Advances in D-Amino Acids in Neurological Research


Abstract
:1. Introduction
1.1. Sources of D-Amino Acids
1.2. Elimination of D-Amino Acids
2. NMDA Receptor Agonists or Co-Agonists
2.1. D-Alanine
2.2. D-Aspartate
2.3. D-Glutamate
2.4. D-Serine
3. Neuroactive D-Isomers without Direct NMDA Receptor Interaction
3.1. D-Isoleucine
3.2. D-Leucine
3.3. D-Phenylalanine
3.4. D-Threonine
3.5. D-Tyrosine
4. Inactive D-Amino Acids with Interesting Derivatives or Other Activities
4.1. D-Arginine
4.2. D-Cysteine
4.3. D-Histidine
4.4. D-Lysine
4.5. D-Methionine
4.6. D-Proline
4.7. D-Tryptophan
4.8. D-Valine
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NMDA | N-methyl-D-aspartate |
| DAAO | D-amino acid oxidase |
| DDO | D-aspartate oxidase |
| Asc1 | Alanine-serine-cysteine-1 |
| TAS1R2/R3 | Taste receptor type 1 member 2/3 |
| iNOS | Inducible nitric oxide synthase |
| R18D | 18 amino acid poly-D-arginine-peptide |
| H2S | Hydrogen sulfide |
| D-cis-Fpro | Cis-4-[18F]fluoro-D-proline |
| PET | Positronic emissions tomography |
| 5F-ADB | N-[[1-(5-fluoropentyl)-1H-indazol-3-yl]carbonyl]-3-methyl-D-valine methyl ester |
References
- Neuberger, A. Stereochemistry of amino acids. Adv. Protein. Chem. 1948, 4, 297–383. [Google Scholar] [CrossRef] [PubMed]
- Stevens, C.M.; Halpern, P.E.; Gigger, R.P. Occurrence of D-amino acids in some natural materials. J. Boil. Chem. 1951, 190. [Google Scholar]
- Corrigan, J.J. D-Amino Acids in Animals. Science 1969, 164, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T. D-amino acids in higher plants. Life Sci. 1976, 19, 1097–1102. [Google Scholar] [CrossRef]
- Shapira, R.; Austin, G.E.; Mirra, S.S. Neuritic Plaque Amyloid in Alzheimer’s Disease Is Highly Racemized. J. Neurochem. 1988, 50, 69–74. [Google Scholar] [CrossRef]
- Man, E.H.; Sandhouse, M.E.; Burg, J.; Fisher, G.H. Accumulation of D-aspartic acid with age in the human brain. Science 1983, 220, 1407–1408. [Google Scholar] [CrossRef]
- Man, E.H.; Fisher, G.H.; Payan, I.L.; Cadilla-Perezrios, R.; Garcia, N.M.; Chemburkar, R.; Arends, G.; Frey, W.H. d-Aspartate in Human Brain. J. Neurochem. 1987, 48, 510–515. [Google Scholar] [CrossRef]
- Nagata, Y.; Borghi, M.; Fisher, G.H.; D’Aniello, A. Free d-serine concentration in normal and Alzheimer human brain. Brain Res. Bull. 1995, 38, 181–183. [Google Scholar] [CrossRef]
- Fadda, E.; Danysz, W.; Wroblewski, J.; Costa, E. Glycine and D-serine increase the affinity of N-methyl-D-aspartate sensitive glutamate binding sites in rat brain synaptic membranes. Neuropharmacology 1988, 27, 1183–1185. [Google Scholar] [CrossRef]
- Errico, F.; Mothet, J.-P.; Usiello, A. d-Aspartate: An endogenous NMDA receptor agonist enriched in the developing brain with potential involvement in schizophrenia. J. Pharm. Biomed. Anal. 2015, 116, 7–17. [Google Scholar] [CrossRef]
- Mothet, J.P.; Parent, A.T.; Wolosker, H.; Brady, R.O., Jr.; Linden, D.J.; Ferris, C.D.; Rogawski, M.A.; Snyder, S.H. D-Serine is an endogenous ligand for the glycine site of the N-methyl-D-aspartate receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 4926–4931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, M. Origin, Microbiology, Nutrition, and Pharmacology of D-Amino Acids. Chem. Biodivers. 2010, 7, 1491–1530. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, A.; Vetere, A.; Fisher, G.H.; Cusano, G.; Chavez, M.; Petrucelli, L. Presence of d-alanine in proteins of normal and Alzheimer human brain. Brain Res. 1992, 592, 44–48. [Google Scholar] [CrossRef]
- Chang, C.-H.; Lin, C.-H.; Lane, H.-Y. d-glutamate and Gut Microbiota in Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, S.A.; Berger, R.; Klomp, L.W.; De Koning, T.J. d-Amino acids in the central nervous system in health and disease. Mol. Genet. Metab. 2005, 85, 168–180. [Google Scholar] [CrossRef]
- Topo, E.; Soricelli, A.; Di Maio, A.; D’Aniello, E.; Di Fiore, M.M.; D’Aniello, A. Evidence for the involvement of d-aspartic acid in learning and memory of rat. Amino Acids 2009, 38, 1561–1569. [Google Scholar] [CrossRef]
- Papouin, T.; Ladépêche, L.; Ruel, J.; Sacchi, S.; Labasque, M.; Hanini, M.; Groc, L.; Pollegioni, L.; Mothet, J.-P.; Oliet, S.H. Synaptic and Extrasynaptic NMDA Receptors Are Gated by Different Endogenous Coagonists. Cell 2012, 150, 633–646. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, D.; Artoul, S.; Segal, A.C.; Kolodney, G.; Radzishevsky, I.; Dikopoltsev, E.; Foltyn, V.N.; Inoue, R.; Mori, H.; Billard, J.-M.; et al. Neuronal d-Serine and Glycine Release Via the Asc-1 Transporter Regulates NMDA Receptor-Dependent Synaptic Activity. J. Neurosci. 2013, 33, 3533–3544. [Google Scholar] [CrossRef] [Green Version]
- Holden, K.; Hartman, A. d-Leucine: Evaluation in an epilepsy model. Epilepsy Behav. 2017, 78, 202–209. [Google Scholar] [CrossRef]
- Temperini, C.; Scozzafava, A.; Vullo, D.; Supuran, C.T. Carbonic Anhydrase Activators. Activation of Isoforms I, II, IV, VA, VII, and XIV withl- andd-Phenylalanine and Crystallographic Analysis of Their Adducts with Isozyme II: Stereospecific Recognition within the Active Site of an Enzyme and Its Consequences for the Drug Design. J. Med. Chem. 2006, 49, 3019–3027. [Google Scholar] [CrossRef]
- Luengo, J.G.; Muñoz, M.-D.; Álvarez-Merz, I.; Herranz, A.S.; González, J.C.; Del Río, R.M.; Hernández-Guijo, J.M.; Solís, J.M. Intracellular accumulation of amino acids increases synaptic potentials in rat hippocampal slices. Amino Acids 2019, 51, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jung, H.; Kim, K.; Lim, K.-M.; Kim, J.-Y.; Jho, E.-H.; Oh, E.-S. D-tyrosine negatively regulates melanin synthesis by competitively inhibiting tyrosinase activity. Pigment. Cell Melanoma Res. 2017, 31, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Meloni, B.P.; Milani, D.; Edwards, A.B.; Anderton, R.S.; Doig, R.L.O.; Fitzgerald, M.; Palmer, T.N.; Knuckey, N.W. Neuroprotective peptides fused to arginine-rich cell penetrating peptides: Neuroprotective mechanism likely mediated by peptide endocytic properties. Pharmacol. Ther. 2015, 153, 36–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-L.; Niu, Y.-Y.; Jiang, W.-Z.; Tang, H.-L.; Zhang, C.; Xia, Q.-M.; Tang, X.-Q. Neuroprotective effects of hydrogen sulfide and the underlying signaling pathways. Rev. Neurosci. 2015, 26. [Google Scholar] [CrossRef] [PubMed]
