The Interplay between Toxic and Essential Metals for Their Uptake and Translocation Is Likely Governed by DNA Methylation and Histone Deacetylation in Maize

 ,
, 

Abstract
:1. Introduction
2. Results
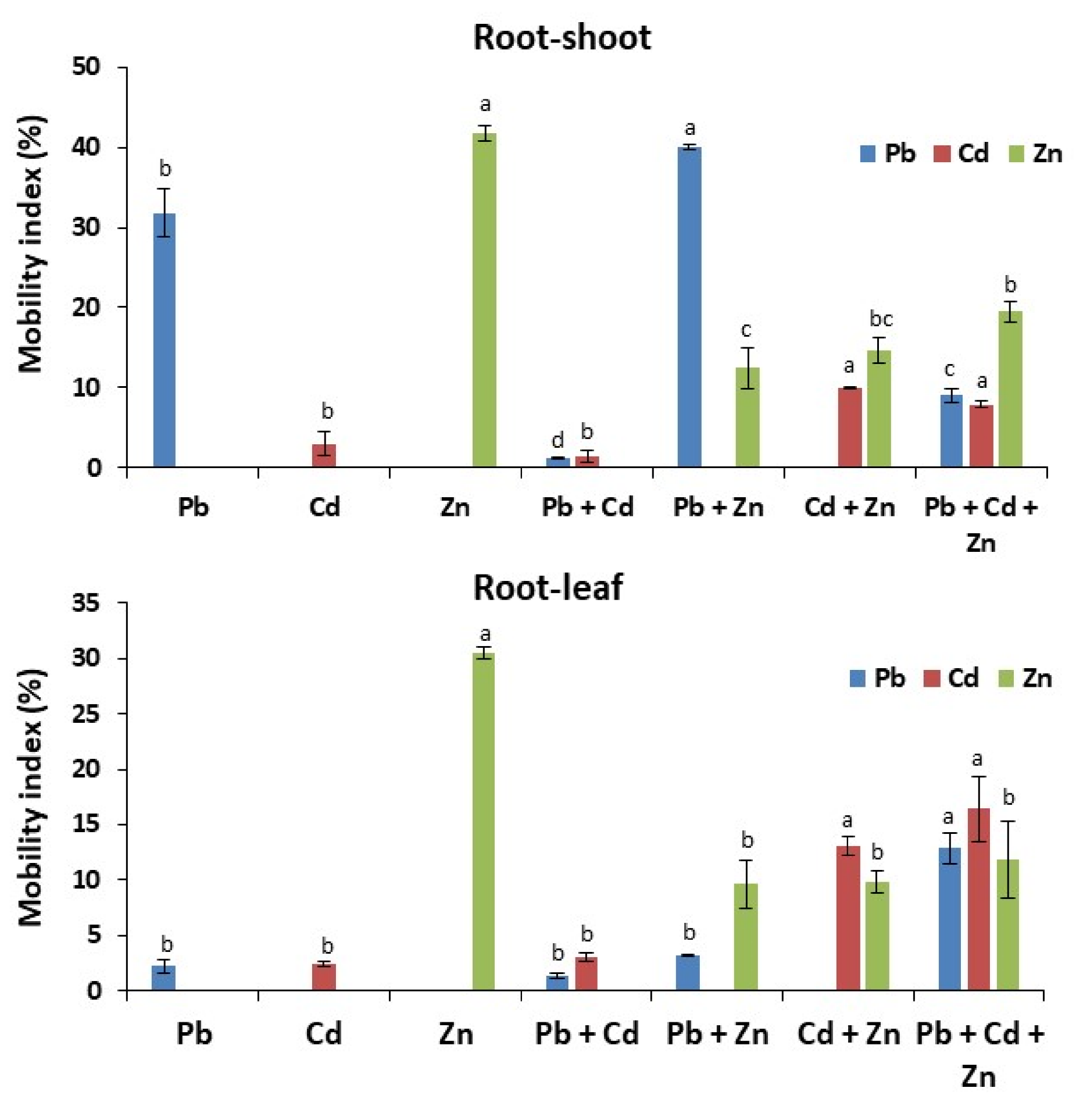
2.1. Zn Favors the Accumulation and Mobility of Pb/Cd to the Aerial Parts of Maize Plants
2.2. Pb/Cd Block the Accumulation and Transport of Zn to the Aerial Parts of Maize Plants
2.3. Zn, Pb, and Cd Influence the Mobility of Divalent Calcium (Ca) and Magnesium (Mg) But Not the Monovalent Potassium (K)
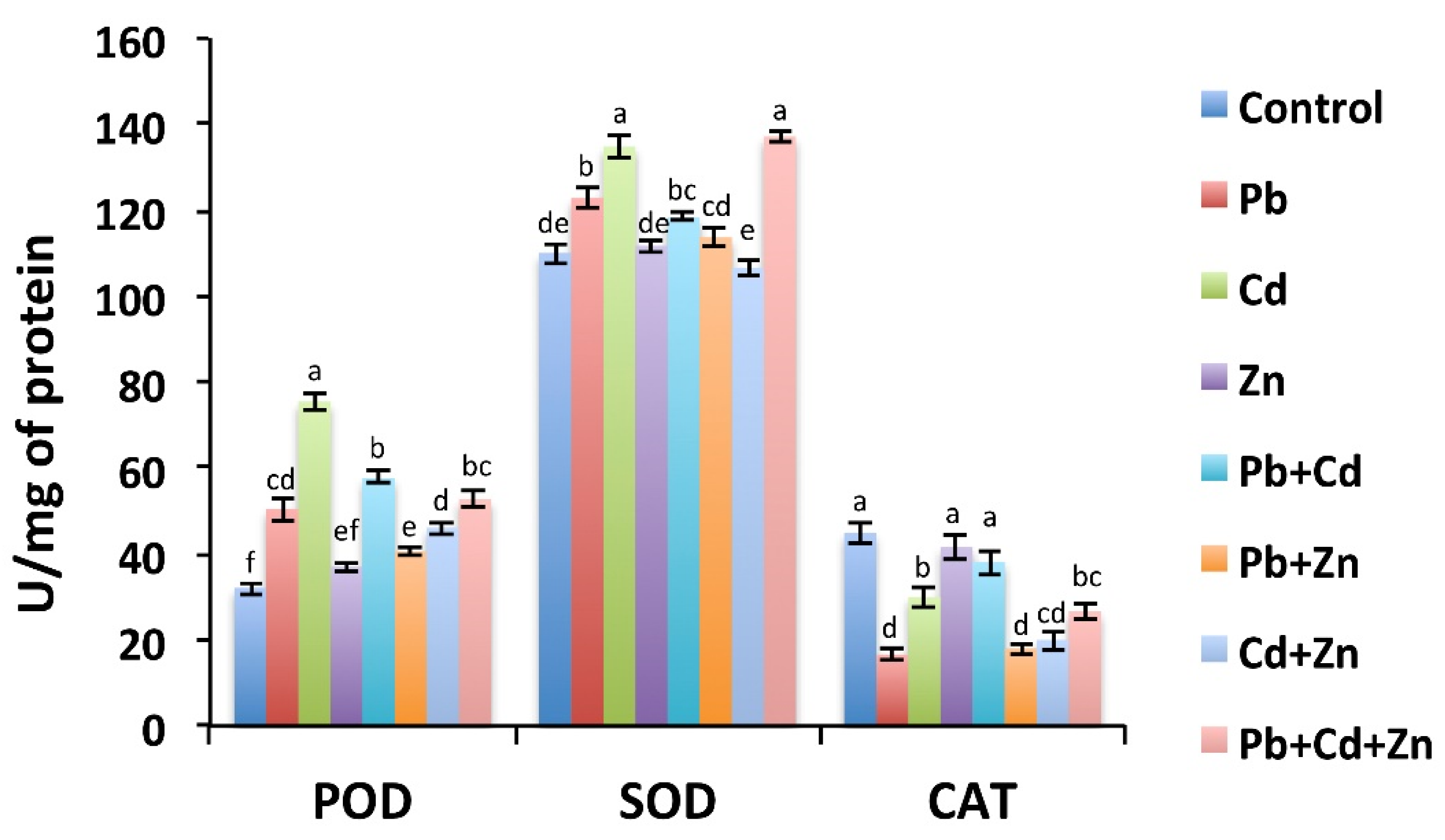
2.4. Antioxidant Activity Is Altered in Response to Zn, Pb, and Cd Applied Alone and in Combinations
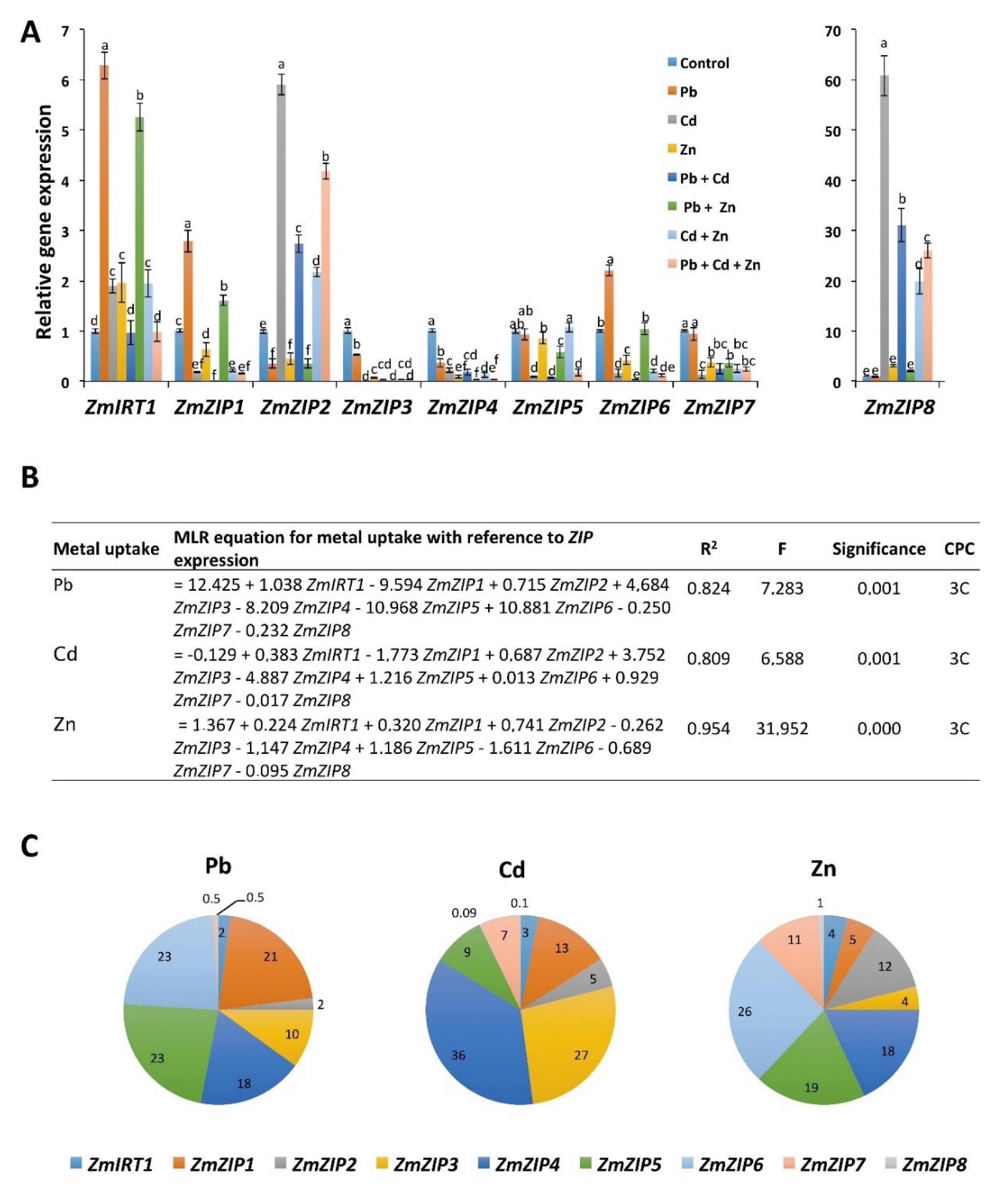
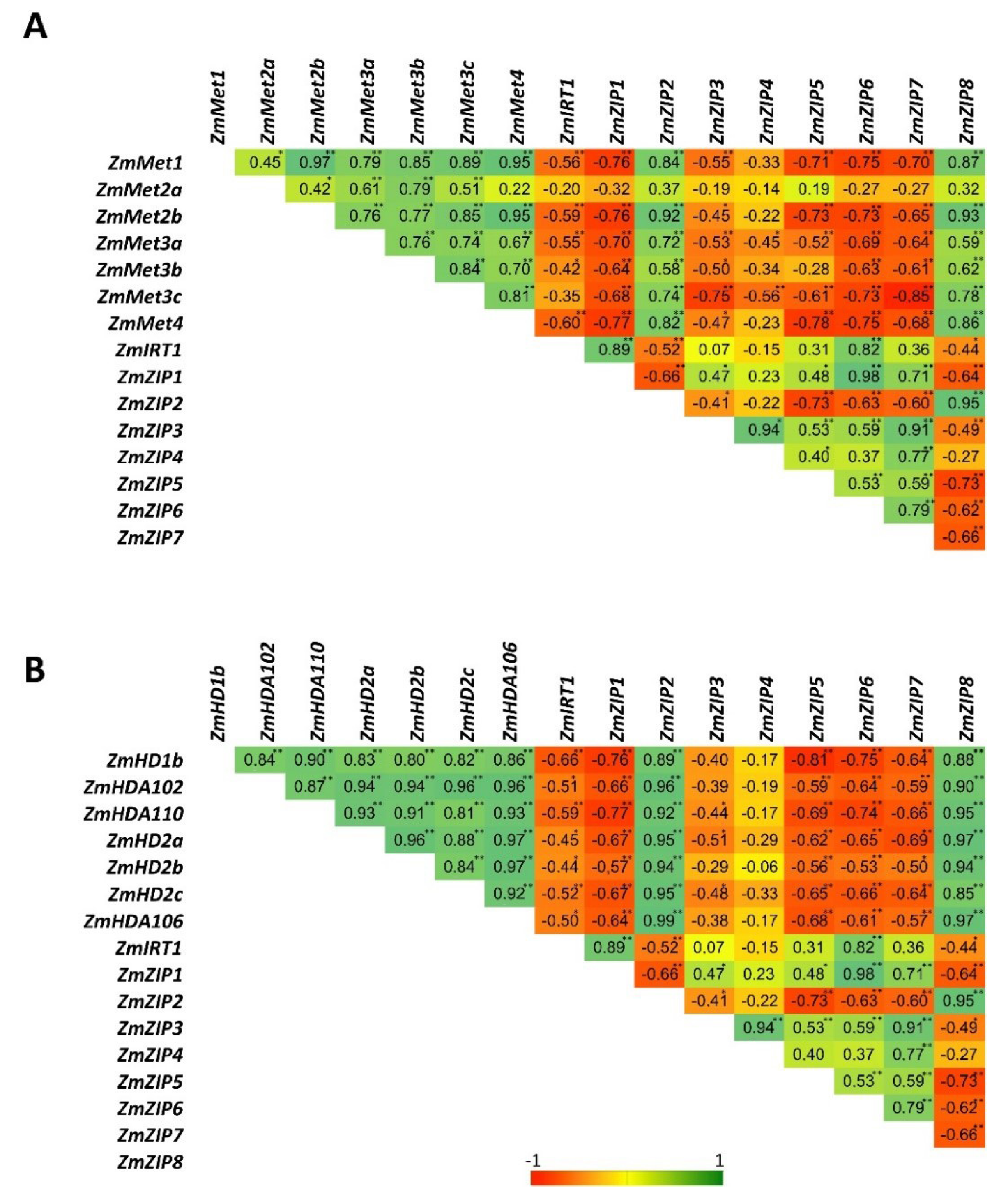
2.5. Pb, Cd, and Zn Alone and in Combinations Differentially Regulate the Expression of ZIP Transporters
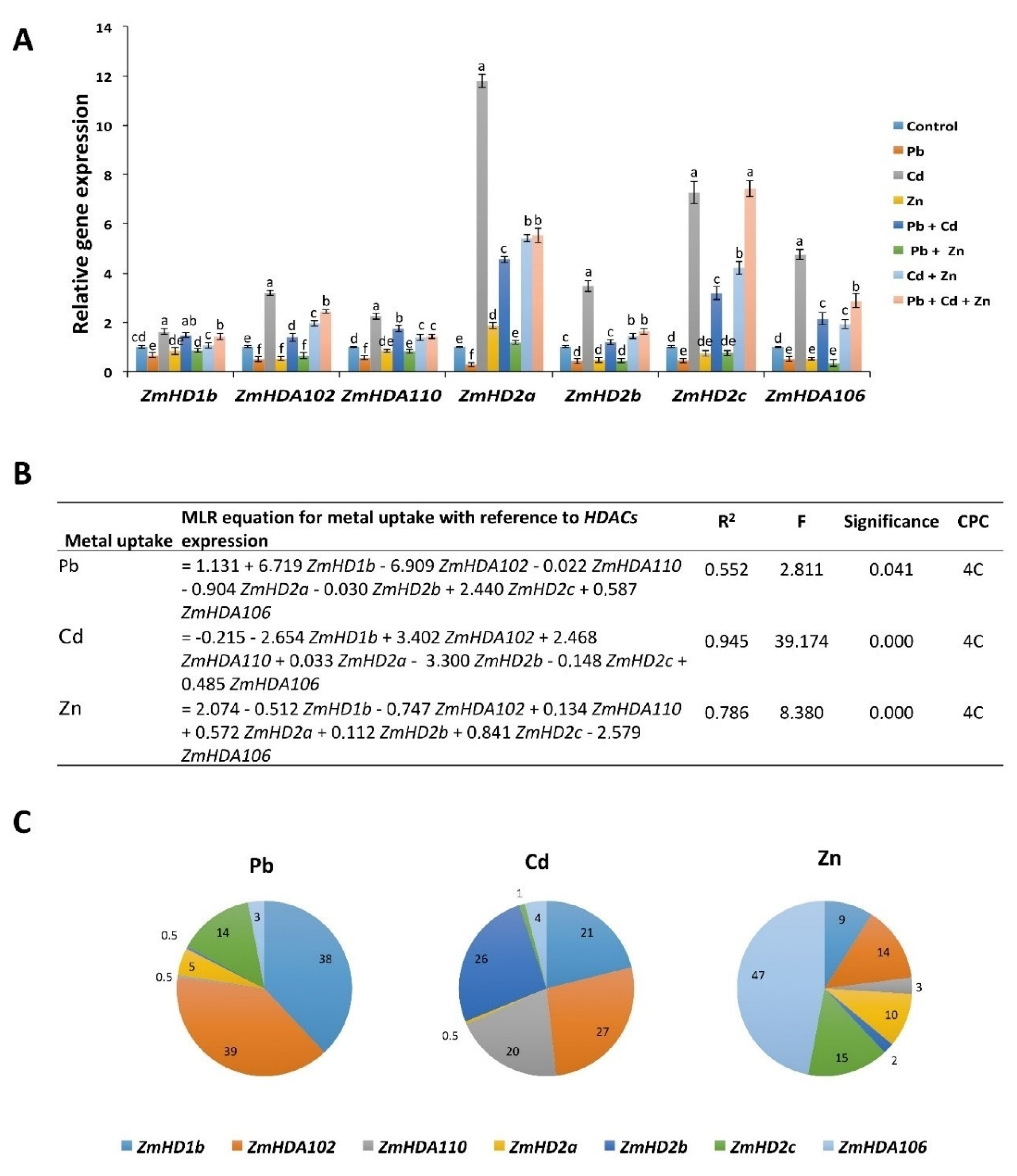
2.6. Pb, Cd, and Zn Alone and in Combinations Differentially Regulate the Expression of Histone Deacetylases (HDACs)
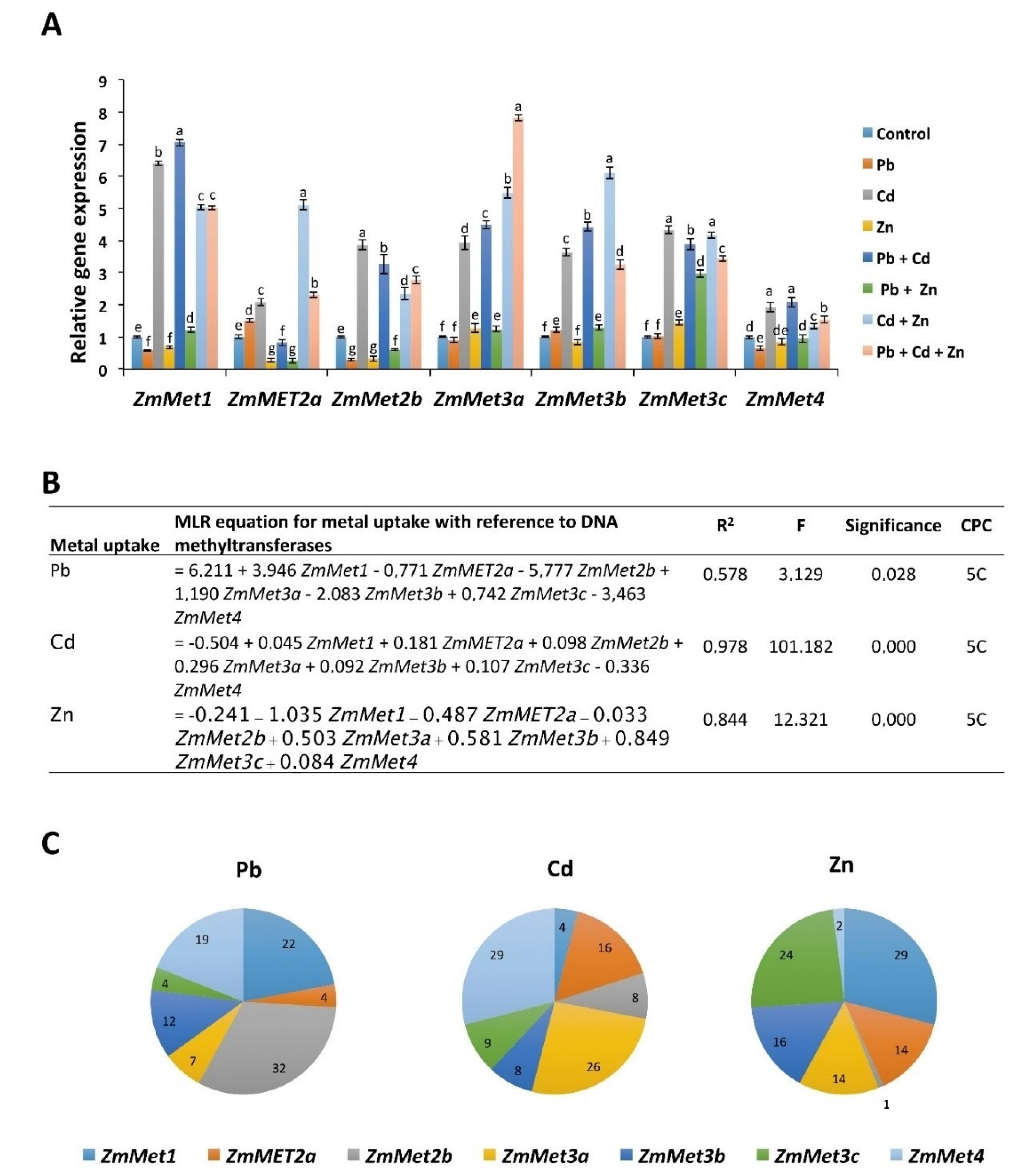
2.7. Pb, Cd, and Zn Alone and in Combinations Differentially Regulate the Expression of DNA Methyltransferases
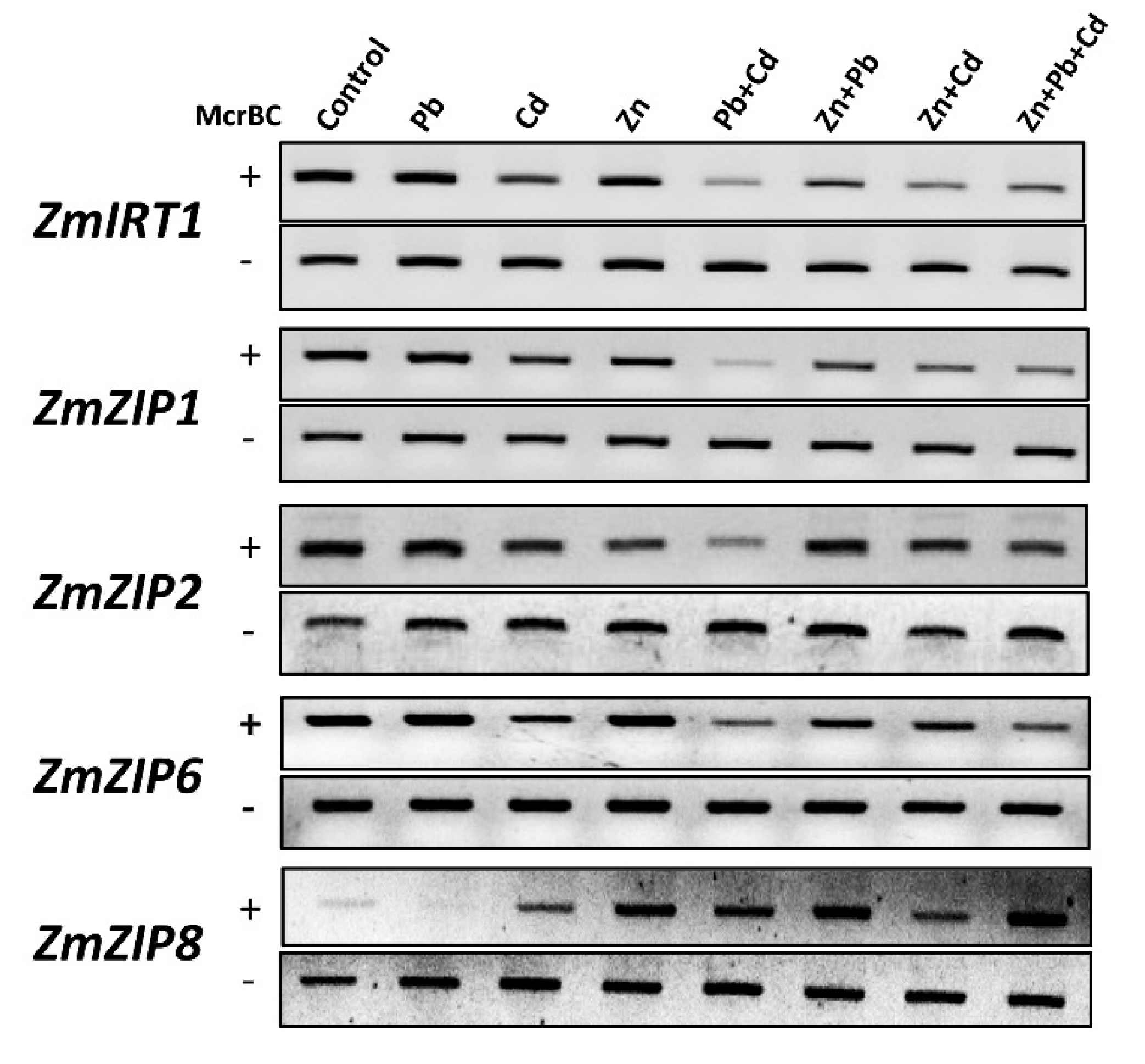
2.8. Zn, Pb, and Cd Combinations Alter the DNA Methylation Levels at the Promoter of ZIP Transporters
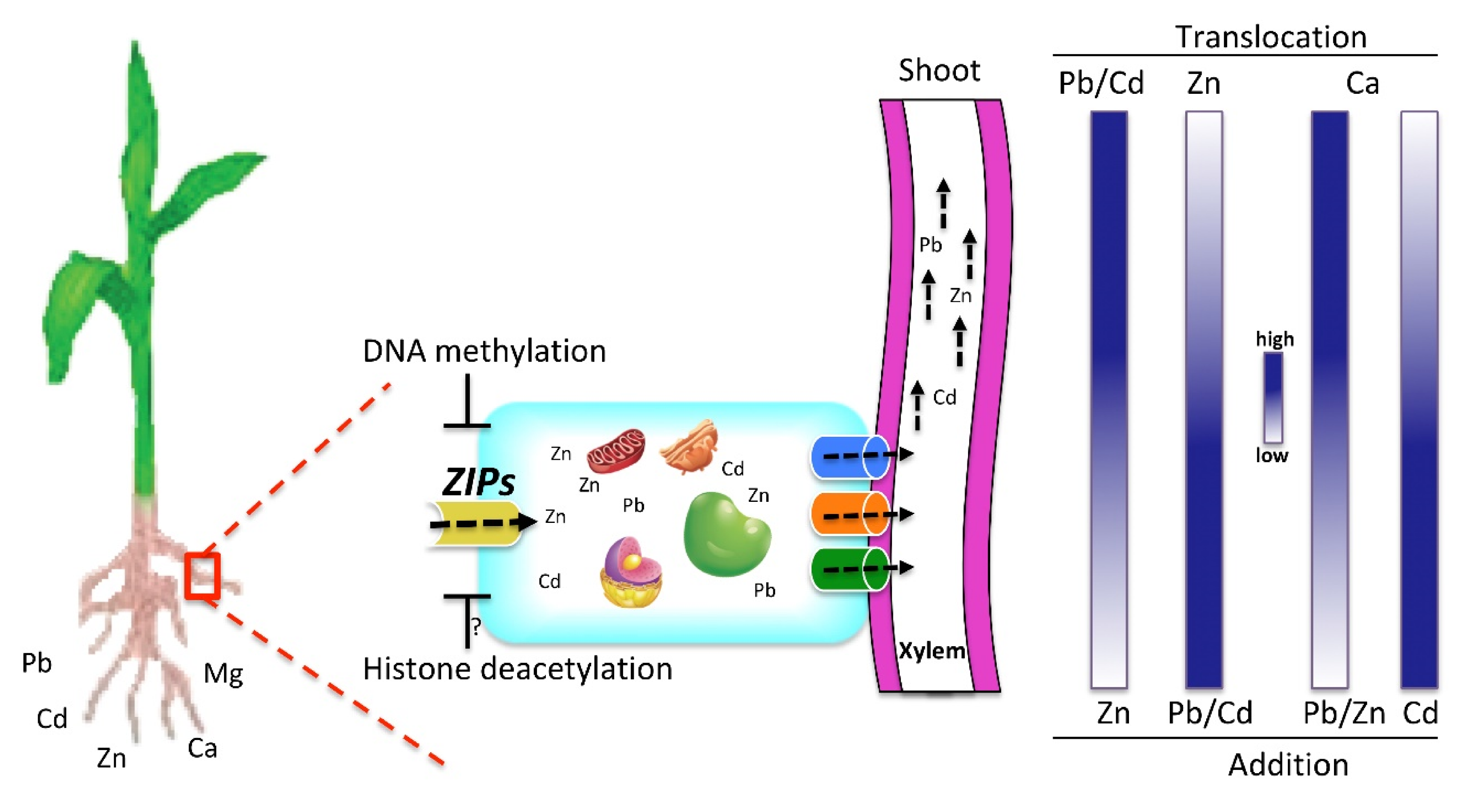
3. Discussion
4. Materials and Methods
4.1. Hydroponics
4.2. Atomic Absorption Analysis
4.3. Extraction and Quantification of Antioxidant Enzymes
4.4. Gene Expression Analysis
4.5. ChoP-PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Singh, S.; Parihar, P.; Singh, R.; Singh, D.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovečka, M.; Takáč, T. Managing heavy metal toxicity stress in plants: Biological and biotechnological tools. Biotechnol. Adv. 2014, 32, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, S.; Liu, R.; Zhou, W.; Jin, J. Changes of photosynthetic activities of maize (Zea mays L.) seedlings in response to cadmium stress. Photosynthetica 2009, 47, 277–283. [Google Scholar] [CrossRef]
- Cheng, W.-D.; Zhang, G.; Yao, H.-G.; Wu, W.; Xu, M. Genotypic and environmental variation in cadmium, chromium, arsenic, nickel, and lead concentrations in rice grains. J. Zhejiang Univ. Sci. B 2006, 7, 565–571. [Google Scholar] [CrossRef] [Green Version]
