M.; Hannemann, A. Reference intervals for serum concentrations of three bone turnover markers for men and women. Bone 2013, 57, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Ardawi, M.S.M.; Maimani, A.A.; Bahksh, T.A.; Rouzi, A.A.; Qari, M.H.; Raddadi, R.M. Reference intervals of biochemical bone turnover markers for Saudi Arabian women: A cross-sectional study. Bone 2010, 47, 804–814. [Google Scholar] [CrossRef]
- Jenkins, N.; Black, M.; Paul, E.; Pasco, J.A.; Kotowicz, M.A.; Schneider, H.G. Age-related reference intervals for bone turnover markers from an Australian reference population. Bone 2013, 55, 271–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Li, Y.; Deng, W.; Zhang, Z.; Deng, Z.; Hu, Y.; Xia, W.; Xu, L. Chinese Bone Turnover Marker Study: Reference ranges for C-terminal telopeptide of type i collagen and procollagen I N-terminal peptide by age and gender. PLoS ONE 2014, 9, e103841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jørgensen, N.R.; Møllehave, L.T.; Hansen, Y.B.L.; Quardon, N.; Lylloff, L.; Linneberg, A. Comparison of two automated assays of BTM (CTX and P1NP) and reference intervals in a Danish population. Osteoporos. Int. 2017, 28, 2103–2113. [Google Scholar] [CrossRef] [PubMed]
- Kushida, K.; Takahashi, M.; Kawana, K.; Inoue, T. Comparison of markers for bone formation and resorption in premenopausal and postmenopausal subjects, and osteoporosis patients. J. Clin. Endocrinol. Metab. 1995, 80, 2447–2450. [Google Scholar]
- Møller, A.M.J.; Delaissé, J.-M.; Olesen, J.B.; Madsen, J.S.; Canto, L.M.; Bechmann, T.; Rogatto, S.R.; Søe, K. Aging and menopause reprogram osteoclast precursors for aggressive bone resorption. Bone Res. 2020, 8, 27. [Google Scholar] [CrossRef]
- Chin, K.Y. The relationship between follicle-stimulating hormone and bone health: Alternative explanation for bone loss beyond oestrogen? Int. J. Med. Sci. 2018, 15, 1373–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randolph, J.F.; Zheng, H.; Sowers, M.F.R.; Crandall, C.; Crawford, S.; Gold, E.B.; Vuga, M. Change in follicle-stimulating hormone and estradiol across the menopausal transition: Effect of age at the final menstrual period. J. Clin. Endocrinol. Metab. 2011, 96, 746–754. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Yu, C.; Zhang, X.; Zhang, H.; Guan, Q.; Zhao, J.; Xu, J. Follicle-stimulating hormone increases the risk of postmenopausal osteoporosis by stimulating osteoclast differentiation. PLoS ONE 2015, 10, e0134986. [Google Scholar] [CrossRef] [Green Version]
- Riggs, B.L. The mechanisms of estrogen regulation of bone resorption. J. Clin. Investig. 2000, 106, 1203–1204. [Google Scholar] [CrossRef] [Green Version]
- Khosla, S.; Oursler, M.J.; Monroe, D.G. Estrogen and the skeleton. Trends Endocrinol. Metab. 2012, 23, 576–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, M. New Insights into the Molecular Mechanisms of Action of Bisphosphonates. Curr. Pharm. Des. 2005, 9, 2643–2658. [Google Scholar] [CrossRef] [PubMed]