- Mostyn, S.N.; Rawling, T.; Mohammadi, S.; Shimmon, S.; Frangos, Z.J.; Sarker, S.; Yousuf, A.; Vetter, I.; Ryan, R.M.; Christie, M.J.; et al. Development of an N-Acyl Amino Acid That Selectively Inhibits the Glycine Transporter 2 To Produce Analgesia in a Rat Model of Chronic Pain. J. Med. Chem. 2019, 62, 2466–2484. [Google Scholar] [CrossRef]
- Verger, A.; Stoffels, G.; Galldiks, N.; Lohmann, P.; Willuweit, A.; Neumaier, B.; Geisler, S.; Langen, K.-J. Investigation of cis-4-[18F]Fluoro-D-Proline Uptake in Human Brain Tumors After Multimodal Treatment. Mol. Imaging Boil. 2018, 20, 1035–1043. [Google Scholar] [CrossRef]
- Araújo, T.D.S.; Filho, A.J.M.C.; Monte, A.S.; Queiroz, A.I.D.G.; Cordeiro, R.C.; Machado, M.D.J.S.; Lima, R.F.; De Lucena, D.F.; Maes, M.; Macedo, D.S. Reversal of schizophrenia-like symptoms and immune alterations in mice by immunomodulatory drugs. J. Psychiatr. Res. 2017, 84, 49–58. [Google Scholar] [CrossRef]
- Koga, R.; Miyoshi, Y.; Sakaue, H.; Hamase, K.; Konno, R. Mouse d-Amino-Acid Oxidase: Distribution and Physiological Substrates. Front. Mol. Biosci. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Popiolek, M.; Tierney, B.; Steyn, S.J.; De Vivo, M. Lack of Effect of Sodium Benzoate at Reported Clinical Therapeutic Concentration on d-Alanine Metabolism in Dogs. ACS Chem. Neurosci. 2018, 9, 2832–2837. [Google Scholar] [CrossRef]
- Rojas, C.; Wilmoth, H.; Rais, R.; Hin, N.; DeVivo, M.; Popiolek, M.; Alt, J.; Ator, N.A.; Tsukamoto, T.; Slusher, B.S. Oral administration of D-alanine in monkeys robustly increases plasma and cerebrospinal fluid levels but experimental D-amino acid oxidase inhibitors had minimal effect. J. Psychopharmacol. 2016, 30, 887–895. [Google Scholar] [CrossRef]
- Lee, C.J.; Qiu, T.A.; Sweedler, J.V. d-Alanine: Distribution, origin, physiological relevance, and implications in disease. Biochimica Biophysica Acta Proteins Proteom. 2020, 1868. [Google Scholar] [CrossRef] [PubMed]
- Tsai, G.E.; Yang, P.; Chang, Y.-C.; Chong, M.-Y. D-Alanine Added to Antipsychotics for the Treatment of Schizophrenia. Boil. Psychiatry 2006, 59, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Yang, H.-T.; Lane, H.-Y. D-glutamate, D-serine, and D-alanine differ in their roles in cognitive decline in patients with Alzheimer’s disease or mild cognitive impairment. Pharmacol. Biochem. Behav. 2019, 185, 172760. [Google Scholar] [CrossRef]
- Ito, T.; Hayashida, M.; Kobayashi, S.; Muto, N.; Hayashi, A.; Yoshimura, T.; Mori, H. Serine racemase is involved in d-aspartate biosynthesis. J. Biochem. 2016, 160, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Puggioni, V.; Savinelli, A.; Miceli, M.; Molla, G.; Pollegioni, L.; Sacchi, S. Biochemical characterization of mouse d-aspartate oxidase. Biochimica Biophysica Acta Proteins Proteom. 2020, 1868. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, S.; De Novellis, V.; Paolone, G.; Nuzzo, T.; Iannotta, M.; Belardo, C.; Squillace, M.; Bolognesi, P.; Rosini, E.; Motta, Z.; et al. Olanzapine, but not clozapine, increases glutamate release in the prefrontal cortex of freely moving mice by inhibiting D-aspartate oxidase activity. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Errico, F.; Nisticò, R.; Di Giorgio, A.; Squillace, M.; Vitucci, D.; Galbusera, A.; Piccinin, S.; Mango, D.; Fazio, L.; Middei, S.; et al. Free D-aspartate regulates neuronal dendritic morphology, synaptic plasticity, gray matter volume and brain activity in mammals. Transl. Psychiatry 2014, 4, e417. [Google Scholar] [CrossRef] [PubMed]
- Punzo, D.; Errico, F.; Cristino, L.; Sacchi, S.; Keller, S.; Belardo, C.; Luongo, L.; Nuzzo, T.; Imperatore, R.; Florio, E.; et al. Age-Related Changes in D-Aspartate Oxidase Promoter Methylation Control Extracellular D-Aspartate Levels and Prevent Precocious Cell Death during Brain Aging. J. Neurosci. 2016, 36, 3064–3078. [Google Scholar] [CrossRef]
- Errico, F.; Napolitano, F.; Squillace, M.; Vitucci, D.; Blasi, G.; De Bartolomeis, A.; Bertolino, A.; D’Aniello, A.; Usiello, A. Decreased levels of d-aspartate and NMDA in the prefrontal cortex and striatum of patients with schizophrenia. J. Psychiatr. Res. 2013, 47, 1432–1437. [Google Scholar] [CrossRef]
- Lin, C.-H.; Yang, H.-T.; Chiu, C.-C.; Lane, H.-Y. Blood levels of D-amino acid oxidase vs. D-amino acids in reflecting cognitive aging. Sci. Rep. 2017, 7, 14849. [Google Scholar] [CrossRef]
- Wong, D.; Atiya, S.; Fogarty, J.; Montero-Odasso, M.; Pasternak, S.H.; Brymer, C.; Borrie, M.J.; Bartha, R. Reduced Hippocampal Glutamate and Posterior Cingulate N-Acetyl Aspartate in Mild Cognitive Impairment and Alzheimer’s Disease Is Associated with Episodic Memory Performance and White Matter Integrity in the Cingulum: A Pilot Study. J. Alzheimer’s Dis. 2020, 73, 1385–1405. [Google Scholar] [CrossRef] [PubMed]
- Murtas, G.; Sacchi, S.; Valentino, M.; Pollegioni, L. Biochemical Properties of Human D-Amino Acid Oxidase. Front. Mol. Biosci. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balu, D.T.; Pantazopoulos, H.; Huang, C.C.; Muszynski, K.; Harvey, T.L.; Uno, Y.; Rorabaugh, J.M.; Galloway, C.R.; Botz-Zapp, C.; Berretta, S.; et al. Neurotoxic astrocytes express the d-serine synthesizing enzyme, serine racemase, in Alzheimer’s disease. Neurobiol. Dis. 2019, 130, 104511. [Google Scholar] [CrossRef] [PubMed]