- Fahr, M.; Laplaze, L.; Bendaou, N.; Hocher, V.; El Mzibri, M.; Bogusz, D.; Smouni, A. Effect of lead on root growth. Front. Plant Sci. 2013, 4, 175. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Nishizawa, N.K. Iron Uptake, Translocation, and Regulation in Higher Plants. Annu. Rev. Plant Boil. 2012, 63, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E. Essential and Beneficial Trace Elements in Plants, and Their Transport in Roots: A Review. Appl. Biochem. Biotechnol. 2016, 181, 464–482. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef]
- Morrissey, J.; Guerinot, M.L. Iron Uptake and Transport in Plants: The Good, the Bad, and the Ionome. Chem. Rev. 2009, 109, 4553–4567. [Google Scholar] [CrossRef] [Green Version]
- Sadeghzadeh, B.; Rengel, Z. Zinc in Soils and Crop Nutrition. In The Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; Wiley: Chichester, UK; Hoboken, NJ, USA, 2011; pp. 335–375. [Google Scholar]
- Küpper, H.; Andresen, E. Mechanisms of metal toxicity in plants. Metallomics 2016, 8, 269–285. [Google Scholar] [CrossRef]
- Song, W.-Y.; Park, J.; Eisenach, C.; Maeshima, M.; Lee, Y.; Martinoia, E. ABC Transporters and Heavy Metals. In Plant ABC Transporters. Signaling and Communication in Plants; Springer: Cham, Switzerland, 2014; pp. 1–17. [Google Scholar]
- Milner, M.J.; Seamon, J.; Craft, E.; Kochian, L. Transport properties of members of the ZIP family in plants and their role in Zn and Mn homeostasis. J. Exp. Bot. 2012, 64, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiong, J.; McDonald, G.; Genç, Y.; Shirley, N.J.; Langridge, P.; Huang, C.Y. Increased expression of sixZIPfamily genes by zinc (Zn) deficiency is associated with enhanced uptake and root-to-shoot translocation of Zn in barley (Hordeum vulgare). New Phytol. 2015, 207, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Evens, N.P.; Buchner, P.; Williams, L.E.; Hawkesford, M.J. The role of ZIP transporters and group F bZIP transcription factors in the Zn-deficiency response of wheat (Triticum aestivum). Plant J. 2017, 92, 291–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta (BBA) Biomembr. 2000, 1465, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhou, X.; Huang, Y.; Zhu, L.; Zhang, S.; Zhao, Y.; Guo, J.; Chen, J.; Chen, R. Identification and characterization of the zinc-regulated transporters, iron-regulated transporter-like protein (ZIP) gene family in maize. BMC Plant Boil. 2013, 13, 114. [Google Scholar] [CrossRef] [Green Version]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.D.