- Xing, L. Osteoclast fusion and regulation by RANKL-dependent and independent factors. World J. Orthop. 2012, 3, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Yagi, M.; Miyamoto, T.; Sawatani, Y.; Iwamoto, K.; Hosogane, N.; Fujita, N.; Morita, K.; Ninomiya, K.; Suzuki, T.; Miyamoto, K.; et al. DC-STAMP is essential for cell-cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 2005, 202, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, M.; Miyamoto, T.; Toyama, Y.; Suda, T. Role of DC-STAMP in cellular fusion of osteoclasts and macrophage giant cells. J. Bone Miner. Metab. 2006, 24, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Søe, K.; Delaisse, J.-M.; Borggaard, X.G. Osteoclast Formation at the Bone Marrow/Bone Surface Interface: Importance of Structural Elements, Matrix, and Intercellular Communication. Semin Cell Dev Biol. 2020. [Google Scholar] [CrossRef]
- Wiktor-Jedrzejczak, W.; Bartocci, A.; Ferrante, A.W.; Ahmed-Ansari, A.; Sell, K.W.; Pollard, J.W.; Stanley, E.R. Total absence of colony-stimulating factor 1 in the macrophage-deficient osteopetrotic (op/op) mouse. Proc. Natl. Acad. Sci. USA 1990, 87, 4828–4832. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Teitelbaum, S.L. Osteoclasts: New Insights. Bone Res. 2013, 1, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.I.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A.; et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef] [Green Version]
- Wong, B.R.; Rho, J.; Arron, J.; Robinson, E.; Orlinick, J.; Chao, M.; Kalachikov, S.; Cayani, E.; Bartlett, F.S.; Frankel, W.N.; et al. TRANCE is a novel ligand of the tumor necrosis factor receptor family that activates c-Jun N-terminal kinase in T cells. J. Biol. Chem. 1997, 272, 25190–25194. [Google Scholar] [CrossRef] [Green Version]
- Mensah, K.A.; Ritchlin, C.T.; Schwarz, E.M. RANKL induces heterogeneous DC-STAMPlo and DC-STAMPhi osteoclast precursors of which the DC-STAMPlo precursors are the master fusogens. J. Cell. Physiol. 2010, 223, 76–83. [Google Scholar]
- Chiu, Y.H.; Mensah, K.A.; Schwarz, E.M.; Ju, Y.; Takahata, M.; Feng, C.; McMahon, L.A.; Hicks, D.G.; Panepento, B.; Keng, P.C.; et al. Regulation of human osteoclast development by dendritic cell-specific transmembrane protein (DC-STAMP). J. Bone Miner. Res. 2012, 27, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukita, T.; Wada, N.; Kukita, A.; Kakimoto, T.; Sandra, F.; Toh, K.; Nagata, K.; Iijima, T.; Horiuchi, M.; Matsusaki, H.; et al. RANKL-induced DC-STAMP is essential for osteoclastogenesis. J. Exp. Med. 2004, 200, 941–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Dou, C.; Xu, J.; Dong, S. DC-STAMP, the key fusion-mediating molecule in osteoclastogenesis. J. Cell. Physiol. 2014, 229, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.M.J.; Delaissé, J.M.; Søe, K. Osteoclast Fusion: Time-Lapse Reveals Involvement of CD47 and Syncytin-1 at Different Stages of Nuclearity. J. Cell. Physiol. 2017, 232, 1396–1403. [Google Scholar] [CrossRef] [PubMed]
- Hobolt-Pedersen, A.S.; Delaissé, J.M.; Søe, K. Osteoclast fusion is based on heterogeneity between fusion partners. Calcif. Tissue Int. 2014, 95, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Søe, K.; Hobolt-Pedersen, A.S.; Delaisse, J.M. The elementary fusion modalities of osteoclasts. Bone 2015, 73, 181–189. [Google Scholar] [CrossRef]