- Moaddel, R.; Luckenbaugh, D.A.; Xie, Y.; Villaseñor, A.; Brutsche, N.E.; Machado-Vieira, R.; Ramamoorthy, A.; Lorenzo, M.P.; Garcia, A.; Bernier, M.; et al. D-serine plasma concentration is a potential biomarker of (R,S)-ketamine antidepressant response in subjects with treatment-resistant depression. Psychopharmacology 2014, 232, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Mackay, M.-A.B.; Kravtsenyuk, M.; Thomas, R.; Mitchell, N.D.; Dursun, S.M.; Baker, G.B. D-Serine: Potential Therapeutic Agent and/or Biomarker in Schizophrenia and Depression? Front. Psychiatry 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papouin, T.; Haydon, P.G. D-serine Measurements in Brain Slices or Other Tissue Explants. Bio-protocol 2018, 8, e2698. [Google Scholar] [CrossRef] [PubMed]
- Billard, J.-M. d-Serine in the aging hippocampus. J. Pharm. Biomed. Anal. 2015, 116, 18–24. [Google Scholar] [CrossRef]
- Li, S.; Uno, Y.; Rudolph, U.; Cobb, J.; Liu, J.; Anderson, T.; Levy, D.; Balu, D.T.; Coyle, J.T. Astrocytes in primary cultures express serine racemase, synthesize d -serine and acquire A1 reactive astrocyte features. Biochem. Pharmacol. 2018, 151, 245–251. [Google Scholar] [CrossRef]
- Lam, H.; Oh, N.-C.; Cava, F.; Takacs, C.N.; Clardy, J.; De Pedro, M.A.; Waldor, M.K. D-Amino Acids Govern Stationary Phase Cell Wall Remodeling in Bacteria. Science 2009, 325, 1552–1555. [Google Scholar] [CrossRef] [Green Version]
- Mesuret, G.; Khabbazzadeh, S.; Bischoff, A.M.; Safory, H.; Wolosker, H.; Hülsmann, S. A neuronal role of the Alanine-Serine-Cysteine-1 transporter (SLC7A10, Asc-1) for glycine inhibitory transmission and respiratory pattern. Sci. Rep. 2018, 8, 8536. [Google Scholar] [CrossRef]
- Billard, J.-M.; Freret, T. Asc-1 transporter activation: An alternative to rescue age-related alterations in functional plasticity at rat hippocampal CA3/CA1 synapses. J. Neurochem. 2018, 147, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamase, K.; Inoue, T.; Morikawa, A.; Konno, R.; Zaitsu, K. Determination of Free -Proline and -Leucine in the Brains of Mutant Mice Lacking -Amino Acid Oxidase Activity. Anal. Biochem. 2001, 298, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Mutaguchi, Y.; Ohmori, T.; Wakamatsu, T.; Doi, K.; Ohshima, T. Identification, Purification, and Characterization of a Novel Amino Acid Racemase, Isoleucine 2-Epimerase, from Lactobacillus Species. J. Bacteriol. 2013, 195, 5207–5215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, A.; Santos, P.; O’Riordan, K.J.; Stafstrom, C.E.; Hardwick, J.M. Potent anti-seizure effects of D-leucine. Neurobiol. Dis. 2015, 82, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.D.; Costa, A.; Rani, B.; Nachtigall, E.G.; Passani, M.B.; Carta, F.; Nocentini, A.; Myskiw, J.D.C.; Furini, C.R.G.; Supuran, C.T.; et al. The role of carbonic anhydrases in extinction of contextual fear memory. Proc. Natl. Acad. Sci. USA 2020, 117, 16000–16008. [Google Scholar] [CrossRef]
- Park, J.; Jung, H.; Jang, B.; Song, H.-K.; Han, I.-O.; Oh, E.-S. D-tyrosine adds an anti-melanogenic effect to cosmetic peptides. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Dang, S.; Wang, X.; Zhang, J.; Zhang, L.; Su, Q.; Zhang, H.; Lin, T.; Zhang, X.; Zhang, Y.; et al. NO involvement in the inhibition of ghrelin on voltage-dependent potassium currents in rat hippocampal cells. Brain Res. 2018, 1678, 40–46. [Google Scholar] [CrossRef]
- Milani, D.; Knuckey, N.W.; Anderton, R.S.; Cross, J.L.; Meloni, B.P. The R18 Polyarginine Peptide Is More Effective Than the TAT-NR2B9c (NA-1) Peptide When Administered 60 Minutes after Permanent Middle Cerebral Artery Occlusion in the Rat. Stroke Res. Treat. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Milani, D.; Bakeberg, M.C.; Cross, J.L.; Clark, V.W.; Anderton, R.S.; Blacker, D.J.; Knuckey, N.W.; Meloni, B.P. Comparison of neuroprotective efficacy of poly-arginine R18 and R18D (D-enantiomer) peptides following permanent middle cerebral artery occlusion in the Wistar rat and in vitro toxicity studies. PLoS ONE 2018, 13, e0193884. [Google Scholar] [CrossRef]
- Edwards, A.B.; Anderton, R.S.; Knuckey, N.W.; Meloni, B.P. Assessment of therapeutic window for poly-arginine-18D (R18D) in a P7 rat model of perinatal hypoxic-ischaemic encephalopathy. J. Neurosci. Res. 2018, 96, 1816–1826. [Google Scholar] [CrossRef]
- Edwards, A.B.; Cross, J.L.; Anderton, R.S.; Knuckey, N.W.; Meloni, B.P. Poly-arginine R18 and R18D (D-enantiomer) peptides reduce infarct volume and improves behavioural outcomes following perinatal hypoxic-ischaemic encephalopathy in the P7 rat. Mol. Brain 2018, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Milani, D.; Clark, V.; Cross, J.; Anderton, R.; Knuckey, N.; Meloni, B. Poly-arginine peptides reduce infarct volume in a permanent middle cerebral artery rat stroke model. BMC Neurosci. 2016, 17, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, A.B.; Anderton, R.S.; Knuckey, N.W.; Meloni, B.P. Characterisation of neuroprotective efficacy of modified poly-arginine-9 (R9) peptides using a neuronal glutamic acid excitotoxicity model. Mol. Cell. Biochem. 2016, 426, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Seki, T. Availability of D-cysteine as a protectant for cerebellar neurons. Nihon Yakurigaku Zasshi 2019, 154, 133–137. [Google Scholar] [CrossRef]
- Shibuya, N.; Koike, S.; Tanaka, M.; Ishigami-Yuasa, M.; Kimura, Y.; Ogasawara, Y.; Fukui, K.; Nagahara, N.; Kimura, H. A novel pathway for the production of hydrogen sulfide from D-cysteine in mammalian cells. Nat. Commun. 2013, 4, 1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleixner, A.M.; Hutchison, D.F.; Sannino, S.; Bhatia, T.N.; Leak, L.C.; Flaherty, P.T.; Wipf, P.; Brodsky, J.L.; Leak, R.K. N-Acetyl-l-Cysteine Protects Astrocytes against Proteotoxicity without Recourse to Glutathione. Mol. Pharmacol. 2017, 92, 564–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altawallbeh, G.; Smith, L.; Lewis, S.J.; Authier, S.; Bujold, K.; Gaston, B.; Bederman, I. Pharmacokinetic study of Sudaxine in dog plasma using novel LC–MS/MS method. Drug Test. Anal. 2018, 11, 403–410. [Google Scholar] [CrossRef]
- Yagi, K.; Okamura, K.; Naoi, M.; Takai, A.; Kotaki, A. Reaction of D-amino-acid oxidase with D-lysine. J. Biochem. 1969, 66, 581–589. [Google Scholar] [CrossRef]