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D.; et al. Phylogenetic Relationships within Cation Transporter Families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef] [Green Version]
- Arita, A.; Costa, M. Epigenetics in metal carcinogenesis: Nickel, arsenic, chromium and cadmium. Metallomics 2009, 1, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Berr, A.; Shafiq, S.; Shen, W.-H. Histone modifications in transcriptional activation during plant development. Biochim. Biophys. Acta (BBA) Bioenergy 2011, 1809, 567–576. [Google Scholar] [CrossRef]
- Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. Reduced DNA methylation in Arabidopsis thaliana results in abnormal plant development. Proc. Natl. Acad. Sci. USA 1996, 93, 8449–8454. [Google Scholar] [CrossRef] [Green Version]
- Gehring, M. Genomic Imprinting: Insights From Plants. Annu. Rev. Genet. 2013, 47, 187–208. [Google Scholar] [CrossRef]
- Kakutani, T.; Jeddeloh, J.A.; Flowers, S.K.; Munakata, K.; Richards, E.J. Developmental abnormalities and epimutations associated with DNA hypomethylation mutations. Proc. Natl. Acad. Sci. USA 1996, 93, 12406–12411. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.; Xi, Y.; Cheng, B.; Zhu, S. Genome-wide identification and expression profiling of DNA methyltransferase gene family in maize. Plant Cell Rep. 2014, 33, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Aina, R.; Sgorbati, S.; Santagostino, A.; Labra, M.; Ghiani, A.; Citterio, S. Specific hypomethylation of DNA is induced by heavy metals in white clover and industrial hemp. Physiol. Plant 2004, 121, 472–480. [Google Scholar] [CrossRef]
- Filek, M.; Keskinen, R.; Hartikainen, H.; Szarejko, I.; Janiak, A.; Miszalski, Z.; Golda, A. The protective role of selenium in rape seedlings subjected to cadmium stress. J. Plant Physiol. 2008, 165, 833–844. [Google Scholar] [CrossRef]
- Yang, J.-L.; Liu, L.; Gong, Y.; Huang, D.-Q.; Wang, F.; He, L.-L. Analysis of genomic DNA methylation level in radish under cadmium stress by methylation-sensitive amplified polymorphism technique. Zhi Wu Sheng Li Yu Fen Zi Sheng Wu Xue Xue Bao = J. Plant Physiol. Mol. Boil. 2007, 33, 219–226. [Google Scholar]