- Wood, C.L.; Stenson, C.; Embleton, N. The Developmental Origins of Osteoporosis. Curr. Genom. 2015, 16, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Baird, J.; Kurshid, M.A.; Kim, M.; Harvey, N.; Dennison, E.; Cooper, C. Does birthweight predict bone mass in adulthood? A systematic review and meta-analysis. Osteoporos. Int. 2011, 22, 1323–1334. [Google Scholar] [CrossRef]
- Mundy, G.R. The morphology of osteoclast. In Bone Remodeling and Its Disorders; Martin Dunitz: London, UK, 1995; pp. 12–13. [Google Scholar]
- Roodman, G.D. Paget’s disease and osteoclast biology. Bone 1996, 19, 209–212. [Google Scholar] [CrossRef]
- Addison, W.C. The effect of parathyroid hormone on the numbers of nuclei in feline osteoclasts in vivo. J. Anat. 1980, 130, 479–486. [Google Scholar] [PubMed]
- Piper, K.; Boyde, A.; Jones, S.J. The relationship between the number of nuclei of an osteoclast and its resorptive capability in vitro. Anat. Embryol. 1992, 186, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Boissy, P.; Saltel, F.; Bouniol, C.; Jurdic, P.; Machuca-Gayet, I. Transcriptional activity of nuclei in multinucleated osteoclasts and its modulation by calcitonin. Endocrinology 2002, 143, 1913–1921. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.J.; Ali, N.N.; Boyde, A. Survival and resorptive activity of chick osteoclasts in culture. Anat. Embryol. 1986, 174, 265–275. [Google Scholar] [CrossRef]
- Chung, P.L.; Zhou, S.; Eslami, B.; Shen, L.; Leboff, M.S.; Glowacki, J. Effect of age on regulation of human osteoclast differentiation. J. Cell. Biochem. 2014, 115, 1412–1419. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Venton, L.; Sakata, T.; Halloran, B.P. Expression of RANKL and OPG correlates with age-related bone loss in male C57BL/6 mice. J. Bone Miner. Res. 2003, 18, 270–277. [Google Scholar] [CrossRef]
- Cao, J.J.; Wronski, T.J.; Iwaniec, U.; Phleger, L.; Kurimoto, P.; Boudignon, B.; Halloran, B.P. Aging increases stromal/osteoblastic cell-induced osteoclastogenesis and alters the osteoclast precursor pool in the mouse. J. Bone Miner. Res. 2005, 20, 1659–1668. [Google Scholar] [CrossRef]
- Salamanna, F.; Maglio, M.; Giavaresi, G.; Pagani, S.; Giardino, R.; Fini, M. In vitro method for the screening and monitoring of estrogen-deficiency osteoporosis by targeting peripheral circulating monocytes. Age 2015, 37, 9819. [Google Scholar] [CrossRef] [Green Version]
- Jevon, M.; Sabokbar, A.; Fujikawa, Y.; Hirayama, T.; Neale, S.D.; Wass, J.; Athanasou, N.A. Gender- and age-related differences in osteoclast formation from circulating precursors. J. Endocrinol. 2002, 172, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Pivetta, E.; Wassermann, B.; Bulian, P.; Steffan, A.; Colombatti, A.; Polesel, J.; Spessotto, P. Functional osteoclastogenesis: The baseline variability in blood donor precursors is not associated with age and gender. Oncotarget 2015, 6, 31889–31900. [Google Scholar] [CrossRef]
- Seidler, S.; Zimmermann, H.W.; Bartneck, M.; Trautwein, C.; Tacke, F. Age-dependent alterations of monocyte subsets and monocyte-related chemokine pathways in healthy adults. BMC Immunol. 2010, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, H.M.; Schnelle, J.F.; Thomas, J.K.; Nishanian, P.; Fahey, J.L. Phenotypic and functional characteristics of circulating monocytes of elderly persons. Exp. Gerontol. 1999, 34, 959–970. [Google Scholar] [CrossRef]