- Mostyn, S.N.; Wilson, K.A.; Schumann-Gillett, A.; Frangos, Z.J.; Shimmon, S.; Rawling, T.; Ryan, R.M.; O’Mara, M.L.; Vandenberg, R.J. Identification of an allosteric binding site on the human glycine transporter, GlyT2, for bioactive lipid analgesics. Elife 2019, 8. [Google Scholar] [CrossRef]
- Gallagher, C.I.; Sheipouri, D.; Shimmon, S.; Rawling, T.; Vandenberg, R.J. Identification of N-acyl amino acids that are positive allosteric modulators of glycine receptors. Biochem. Pharmacol. 2020, 180, 114117. [Google Scholar] [CrossRef]
- Hinduja, S.; Kraus, K.S.; Manohar, S.; Salvi, R.J. D-methionine protects against cisplatin-induced neurotoxicity in the hippocampus of the adult rat. Neurotox. Res. 2014, 27, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Mizutani, A.; Nishi, K.; Nakajima, S.; Shikano, N.; Nishii, R.; Fukuchi, K.; Kawai, K. Differences in accumulation and the transport mechanism of l- and d-methionine in high- and low-grade human glioma cells. Nucl. Med. Boil. 2017, 44, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Fox, D.J.; Cooper, M.D.; Speil, C.A.; Roberts, M.H.; Yanik, S.C.; Meech, R.P.; Hargrove, T.L.; Verhulst, S.J.; Rybak, L.P.; Campbell, K.C.M. d-Methionine reduces tobramycin-induced ototoxicity without antimicrobial interference in animal models. J. Cyst. Fibros. 2015, 15, 518–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, K.C.M.; Martin, S.M.; Meech, R.P.; Hargrove, T.L.; Verhulst, S.J.; Fox, D.J. D-methionine (D-met) significantly reduces kanamycin-induced ototoxicity in pigmented guinea pigs. Int. J. Audiol. 2016, 55, 1–6. [Google Scholar] [CrossRef]
- Lopes-Azevedo, S.; Busnardo, C.; Corrêa, F.M.A. Central mechanism of the cardiovascular responses caused by L-proline microinjected into the paraventricular nucleus of the hypothalamus in unanesthetized rats. Brain Res. 2016, 1652, 43–52. [Google Scholar] [CrossRef]
- Sommerauer, M.; Galldiks, N.; Barbe, M.T.; Stoffels, G.; Willuweit, A.; Coenen, H.H.; Schroeter, M.; Timmermann, L.; Fink, G.R.; Langen, K.-J. Cis-4-[18F]fluoro-D-proline detects neurodegeneration in patients with akinetic-rigid parkinsonism. Nucl. Med. Commun. 2019, 40, 383–387. [Google Scholar] [CrossRef]
- Geisler, S.; Willuweit, A.; Schroeter, M.; Zilles, K.; Hamacher, K.; Galldiks, N.; Shah, N.J.; Coenen, H.H.; Langen, K.-J. Detection of remote neuronal reactions in the Thalamus and Hippocampus induced by rat glioma using the PET tracer cis-4-[18F]fluoro-D-proline. J. Cereb. Blood Flow Metab. 2013, 33, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Geisler, S.; Ermert, J.; Stoffels, G.; Willuweit, A.; Galldiks, N.; Filss, C.; Shah, N.J.; Coenen, H.H.; Langen, K.-J. Isomers of 4-[18F]fluoro-proline: Radiosynthesis, biological evaluation and results in humans using PET. Curr. Radiopharm. 2014, 7, 123–132. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Cheng, Y.-S.; Liao, T.-Y.; Lin, C.; Chen, Z.-T.; Twu, W.-I.; Chang, C.-W.; Tan, D.T.-W.; Liu, R.-S.; Tu, P.; et al. Intranasal Administration of a Polyethylenimine-Conjugated Scavenger Peptide Reduces Amyloid-β Accumulation in a Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 53, 1053–1067. [Google Scholar] [CrossRef]
- Notarangelo, F.M.; Wang, X.; Horning, K.J.; Schwarcz, R. Role of d-amino acid oxidase in the production of kynurenine pathway metabolites from d-tryptophan in mice. J. Neurochem. 2016, 136, 804–814. [Google Scholar] [CrossRef] [Green Version]
- Asaoka, N.; Kawai, H.; Nishitani, N.; Kinoshita, H.; Shibui, N.; Nagayasu, K.; Shirakawa, H.; Kaneko, S. A new designer drug 5F-ADB activates midbrain dopaminergic neurons but not serotonergic neurons. J. Toxicol. Sci. 2016, 41, 813–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genchi, G. An overview on d-amino acids. Amino Acids 2017, 49, 1521–1533. [Google Scholar] [CrossRef] [PubMed]
- Coyle, J.T.; Balu, D.T. The Role of Serine Racemase in the Pathophysiology of Brain Disorders. Adv. Pharmacol. 2018, 82, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Aliashkevich, A.; Alvarez, L.; Cava, F. New Insights Into the Mechanisms and Biological Roles of D-Amino Acids in Complex Eco-Systems. Front. Microbiol. 2018, 9, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasabe, J.; Miyoshi, Y.; Rakoff-Nahoum, S.; Zhang, T.; Mita, M.; Davis, B.M.; Hamase, K.; Waldor, M.K. Interplay between microbial d-amino acids and host d-amino acid oxidase modifies murine mucosal defence and gut microbiota. Nat. Microbiol. 2016, 1, 16125. [Google Scholar] [CrossRef] [PubMed]
- Bastings, J.J.; Van Eijk, H.M.; Damink, S.W.M.O.; Rensen, S.S. d-amino Acids in Health and Disease: A Focus on Cancer. Nutrients 2019, 11, 2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kepert, I.; Fonseca, J.; Müller, C.; Milger, K.; Hochwind, K.; Kostric, M.; Fedoseeva, M.; Ohnmacht, C.; Dehmel, S.; Nathan, P.; et al. D-tryptophan from probiotic bacteria influences the gut microbiome and allergic airway disease. J. Allergy Clin. Immunol. 2017, 139, 1525–1535. [Google Scholar] [CrossRef] [Green Version]
- Quigley, E.M. Microbiota-Brain-Gut Axis and Neurodegenerative Diseases. Curr. Neurol. Neurosci. Rep. 2017, 17. [Google Scholar] [CrossRef]
- Yarandi, S.S.; Peterson, D.A.; Treisman, G.J.; Moran, T.H.; Pasricha, P.J. Modulatory Effects of Gut Microbiota on the Central Nervous System: How Gut Could Play a Role in Neuropsychiatric Health and Diseases. J. Neurogastroenterol. Motil. 2016, 22, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, Z.-Q.; Shen, L.-L.; Li, W.-W.; Fu, X.; Zeng, F.; Gui, L.; Lu, Y.; Cai, M.; Zhu, C.; Tan, Y.-L.; et al. Gut Microbiota is Altered in Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 63, 1337–1346. [Google Scholar] [CrossRef] [Green Version]
- Brückner, H.; Westhauser, T. Chromatographic determination of L- and D-amino acids in plants. Amino Acids 2003, 24, 43–55. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, A.; Vetere, A.; Petrucelli, L. Further study on the specificity of d-amino acid oxidase and of d-aspartate oxidase and time course for complete oxidation of d-amino acids. Comp. Biochem. Physiol. Part. B: Comp. Biochem. 1993, 105, 731–734. [Google Scholar] [CrossRef]