- Shafiq, S.; Zeb, Q.; Ali, A.; Sajjad, Y.; Nazir, R.; Widemann, E.; Liu, L. Lead, Cadmium and Zinc Phytotoxicity Alter DNA Methylation Levels to Confer Heavy Metal Tolerance in Wheat. Int. J. Mol. Sci. 2019, 20, 4676. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, T.; Okamoto, T.; Kohno, Y.; Fukiage, K.; Otsuka, S.; Furu, M.; Ito, K.; Jin, Y.; Nagayama, S.; Nakayama, T.; et al. Cell-specific epigenetic regulation of ChM-I gene expression: Crosstalk between DNA methylation and histone acetylation. Biochem. Biophys. Res. Commun. 2008, 365, 124–130. [Google Scholar] [CrossRef]
- Tian, L.; Chen, Z.J. Blocking histone deacetylation in Arabidopsis induces pleiotropic effects on plant gene regulation and development. Proc. Natl. Acad. Sci. USA 2001, 98, 200–205. [Google Scholar] [CrossRef]
- Luo, M.; Wang, Y.-Y.; Liu, X.; Yang, S.; Lu, Q.; Cui, Y.; Wu, K. HD2C interacts with HDA6 and is involved in ABA and salt stress response in Arabidopsis. J. Exp. Bot. 2012, 63, 3297–3306. [Google Scholar] [CrossRef]
- Cigliano, R.A.; Cremona, G.; Paparo, R.; Termolino, P.; Perrella, G.; Gutzat, R.; Consiglio, F.; Conicella, C. Histone Deacetylase AtHDA7 Is Required for Female Gametophyte and Embryo Development in Arabidopsis. Plant Physiol. 2013, 163, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Shafiq, S.; Naeem, M.K.; Widemann, E.; Munir, M.Z.; Jensen, K.B.; Wang, R.R.-C. Histone Deacetylase (HDAC) Gene Family in Allotetraploid Cotton and Its Diploid Progenitors: In Silico Identification, Molecular Characterization, and Gene Expression Analysis under Multiple Abiotic Stresses, DNA Damage and Phytohormone Treatments. Int. J. Mol. Sci. 2020, 21, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Shafiq, S.; Farooq, M.A.; Naeem, M.K.; Widemann, E.; Bakhsh, A.; Jensen, K.B.; Wang, R.R.-C. Comparative Genome-wide Analysis and Expression Profiling of Histone Acetyltransferase (HAT) Gene Family in Response to Hormonal Applications, Metal and Abiotic Stresses in Cotton. Int. J. Mol. Sci. 2019, 20, 5311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varotto, S.; Locatelli, S.; Canova, S.; Pipal, A.; Motto, M.; Rossi, V. Expression Profile and Cellular Localization of Maize Rpd3-Type Histone Deacetylases during Plant Development1. Plant Physiol. 