- Levaot, N.; Ottolenghi, A.; Mann, M.; Guterman-Ram, G.; Kam, Z.; Geiger, B. Osteoclast fusion is initiated by a small subset of RANKL-stimulated monocyte progenitors, which can fuse to RANKL-unstimulated progenitors. Bone 2015, 79, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Cundy, T. Paget’s disease of bone. Metab. Clin. Exp. 2018, 80, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appelman-Dijkstra, N.M.; Papapoulos, S.E. Paget’s disease of bone. Best Pract. Res. Clin. Endocrinol. Metab. 2018, 32, 657–668. [Google Scholar] [CrossRef]
- Albagha, O.M.E.; Wani, S.E.; Visconti, M.R.; Alonso, N.; Goodman, K.; Brandi, M.L.; Cundy, T.; Chung, P.Y.J.; Dargie, R.; Devogelaer, J.-P.; et al. Genome-wide association identifies three new susceptibility loci for Paget’s disease of bone. Nat. Genet. 2011, 43, 685–689. [Google Scholar]
- Mullin, B.H.; Zhu, K.; Brown, S.J.; Mullin, S.; Tickner, J.; Pavlos, N.J.; Dudbridge, F.; Xu, J.; Walsh, J.P.; Wilson, S.G. Genetic regulatory mechanisms in human osteoclasts suggest a role for the STMP1 and DCSTAMP genes in Paget’s disease of bone. Sci. Rep. 2019, 9, 1052. [Google Scholar] [CrossRef] [Green Version]
- Laurier, E.; Amiable, N.; Gagnon, E.; Brown, J.P.; Michou, L. Effect of a rare genetic variant of TM7SF4 gene on osteoclasts of patients with Paget’s disease of bone. BMC Med. Genet. 2017, 18, 133. [Google Scholar] [CrossRef] [Green Version]
- Coleman, R.; Cameron, D.; Dodwell, D.; Bell, R.; Wilson, C.; Rathbone, E.; Keane, M.; Gil, M.; Burkinshaw, R.; Grieve, R.; et al. Adjuvant zoledronic acid in patients with early breast cancer: Final efficacy analysis of the AZURE (BIG 01/04) randomised open-label phase 3 trial. Lancet Oncol. 2014, 15, 997–1006. [Google Scholar] [CrossRef]
- Zampieri, M.; Ciccarone, F.; Calabrese, R.; Franceschi, C.; Bürkle, A.; Caiafa, P. Reconfiguration of DNA methylation in aging. Mech. Ageing Dev. 2015, 151, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.A.; Tsai, P.C.; Joehanes, R.; Zheng, J.; Trajanoska, K.; Soerensen, M.; Forgetta, V.; Castillo-Fernandez, J.E.; Nielsen, M.M.F.; Spector, T.D.; et al. Epigenome-wide Association of DNA Methylation in Whole Blood With Bone Mineral Density. J. Bone Miner. Res. 2017, 32, 1644–1650. [Google Scholar] [CrossRef] [PubMed]
- Bahl, A.; Pöllänen, E.; Ismail, K.; Sipilä, S.; Mikkola, T.M.; Berglund, E.; Lindqvist, C.M.; Syvänen, A.-C.; Rantanen, T.; Kaprio, J.; et al. Hormone Replacement Therapy Associated White Blood Cell DNA Methylation and Gene Expression are Associated With Within-Pair Differences of Body Adiposity and Bone Mass. Twin Res. Hum. Genet. 2015, 18, 647–661. [Google Scholar] [CrossRef]
- Ghayor, C.; Weber, F.E. Epigenetic regulation of bone remodeling and its impacts in osteoporosis. Int. J. Mol. Sci. 2016, 17, 1446. [Google Scholar] [CrossRef] [Green Version]
- Reppe, S.; Lien, T.G.; Hsu, Y.H.; Gautvik, V.T.; Olstad, O.K.; Yu, R.; Bakke, H.G.; Lyle, R.; Kringen, M.K.; Glad, I.K.; et al. Distinct DNA methylation profiles in bone and blood of osteoporotic and healthy postmenopausal women. Epigenetics 2017, 12, 674–687. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Dong, S.S.; Chen, X.F.; Jing, Y.A.; Yang, M.; Yan, H.; Shen, H.; Chen, X.-D.; Tan, L.-J.; Tian, Q.; et al. Integrating Epigenomic Elements and GWASs Identifies BDNF Gene Affecting Bone Mineral Density and Osteoporotic Fracture Risk. Sci. Rep. 2016, 6, 30558. [Google Scholar] [CrossRef] [Green Version]