- Pollegioni, L.; Butò, S.; Tischer, W.; Ghisla, S.; Pilone, M.S. Characterization of D-amino acid oxidase from Trigonopsis variabilis. Biochem. Mol. Boil. Int. 1993, 31. [Google Scholar]
- Kimura, T.; Hesaka, A.; Isaka, Y. d-Amino acids and kidney diseases. Clin. Exp. Nephrol. 2020, 24, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Ariyoshi, M.; Katane, M.; Hamase, K.; Miyoshi, Y.; Nakane, M.; Hoshino, A.; Okawa, Y.; Mita, Y.; Kaimoto, S.; Uchihashi, M.; et al. D-Glutamate is metabolized in the heart mitochondria. Sci. Rep. 2017, 7, 43911. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, T.; Sacchi, S.; Errico, F.; Keller, S.; Palumbo, O.; Florio, E.; Punzo, D.; Napolitano, F.; Copetti, M.; Carella, M.; et al. Decreased free d-aspartate levels are linked to enhanced d-aspartate oxidase activity in the dorsolateral prefrontal cortex of schizophrenia patients. Npj Schizophr. 2017, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vyklicky, V.; Korinek, M.; Smejkalova, T.; Balik, A.; Krausova, B.; Kaniakova, M.; Lichnerova, K.; Cerny, J.; Krusek, J.; Dittert, I.; et al. Structure, Function, and Pharmacology of NMDA Receptor Channels. Physiol. Res. 2014, S191–S203. [Google Scholar] [CrossRef]
- Tajima, N.; Karakas, E.; Grant, T.; Simorowski, N.; Diaz-Avalos, R.; Grigorieff, N.; Furukawa, H. Activation of NMDA receptors and the mechanism of inhibition by ifenprodil. Nat. 2016, 534, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chang, L.; Song, Y.; Li, H.; Wu, Y. The Role of NMDA Receptors in Alzheimer’s Disease. Front. Neurosci. 2019, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Balu, D.T. The NMDA Receptor and Schizophrenia: From Pathophysiology to Treatment. Adv. Pharmacol. 2016, 76, 351–382. [Google Scholar] [CrossRef] [Green Version]
- D’Aniello, A. d-Aspartic acid: An endogenous amino acid with an important neuroendocrine role. Brain Res. Rev. 2007, 53, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Still, J.L.; Buell, M.V. Studies on the cyclophorase system; D-aspartic oxidase. J. Boil. Chem. 1949, 179. [Google Scholar]
- Olverman, H.; Jones, A.; Mewett, K.; Watkins, J. Structure/activity relations of N-methyl-d-aspartate receptor ligands as studied by their inhibition of [3H]d2-amino-5-phosphonopentanoic acid binding in rat brain membranes. Neuroscience 1988, 26, 17–31. [Google Scholar] [CrossRef]
- Hashimoto, A.; Nishikawa, T.; Hayashi, T.; Fujii, N.; Harada, K.; Oka, T.; Takahashi, K. The presence of free D-serine in rat brain. FEBS Lett. 1992, 296, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Nagata, Y.; Horiike, K.; Maeda, T. Distribution of freed-serine in vertebrate brains. Brain Res. 1994, 634, 291–295. [Google Scholar] [CrossRef]
- Hashimoto, A.; Kumashiro, S.; Nishikawa, T.; Oka, T.; Takahashi, K.; Mito, T.; Takashima, S.; Doi, N.; Mizutani, Y.; Yamazaki, T.; et al. Embryonic Development and Postnatal Changes in Free d-Aspartate and d-Serine in the Human Prefrontal Cortex. J. Neurochem. 1993, 61, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Neame, S.; Safory, H.; Radzishevsky, I.; Touitou, A.; Marchesani, F.; Marchetti, M.; Kellner, S.; Berlin, S.; Foltyn, V.N.; Engelender, S.; et al. The NMDA receptor activation by d-serine and glycine is controlled by an astrocytic Phgdh-dependent serine shuttle. Proc. Natl. Acad. Sci. USA 2019, 116, 20736–20742. [Google Scholar] [CrossRef] [Green Version]
- Seif, T.; Simms, J.A.; Lei, K.; Wegner, S.; Bonci, A.; Messing, R.O.; Hopf, F.W. D-Serine and D-Cycloserine Reduce Compulsive Alcohol Intake in Rats. Neuropsychopharmacology 2015, 40, 2357–2367. [Google Scholar] [CrossRef] [Green Version]
- Wegner, S.A.; Hu, B.; Sergio, T.D.O.; Darevsky, D.; Kwok, C.C.-Y.; Lei, K.; Hopf, F.W. A novel NMDA receptor-based intervention to suppress compulsion-like alcohol drinking. Neuropharmacology 2019, 157, 107681. [Google Scholar] [CrossRef]
- Perez, E.J.; Tapanes, S.A.; Loris, Z.B.; Balu, D.T.; Sick, T.J.; Coyle, J.T.; Liebl, D.J. Enhanced astrocytic d-serine underlies synaptic damage after traumatic brain injury. J. Clin. Investig. 2017, 127, 3114–3125. [Google Scholar] [CrossRef]
- Biegon, A.; Liraz-Zaltsman, S.; Shohami, E. Stimulation of N-methyl-D-aspartate receptors by exogenous and endogenous ligands improves outcome of brain injury. Curr. Opin. Neurol. 2018, 31, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Seif, T.; Chang, S.-J.; Simms, J.A.; Gibb, S.L.; Dadgar, J.; Chen, B.T.; Harvey, B.K.; Ron, R.; Messing, R.O.; Bonci, A.; et al. Cortical activation of accumbens hyperpolarization-active NMDARs mediates aversion-resistant alcohol intake. Nat. Neurosci. 2013, 16, 1094–1100. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Punzo, D.; Cuomo, M.; Affinito, O.; Coretti, L.; Sacchi, S.; Florio, E.; Lembo, F.; Carella, M.; Copetti, M.; et al. DNA methylation landscape of the genes regulating D-serine and D-aspartate metabolism in post-mortem brain from controls and subjects with schizophrenia. Sci. Rep. 2018, 8, 10163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Tallawy, H.N.; Saleem, T.H.; El-Ebidi, A.M.; Hassan, M.H.; Gabra, R.H.; Farghaly, W.M.A.; El-Maali, N.A.; Sherkawy, H.S. Clinical and biochemical study of d-serine metabolism among schizophrenia patients. Neuropsychiatr. Dis. Treat. 2017, 13, 1057–1063. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, H.; Masuda, N.; Natori, H.; Shinohara, Y.; Ichida, K. Pharmacokinetics and toxicokinetics of d-serine in rats. J. Pharm. Biomed. Anal. 2019, 162, 264–271. [Google Scholar] [CrossRef]