2003, 133, 606–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, V.; Locatelli, S.; Varotto, S.; Donn, G.; Pirona, R.; Henderson, D.A.; Hartings, H.; Motto, M. Maize Histone Deacetylase hda101 Is Involved in Plant Development, Gene Transcription, and Sequence-Specific Modulation of Histone Modification of Genes and Repeats. Plant Cell 2007, 19, 1145. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Liu, X.; Xin, M.; Du, J.; Hu, Z.; Peng, H.; Rossi, V.; Sun, Q.; Ni, Z.; Yao, Y. Genome-Wide Mapping of Targets of Maize Histone Deacetylase HDA101 Reveals Its Function and Regulatory Mechanism during Seed Development. Plant Cell 2016, 28, 629–645. [Google Scholar] [CrossRef] [Green Version]
- Hou, H.; Zheng, X.; Zhang, H.; Yue, M.; Hu, Y.; Zhou, H.; Wang, Q.; Xie, C.; Wang, P.; Li, L. Histone Deacetylase Is Required for GA-Induced Programmed Cell Death in Maize Aleurone Layers. Plant Physiol. 2017, 175, 1484–1496. [Google Scholar] [CrossRef] [Green Version]
- Ekmekçi, Y.; Tanyolaç, D.; Ayhan, B. Effects of cadmium on antioxidant enzyme and photosynthetic activities in leaves of two maize cultivars. J. Plant Physiol. 2008, 165, 600–611. [Google Scholar] [CrossRef]
- Al-Mureish, K.; Othman, N.A.R.M.; Al-Hakimi, A.M.A. Salicylic Acid-Mediated Alleviation of Cadmium Toxicity in Maize Leaves. J. Plant Sci. (Sci. Publ. Group) 2014, 2, 276–281. [Google Scholar] [CrossRef]
- Kankel, M.W.; Ramsey, D.E.; Stokes, T.L.; Flowers, S.K.; Haag, J.R.; Jeddeloh, J.A.; Riddle, N.C.; Verbsky, M.L.; Richards, E.J. Arabidopsis MET1 cytosine methyltransferase mutants. Genetics 2003, 163, 1109–1122. [Google Scholar]
- Lindroth, A.M.; Cao, X.; Jackson, J.P.; Zilberman, D.; McCallum, C.M.; Henikoff, S.; Jacobsen, S.E. Requirement of CHROMOMETHYLASE3 for Maintenance of CpXpG Methylation. Science 2001, 292, 2077–2080. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Jacobsen, S.E. Role of the Arabidopsis DRM Methyltransferases in De Novo DNA Methylation and Gene Silencing. Curr. Boil. 2002, 12, 1138–1144. [Google Scholar] [CrossRef] [Green Version]
- Taspinar, M.S.; Agar, G.; Alpsoy, L.; Yildirim, N.; Bozari, S.; Sevsay, S. The protective role of zinc and calcium in Vicia faba seedlings subjected to cadmium stress. Toxicol. Ind. Health 2010, 27, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, X.; Li, S.; Wang, Z. Effect of nickel chloride on Arabidopsis genomic DNA and methylation of 18S rDNA. Electron. J. Biotechnol. 