- Søe, K.; Delaissé, J.M. Glucocorticoids maintain human osteoclasts in the active mode of their resorption cycle. J. Bone Miner. Res. 2010, 25, 2184–2192. [Google Scholar] [CrossRef]
- Møller, A.M.J.; Füchtbauer, E.M.; Brüel, A.; Andersen, T.L.; Borggaard, X.G.; Pavlos, N.J.; Thomsen, J.; Pedersen, F.S.; Delaissé, J.-M.; Søe, K. Septins are critical regulators of osteoclastic bone resorption. Sci. Rep. 2018, 8, 13016. [Google Scholar] [CrossRef] [Green Version]
- Søe, K.; Andersen, T.L.; Hobolt-Pedersen, A.S.; Bjerregaard Bolette, B.; Larsson, L.I.; Delaissé, J.M. Involvement of human endogenous retroviral syncytin-1 in human osteoclast fusion. Bone 2011, 48, 837–846. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Features | n | % | |
|---|---|---|---|
| Age | 40–44 | 8 | 16.3 |
| 45–49 | 6 | 12.2 | |
| 50–54 | 15 | 30.6 | |
| 55–59 | 11 | 22.5 | |
| 60–66 | 9 | 18.4 | |
| Menopause Status | Premenopausal | 17 | 34.7 |
| Postmenopausal | 32 | 65.3 | |
| Smoking Status | Nonsmoker | 42 | 85.7 |
| Smoker | 7 | 14.3 | |
| Comorbidity | No | 41 | 83.7 |
| Yes | 8 | 16.3 | |
| Hypothyroidism | 3 | ||
| Asthma/Allergy | 3 | ||
| Ulcers | 2 | ||
| Median (IQR2) [Range] | |||
| Years since Menopause | 3.5 (8.25) [0; 23] | 49 | |
| Mean (SD3) [Range] | |||
| Age | 53.0 (6.7) [40; 66] | 49 | |
| Age of Premenopausal Women | 46.4 (5.0) [40; 54] | 17 | |
| Age of Postmenopausal Women | 56.5 (4.9) [45; 66] | 32 | |
| Height (m) | 1.70 (0.6) [1.56; 1.84] | 49 | |
| Weight (kg) | 73.2 (13.35) [55; 124] | 49 | |
| BMI4 | 25.4 (4.0) [19.5; 37.8] | 49 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Møller, A.M.J.; Delaissé, J.-M.; Olesen, J.B.; Canto, L.M.; Rogatto, S.R.; Madsen, J.S.; Søe, K. Fusion Potential of Human Osteoclasts In Vitro Reflects Age, Menopause, and In Vivo Bone Resorption Levels of Their Donors—A Possible Involvement of DC-STAMP. Int. J. Mol. Sci. 2020, 21, 6368. https://doi.org/10.3390/ijms21176368
Møller AMJ, Delaissé J-M, Olesen JB, Canto LM, Rogatto SR, Madsen JS, Søe K. Fusion Potential of Human Osteoclasts In Vitro Reflects Age, Menopause, and In Vivo Bone Resorption Levels of Their Donors—A Possible Involvement of DC-STAMP. International Journal of Molecular Sciences. 2020; 21(17):6368. https://doi.org/10.3390/ijms21176368
Chicago/Turabian StyleMøller, Anaïs M. J., Jean-Marie Delaissé, Jacob B. Olesen, Luisa M. Canto, Silvia R. Rogatto, Jonna S. Madsen, and Kent Søe. 2020. "Fusion Potential of Human Osteoclasts In Vitro Reflects Age, Menopause, and In Vivo Bone Resorption Levels of Their Donors—A Possible Involvement of DC-STAMP" International Journal of Molecular Sciences 21, no. 17: 6368. https://doi.org/10.3390/ijms21176368
APA StyleMøller, A. M. J., Delaissé, J. -M., Olesen, J. B., Canto, L. M., Rogatto, S. R., Madsen, J. S., & Søe, K. (2020). Fusion Potential of Human Osteoclasts In Vitro Reflects Age, Menopause, and In Vivo Bone Resorption Levels of Their Donors—A Possible Involvement of DC-STAMP. International Journal of Molecular Sciences, 21(17), 6368. https://doi.org/10.3390/ijms21176368