- Cho, S.-E.; Na, K.-S.; Cho, S.-J.; Kang, S.-G. Low d-serine levels in schizophrenia: A systematic review and meta-analysis. Neurosci. Lett. 2016, 634, 42–51. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Malhotra, A.K.; Cornblatt, B.; Silipo, G.; Balla, A.; Suckow, R.F.; D’Souza, C.; Saksa, J.; Woods, S.W.; Javitt, D.C. High dose D-serine in the treatment of schizophrenia. Schizophr. Res. 2010, 121, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Kantrowitz, J.T.; Woods, S.W.; Petkova, E.; Cornblatt, B.; Corcoran, C.M.; Chen, H.; Silipo, G.; Javitt, D.C. D-serine for the treatment of negative symptoms in individuals at clinical high risk of schizophrenia: A pilot, double-blind, placebo-controlled, randomised parallel group mechanistic proof-of-concept trial. Lancet Psychiatry 2015, 2, 403–412. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Epstein, M.L.; Lee, M.; Lehrfeld, N.; Nolan, K.; Shope, C.; Petkova, E.; Silipo, G.; Javitt, D.C. Improvement in mismatch negativity generation during d -serine treatment in schizophrenia: Correlation with symptoms. Schizophr. Res. 2018, 191, 70–79. [Google Scholar] [CrossRef]
- Beltrán-Castillo, S.; Eugenín, J.; Von Bernhardi, R. Impact of Aging in Microglia-Mediated D-Serine Balance in the CNS. Mediat. Inflamm. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Nuzzo, T.; Punzo, D.; Devoto, P.; Rosini, E.; Paciotti, S.; Sacchi, S.; Li, Q.; Thiolat, M.-L.; Véga, C.; Carella, M.; et al. The levels of the NMDA receptor co-agonist D-serine are reduced in the substantia nigra of MPTP-lesioned macaques and in the cerebrospinal fluid of Parkinson’s disease patients. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talevi, A.; Enrique, A.V.; Bruno-Blanch, L.E. Anticonvulsant activity of artificial sweeteners: A structural link between sweet-taste receptor T1R3 and brain glutamate receptors. Bioorgan. Med. Chem. Lett. 2012, 22, 4072–4074. [Google Scholar] [CrossRef] [PubMed]
- Canto-De-Souza, L.; Provensi, G.; Vullo, D.; Carta, F.; Scozzafava, A.; Costa, A.; Schmidt, S.D.; Passani, M.B.; Supuran, C.T.; Blandina, P. Carbonic anhydrase activation enhances object recognition memory in mice through phosphorylation of the extracellular signal-regulated kinase in the cortex and the hippocampus. Neuropharmacology 2017, 118, 148–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galarreta, M.; Bustamante, J.; Del Río, R.M.; Solís, J.M. Taurine induces a long-lasting increase of synaptic efficacy and axon excitability in the hippocampus. J. Neurosci. 1996, 16, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Chepkova, A.N.; Sergeeva, O.A.; Haas, H.L. Taurine rescues hippocampal long-term potentiation from ammonia-induced impairment. Neurobiol. Dis. 2006, 23, 512–521. [Google Scholar] [CrossRef]
- Carballo-Carbajal, I.; Laguna, A.; Romero-Giménez, J.; Cuadros, T.; Bové, J.; Martinez-Vicente, M.; Parent, A.; Gonzalez-Sepúlveda, M.; Peñuelas, N.; Torra, A.; et al. Brain tyrosinase overexpression implicates age-dependent neuromelanin production in Parkinson’s disease pathogenesis. Nat. Commun. 2019, 10, 973. [Google Scholar] [CrossRef] [Green Version]
- Greggio, E.; Bergantino, E.; Carter, D.; Ahmad, R.; Costin, G.-E.; Hearing, V.J.; Clarimón, J.; Singleton, A.; Eerola, J.; Hellström, O.; et al. Tyrosinase exacerbates dopamine toxicity but is not genetically associated with Parkinson’s disease. J. Neurochem. 2005, 93, 246–256. [Google Scholar] [CrossRef]
- Tessari, I.; Bisaglia, M.; Valle, F.; Samorì, B.; Bergantino, E.; Mammi, S.; Bubacco, L.; Samorǐ, B. The Reaction of α-Synuclein with Tyrosinase. J. Boil. Chem. 2008, 283, 16808–16817. [Google Scholar] [CrossRef] [Green Version]
- Milani, D.; Cross, J.L.; Anderton, R.S.; Blacker, D.J.; Knuckey, N.W.; Meloni, B.P. Neuroprotective efficacy of poly-arginine R18 and NA-1 (TAT-NR2B9c) peptides following transient middle cerebral artery occlusion in the rat. Neurosci. Res. 2017, 114, 9–15. [Google Scholar] [CrossRef]
- Meloni, B.P.; Brookes, L.; Clark, V.W.; Cross, J.L.; Edwards, A.B.; Anderton, R.S.; Hopkins, R.M.; Hoffmann, K.; Knuckey, N.W. Poly-Arginine and Arginine-Rich Peptides are Neuroprotective in Stroke Models. J. Cereb. Blood Flow Metab. 2015, 35, 993–1004. [Google Scholar] [CrossRef]
- Meloni, B.P.; Milani, D.; Cross, J.L.; Clark, V.W.; Edwards, A.B.; Anderton, R.S.; Blacker, D.J.; Knuckey, N.W. Assessment of the Neuroprotective Effects of Arginine-Rich Protamine Peptides, Poly-Arginine Peptides (R12-Cyclic, R22) and Arginine–Tryptophan-Containing Peptides Following In Vitro Excitotoxicity and/or Permanent Middle Cerebral Artery Occlusion in Rats. Neuromol. Med. 2017, 19, 271–285. [Google Scholar] [CrossRef]
- Liddle, L.; Reinders, R.; South, S.; Blacker, D.; Knuckey, N.; Colbourne, F.; Meloni, B. Poly-arginine-18 peptides do not exacerbate bleeding, or improve functional outcomes following collagenase-induced intracerebral hemorrhage in the rat. PLoS ONE 2019, 14, e0224870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenna, J.E.; Anderton, R.S.; Knuckey, N.W.; Meloni, B.P. Assessment of recombinant tissue plasminogen activator (rtPA) toxicity in cultured neural cells and subsequent treatment with poly-arginine peptide R18D. Neurochem. Res. 2020, 45, 1215–1229. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.B.; Anderton, R.S.; Knuckey, N.W.; Meloni, B.P. Perinatal Hypoxic-Ischemic Encephalopathy and Neuroprotective Peptide Therapies: A Case for Cationic Arginine-Rich Peptides (CARPs). Brain Sci. 2018, 8, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meloni, B.P.; South, S.M.; Gill, D.A.; Marriott, A.L.; Déziel, R.A.; Jacques, A.; Blacker, D.J.; Knuckey, N.W. Poly-Arginine Peptides R18 and R18D Improve Functional Outcomes After Endothelin-1-Induced Stroke in the Sprague Dawley Rat. J. Neuropathol. Exp. Neurol. 2019, 78, 426–435. [Google Scholar] [CrossRef] [PubMed]