2015, 18, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Ogutcu, H.; Arslan, E.; Agar, G.; Gulluce, M.; Turan, M.; Sahin, F. Protective Role of Calcium on DNA Methylation Caused Cadmium Stress in Vicia Faba Seedlings. In Proceedings of the International Conference on Agricultural, Ecological and Medical Sciences, Bali, Indonesia, 6–7 February 2014. [Google Scholar]
- Li, X.; Yang, Y.; Jia, L.; Chen, H.; Wei, X. Zinc-induced oxidative damage, antioxidant enzyme response and proline metabolism in roots and leaves of wheat plants. Ecotoxicol. Environ. Saf. 2013, 89, 150–157. [Google Scholar] [CrossRef]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.-F.; Curie, C. IRT1, an Arabidopsis Transporter Essential for Iron Uptake from the Soil and for Plant Growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korshunova, Y.O.; Eide, D.; Clark, W.G.; Guerinot, M.L.; Pakrasi, H.B. The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range. Plant Mol. Boil. 1999, 40, 37–44. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef]
- Palmer, C.; Guerinot, M.L. A Question of Balance: Facing the challenges of Cu, Fe and Zn Homeostasis. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.A.; Piyatida, P.; Da Silva, J.A.T.; Fujita, M. Molecular Mechanism of Heavy Metal Toxicity and Tolerance in Plants: Central Role of Glutathione in Detoxification of Reactive Oxygen Species and Methylglyoxal and in Heavy Metal Chelation. J. Bot. 2012, 2012, 1–37. [Google Scholar] [CrossRef]
- Dresler, S.; Hanaka, A.; Bednarek, W.; Maksymiec, W. Accumulation of low-molecular-weight organic acids in roots and leaf segments of Zea mays plants treated with cadmium and copper. Acta Physiol. Plant 2014, 36, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Ridošková, A.; Sobrova, P.; Kryštofová, O.; Sochor, J.; Zitka, O.; Babula, P.; Adam, V.; Docekalová, H.; Kizek, R. Cadmium(II) and Zinc(II) Ions Effects on Maize Plants revealed by Spectroscopy and Electrochemistry. Int. J. Electrochem. Sci. 2011, 6, 6011–6031. [Google Scholar]
- Małkowski, E.; Kita, A.; Galas, W.; Karcz, W.; Kuperberg, J.M. Lead distribution in corn seedlings (Zea mays L.) and its effect on growth and the concentrations of potassium and calcium. Plant Growth Regul. 2002, 37, 69–76. [Google Scholar] [CrossRef]
- Helrich, K. Official Methods of Analysis of the Association of Official Analytical Chemisty, 15th ed.; The Association: Arlington, VA, USA, 1990. [Google Scholar]
- Csiszár, J.; Lantos, E.; Tari, I.; Madosa, E.; Wodala, B.; Vashegyi, Á.; Horváth, F.; Pécsváradi, A.; Szabó, M.; Bartha, B.; et al. Antioxidant enzyme activities in Allium species and their cultivars under water stress. Plant Soil Environ. 2008, 53, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Boil. Chem. 1951, 193, 265–275. [Google Scholar]
- Berr, A.; Shafiq, S.; Pinon, V.; Dong, A.; Shen, W.-H. The trxG family histone methyltransferase SET DOMAIN GROUP 26 promotes flowering via a distinctive genetic pathway. Plant J. 2014, 81, 316–328. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, K.; Wang, B.; Duan, C.-G.; Lang, Z.; Zhu, J.-K. Protocol: A beginner’s guide to the analysis of RNA-directed DNA methylation in plants. Plant Methods 2014, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.H.A.; Nawaz, I.; Qu, Z.; Butt, T.A.; Yousaf, S.; Iqbal, M. Reduced growth response of ornamental plant Nicotiana alata L. upon selected heavy metals uptake, with co-application of ethylenediaminetetraacetic acid. Chemosphere 2019, 241, 125006. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Pb (mg/g) | Cd (mg/g) | Zn (mg/g) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf | Shoot | Root | Total | Leaf | Shoot | Root | Total | Leaf | Shoot | Root | Total | |
| Control | 0.10 c | 0.08 d | 0.08 d | 0.27 c | 0.01 c | 0.01 d | 0.01 c | 0.03 c | 0.10 cd | 0.03 d | 0.08 d | 0.22 d |
| Pb | 0.12 c | 1.78 b | 5.62 c | 7.52 b | 0.01 c | 0.01 d | 0.02 c | 0.04 c | 0.03 d | 0.04 d | 0.06 d | 0.14 d |
| Cd | 0.11 c | 0.22 d | 0.09 d | 0.42 c | 0.04 c | 0.05 c | 1.92 b | 2.02 b | 0.06 d | 0.04 d | 0.07 d | 0.19 d |
| Zn | 0.09 c | 0.06 d | 0.10 d | 0.26 c | 0.03 c | 0.01 d | 0.02 c | 0.06 c | 0.67 a | 0.92 a | 2.20 a | 3.80 a |
| Pb + Cd | 0.09 c | 0.09 d | 7.23 b | 7.43 b | 0.05 c | 0.02 cd | 1.68 b | 1.75 b | 0.03 d | 0.03 d | 0.11 d | 0.18 d |
| Pb + Zn | 0.22 b | 2.76 a | 6.90 b | 9.89 a | 0.04 c | 0.03 cd | 0.11 c | 0.19 c | 0.19 bc | 0.24 c | 1.98 c | 2.41 c |
| Cd + Zn | 0.13 c | 0.14 d | 0.17 d | 0.99 c | 0.40 b | 0.30 a | 3.07 a | 3.78 a | 0.20 bc | 0.30 c | 2.06 b | 2.57 bc |
| Pb + Cd + Zn | 1.10 a | 0.77 c | 8.60 a | 10.49 a | 0.47 a | 0.22 b | 2.88 a | 3.58 a | 0.25 b | 0.41 b | 2.11 ab | 2.78 b |