- MacDougall, G.; Anderton, R.S.; Edwards, A.B.; Knuckey, N.W.; Meloni, B.P. The Neuroprotective Peptide Poly-Arginine-12 (R12) Reduces Cell Surface Levels of NMDA NR2B Receptor Subunit in Cortical Neurons; Investigation into the Involvement of Endocytic Mechanisms. J. Mol. Neurosci. 2016, 61, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.S.; Anderton, R.S.; Cross, J.L.; Clark, V.W.; Edwards, A.B.; Knuckey, N.W.; Meloni, B.P. Assessment of R18, COG1410, and APP96-110 in excitotoxicity and traumatic brain injury. Transl. Neurosci. 2017, 8, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.; Wong, K.Y.; Rupasinghe, C.N.; Tiwari, R.; Zhao, X.; Berberoglu, E.D.; Sinkler, C.; Liu, J.; Lee, I.; Parang, K.; et al. Inhibition ofN-Methyl-d-aspartate-induced Retinal Neuronal Death by Polyarginine Peptides Is Linked to the Attenuation of Stress-induced Hyperpolarization of the Inner Mitochondrial Membrane Potential. J. Boil. Chem. 2015, 290, 22030–22048. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; Alavi, M.V.; Kim, K.-Y.; Kang, T.; Scott, R.T.; Noh, Y.H.; Lindsey, J.D.; Wissinger, B.; Ellisman, M.H.; Weinreb, R.N.; et al. A new vicious cycle involving glutamate excitotoxicity, oxidative stress and mitochondrial dynamics. Cell Death Dis. 2011, 2, e240. [Google Scholar] [CrossRef]
- Kown, M.H.; Lijkwan, M.A.; Jahncke, C.L.; Murata, S.; Rothbard, J.B.; Robbins, R.C. L-arginine polymers enhance coronary flow and reduce oxidative stress following cardiac transplantation in rats. J. Thorac. Cardiovasc. Surg. 2003, 126, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Narasimhulu, C.A.; Selvarajan, K.; Brown, M.; Parthasarathy, S. Cationic peptides neutralize Ox-LDL, prevent its uptake by macrophages, and attenuate inflammatory response. Atherosclerosis 2014, 236, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Won, Y.-W.; Kim, H.A.; Lee, M.; Kim, Y.-H. Reducible Poly(oligo-D-arginine) for Enhanced Gene Expression in Mouse Lung by Intratracheal Injection. Mol. Ther. 2010, 18, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Hyun, H.; Won, Y.-W.; Kim, K.-M.; Lee, J.; Lee, M.; Kim, Y.-H. Therapeutic effects of a reducible poly (oligo-d-arginine) carrier with the heme oxygenase-1 gene in the treatment of hypoxic-ischemic brain injury. Biomaterials 2010, 31, 9128–9134. [Google Scholar] [CrossRef] [PubMed]
- Won, Y.-W.; Kim, K.-M.; An, S.S.; Lee, M.; Ha, Y.; Kim, Y.-H. Suicide gene therapy using reducible poly (oligo-d-arginine) for the treatment of spinal cord tumors. Biomaterials 2011, 32, 9766–9775. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.; Bae, S.-H.; Kim, B.; Park, J.S.; Jung, S.; Lee, M.; Kim, Y.-H.; Choi, D. Cardiac Usage of Reducible Poly(oligo-D-arginine) As a Gene Carrier for Vascular Endothelial Growth Factor Expression. PLoS ONE 2015, 10, e0144491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, H. Physiological role of hydrogen sulfide and polysulfide in the central nervous system. Neurochem. Int. 2013, 63, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Sato, M.; Konno, A.; Hirai, H.; Kurauchi, Y.; Hisatsune, A.; Katsuki, H. d-Cysteine promotes dendritic development in primary cultured cerebellar Purkinje cells via hydrogen sulfide production. Mol. Cell. Neurosci. 2018, 93, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Borowicz, K.K.; Swiader, M.; Kamiński, R.; Kuźniar, H.; Kleinrok, Z.; Czuczwar, S.J. Two essential amino acids, L-lysine and L-histidine, in five types of experimental seizures. Pol. J. Pharmacol. 2001, 52. [Google Scholar]
- Ikeda, Y.; Mochizuki, Y.; Matsumoto, H.; Nakamura, Y.; Dohi, K.; Jimbo, H.; Shimazu, M.; Hayashi, M.; Matsumoto, K. L-histidine but not D-histidine attenuates brain edema following cryogenic injury in rats. Acta Neurochirurgica Suppl. 2000, 76, 195–197. [Google Scholar] [CrossRef]
- Yamakami, J.; Sakurai, E.; Sakurada, T.; Maeda, K.; Hikichi, N. Stereoselective blood-brain barrier transport of histidine in rats. Brain Res. 1998, 812, 105–112. [Google Scholar] [CrossRef]
- Torii, K.; Niijima, A. Effect of lysine on afferent activity of the hepatic branch of the vagus nerve in normal and l-lysine-deficient rats. Physiol. Behav. 2001, 72, 685–690. [Google Scholar] [CrossRef]
- Grove, J.; Henderson, L. The metabolism of d- and l-lysine in the intact rat, perfused liver and liver mitochondria. Biochimica Biophysica Acta 1968, 165, 113–120. [Google Scholar] [CrossRef]
- Vandenberg, R.J.; Ryan, R.M.; Carland, J.E.; Imlach, W.; Christie, M.J. Glycine transport inhibitors for the treatment of pain. Trends Pharmacol. Sci. 2014, 35, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Glycine Transport Inhibitors for the Treatment of Schizophrenia. Open Med. Chem. J. 2010, 4, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Claussen, A.D.; Fox, D.J.; Yu, X.C.; Meech, R.P.; Verhulst, S.J.; Hargrove, T.L.; Campbell, K.C.M. D-methionine pre-loading reduces both noise-induced permanent threshold shift and outer hair cell loss in the chinchilla. Int. J. Audiol. 2013, 52, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Gopal, K.V.; Moore, E.J.; Gross, G.W. Antioxidants l-carnitine and d-methionine modulate neuronal activity through GABAergic inhibition. J. Neural Transm. 2014, 121, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qu, Y.; Chen, X.; Zhang, P.; Su, D.; Wang, L.; Yang, F.; Yang, J. Effects of D-methionine in mice with noise-induced hearing loss mice. J. Int. Med. Res. 2019, 47, 3874–3885. [Google Scholar] [CrossRef] [Green Version]
- Garweg, G.; Dahnke, H.G. Is the conversion of D-proline into L-proline in the brain a prerequisite for the incorporation into nerve-cell proteins? Verh Anat Ges. 1974, 68, 375–379. [Google Scholar]
- Langen, K.-J.; Hamacher, K.; Bauer, D.; Bröer, S.; Pauleit, D.; Herzog, H.; Floeth, F.; Zilles, K.; Coenen, H.H. Preferred Stereoselective Transport of the D-isomer ofcis-4-[18F]fluoro-proline at the Blood–Brain Barrier. J. Cereb. Blood Flow Metab. 2005, 25, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Langen, K.-J.; Salber, D.; Hamacher, K.; Stoffels, G.; Reifenberger, G.; Pauleit, D.; Coenen, H.H.; Zilles, K. Detection of Secondary Thalamic Degeneration After Cortical Infarction Using cis-4-18F-Fluoro- D-Proline. J. Nucl. Med. 2007, 48, 1482–1491. [Google Scholar] [CrossRef] [Green Version]