| Treatments | Ca (mg/g) | Mg (mg/g) | ||||||
|---|---|---|---|---|---|---|---|---|
| Leaf | Shoot | Root | Total | Leaf | Shoot | Root | Total | |
| Control | 20.76 c | 17.08 b | 25.45 b | 63.29 bc | 5.175 ab | 3.22 cd | 2.55 e | 10.96 de |
| Pb | 21.78 bc | 27.59 a | 10.68 e | 60.06 c | 3.47 d | 5.53 a | 3.57 d | 12.57 cd |
| Cd | 12.03 d | 8.99 c | 20.69bcd | 41.72 d | 4.301 bcd | 6.07 a | 5.46 ab | 15.84 ab |
| Zn | 26.29 b | 26.29 a | 39.34 a | 91.93 a | 4.98 ab | 4.82 ab | 5.67 ab | 15.48 ab |
| Pb + Cd | 15.20 d | 9.28 c | 21.89 bc | 46.38 d | 3.78 cd | 2 d | 4.25 cd | 10.04 e |
| Pb + Zn | 36.52 a | 26.90 a | 25.77 b | 89.21 a | 5.56 a | 4.03 bc | 4.90 bc | 14.51 bc |
| Cd + Zn | 21.05 c | 29.07 a | 19.24 cd | 69.37 b | 4.54 bc | 5.40 a | 5.47 ab | 15.42 ab |
| Pb + Cd + Zn | 26.62 b | 27.86 a | 16.32 d | 70.81 b | 5.56 a | 5.46 a | 5.98 a | 17.01 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shafiq, S.; Ali, A.; Sajjad, Y.; Zeb, Q.; Shahzad, M.; Khan, A.R.; Nazir, R.; Widemann, E. The Interplay between Toxic and Essential Metals for Their Uptake and Translocation Is Likely Governed by DNA Methylation and Histone Deacetylation in Maize. Int. J. Mol. Sci. 2020, 21, 6959. https://doi.org/10.3390/ijms21186959
Shafiq S, Ali A, Sajjad Y, Zeb Q, Shahzad M, Khan AR, Nazir R, Widemann E. The Interplay between Toxic and Essential Metals for Their Uptake and Translocation Is Likely Governed by DNA Methylation and Histone Deacetylation in Maize. International Journal of Molecular Sciences. 2020; 21(18):6959. https://doi.org/10.3390/ijms21186959
Chicago/Turabian StyleShafiq, Sarfraz, Asim Ali, Yasar Sajjad, Qudsia Zeb, Muhammad Shahzad, Abdul Rehman Khan, Rashid Nazir, and Emilie Widemann. 2020. "The Interplay between Toxic and Essential Metals for Their Uptake and Translocation Is Likely Governed by DNA Methylation and Histone Deacetylation in Maize" International Journal of Molecular Sciences 21, no. 18: 6959. https://doi.org/10.3390/ijms21186959
APA StyleShafiq, S., Ali, A., Sajjad, Y., Zeb, Q., Shahzad, M., Khan, A. R., Nazir, R., & Widemann, E. (2020). The Interplay between Toxic and Essential Metals for Their Uptake and Translocation Is Likely Governed by DNA Methylation and Histone Deacetylation in Maize. International Journal of Molecular Sciences, 21(18), 6959. https://doi.org/10.3390/ijms21186959