- Perkins, M.; Stone, T.W. An iontophoretic investigation of the actions of convulsant kynurenines and their interaction with the endogenous excitant quinolinic acid. Brain Res. 1982, 247, 184–187. [Google Scholar] [CrossRef]
- Kessler, M.; Terramani, T.; Lynch, G.; Baudry, M. A Glycine Site Associated with N-Methyl-d-Aspartic Acid Receptors: Characterization and Identification of a New Class of Antagonists. J. Neurochem. 1989, 52, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- DiNatale, B.C.; Murray, I.A.; Schroeder, J.C.; Flaveny, C.A.; Lahoti, T.S.; Laurenzana, E.M.; Omiecinski, C.J.; Perdew, G.H. Kynurenic Acid Is a Potent Endogenous Aryl Hydrocarbon Receptor Ligand that Synergistically Induces Interleukin-6 in the Presence of Inflammatory Signaling. Toxicol. Sci. 2010, 115, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plitman, E.; Iwata, Y.; Caravaggio, F.; Nakajima, S.; Chung, J.K.; Gerretsen, P.; Kim, J.; Takeuchi, H.; Chakravarty, M.M.; Remington, G.; et al. Kynurenic Acid in Schizophrenia: A Systematic Review and Meta-analysis. Schizophr. Bull. 2017, 43, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Schwieler, L.; Larsson, M.K.; Skogh, E.; Kegel, M.E.; Orhan, F.; Abdelmoaty, S.; Finn, A.; Bhat, M.; Samuelsson, M.; Lundberg, K.; et al. Increased levels of IL-6 in the cerebrospinal fluid of patients with chronic schizophrenia—Significance for activation of the kynurenine pathway. J. Psychiatry Neurosci. 2015, 40, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Coleman, M.T.; Hart, R.W.; Liss, L.; Yates, A.J. Changes in Growth and Morphology of Human Gliomas and Fibroblasts Cultured in D-Valine Medium. J. Neuropathol. Exp. Neurol. 1979, 38, 606–613. [Google Scholar] [CrossRef]
- Estin, C.; Vernadakis, A. Primary glial cells and brain fibroblasts: Interactions in culture. Brain Res. Bull. 1986, 16, 723–731. [Google Scholar] [CrossRef]
- Shanks, K.G.; Behonick, G.S. Death after use of the synthetic cannabinoid 5F-AMB. Forensic Sci. Int. 2016, 262, e21–e24. [Google Scholar] [CrossRef]
- Kusano, M.; Zaitsu, K.; Taki, K.; Hisatsune, K.; Nakajima, J.; Moriyasu, T.; Asano, T.; Hayashi, Y.; Tsuchihashi, H.; Ishii, A. Fatal intoxication by 5F-ADB and diphenidine: Detection, quantification, and investigation of their main metabolic pathways in humans by LC/MS/MS and LC/Q-TOFMS. Drug Test. Anal. 2017, 10, 284–293. [Google Scholar] [CrossRef]
- Banister, S.D.; Longworth, M.; Kevin, R.; Sachdev, S.; Santiago, M.; Stuart, J.M.; Mack, J.B.C.; Glass, M.; McGregor, I.S.; Connor, M.; et al. Pharmacology of Valinate and tert-Leucinate Synthetic Cannabinoids 5F-AMBICA, 5F-AMB, 5F-ADB, AMB-FUBINACA, MDMB-FUBINACA, MDMB-CHMICA, and Their Analogues. ACS Chem. Neurosci. 2016, 7, 1241–1254. [Google Scholar] [CrossRef]
- Loopuijt, L.D.; Schmidt, W.J. The role of NMDA receptors in the slow neuronal degeneration of Parkinson’s disease. Amino Acids 1998, 14, 17–23. [Google Scholar] [CrossRef] [PubMed]


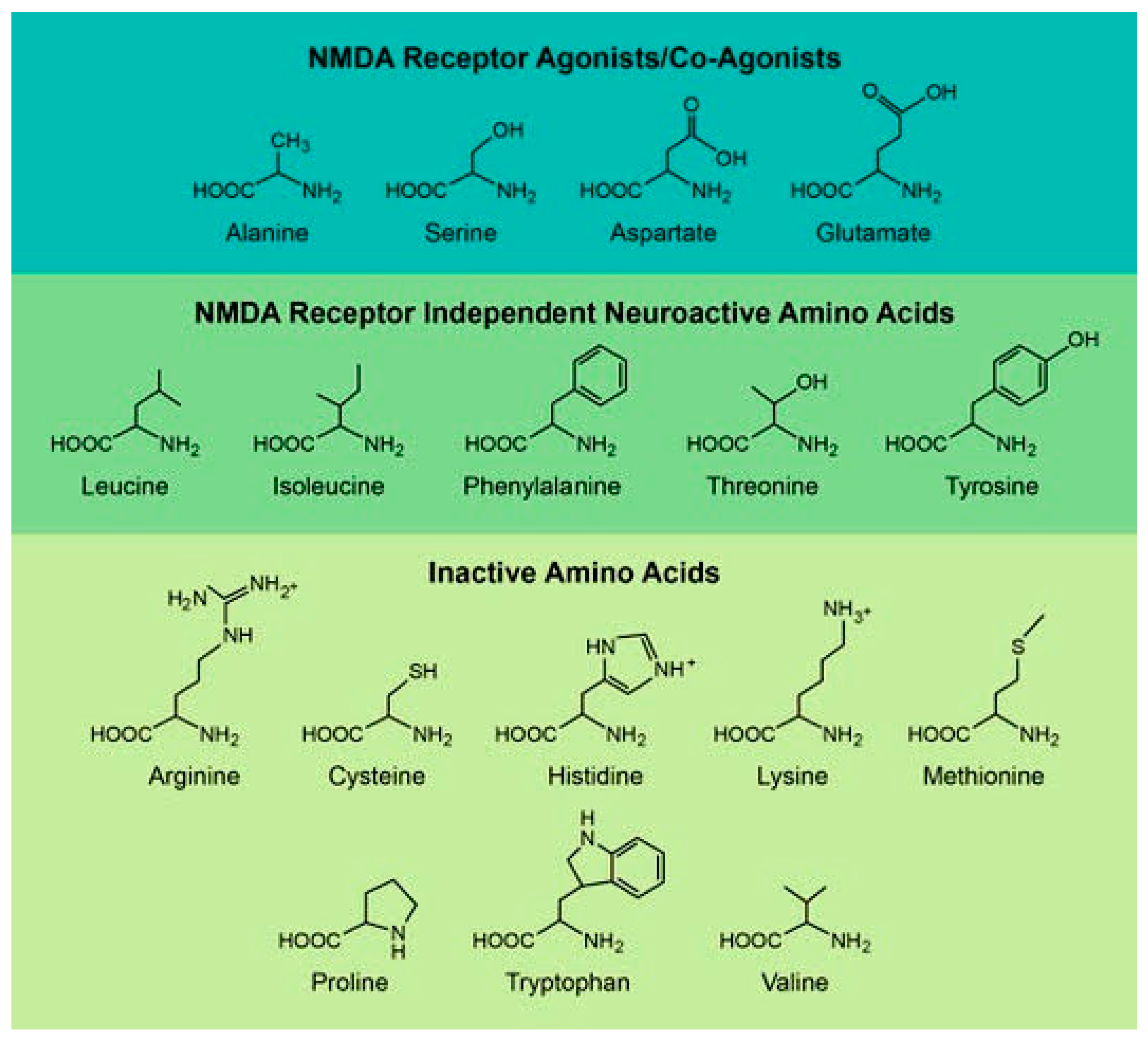{kind=link}
| Amino Acid | Primary Source | Description | Reference |
|---|---|---|---|
| NMDA Receptor Agonists or Co-Agonists | |||
| Alanine | Bacteria/Ingestion | NMDAR 1 co-agonist with links to Alzheimer’s behavioral changes | [13,28,29,30,31,32,33] |
| Aspartate | Bacteria/Racemase | NMDAR 1 agonist with various activities within the brain | [16,34,35,36,37,38,39] |
| Glutamate | Bacteria/Ingestion | NMDAR 1 agonist with links to Alzheimer’s behavioral changes | [14,33,40,41] |
| Serine | Bacteria/Racemase | NMDAR 1 co-agonist with links to Alzheimer’s Disease and Schizophrenia | [42,43,44,45,46,47,48] |
| Active in the Brain through non-NMDA Receptor Pathways | |||
| Isoleucine | Ingestion | Stimulates D-serine release and inhibits glycine release in the brain | [18,49,50,51] |
| Leucine | Ingestion | Anti-seizure agent | [19,52,53,54] |
| Phenylalanine | Ingestion | Carbonic anhydrase activator | [20,55] |
| Threonine | Ingestion | Potentiates currents within the hippocampus | [21] |
| Tyrosine | Ingestion | Tyrosinase inhibitors | [22,56] |
| Amino Acid + B2 | Primary Source | Derivatives | Description | Reference |
|---|---|---|---|---|
| Arginine | Ingestion | poly-18-D-arginine | Derivative is neuro-protective in brain injury models | [57,58,59,60,61,62,63] |
| Cysteine | Ingestion | D-cystine dimethylester | Metabolized to hydrogen sulfide in the brain, and the derivative can rescue opioid induced respiratory depression | [64,65,66,67] |
| Histidine | Ingestion | n.d. 1 | No recently studied activity | None |
| Lysine | Ingestion | Oleoyl-D-lysine | Derivative is a non-opioid analgesic which acts via blocking glycine transport | [25,68,69,70] |
| Methionine | Ingestion | n.d. 1 | No known activity in brain, but protects against ototoxicity in the ear | [71,72,73,74] |
| Proline | Bacteria/Ingestion | D-cis-Fpro | Derivative is a fluorescent probe used to monitor cancer progression | [75,76,77,78,79] |
| Tryptophan | Ingestion | 1-methyl-D-tryptophan | Metabolized to kynurenic acid in the brain, and derivative can be used to inhibit kynurenic acid production | [27,80] |
| Valine | Ingestion | 5F-ADB 2 | Inhibits fibroblast growth, and the derivative is a synthetic cannabinoid with severe and often fatal side effects | [81] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seckler, J.M.; Lewis, S.J. Advances in D-Amino Acids in Neurological Research. Int. J. Mol. Sci. 2020, 21, 7325. https://doi.org/10.3390/ijms21197325
Seckler JM, Lewis SJ. Advances in D-Amino Acids in Neurological Research. International Journal of Molecular Sciences. 2020; 21(19):7325. https://doi.org/10.3390/ijms21197325
Chicago/Turabian StyleSeckler, James M., and Stephen J. Lewis. 2020. "Advances in D-Amino Acids in Neurological Research" International Journal of Molecular Sciences 21, no. 19: 7325. https://doi.org/10.3390/ijms21197325
APA StyleSeckler, J. M., & Lewis, S. J. (2020). Advances in D-Amino Acids in Neurological Research. International Journal of Molecular Sciences, 21(19), 7325. https://doi.org/10.3390/ijms21197325