Full-Length Transcriptome Analysis of the ABCB, PIN/PIN-LIKES, and AUX/LAX Families Involved in Somatic Embryogenesis of Lilium pumilum DC. Fisch.



Abstract
:1. Introduction
2. Results
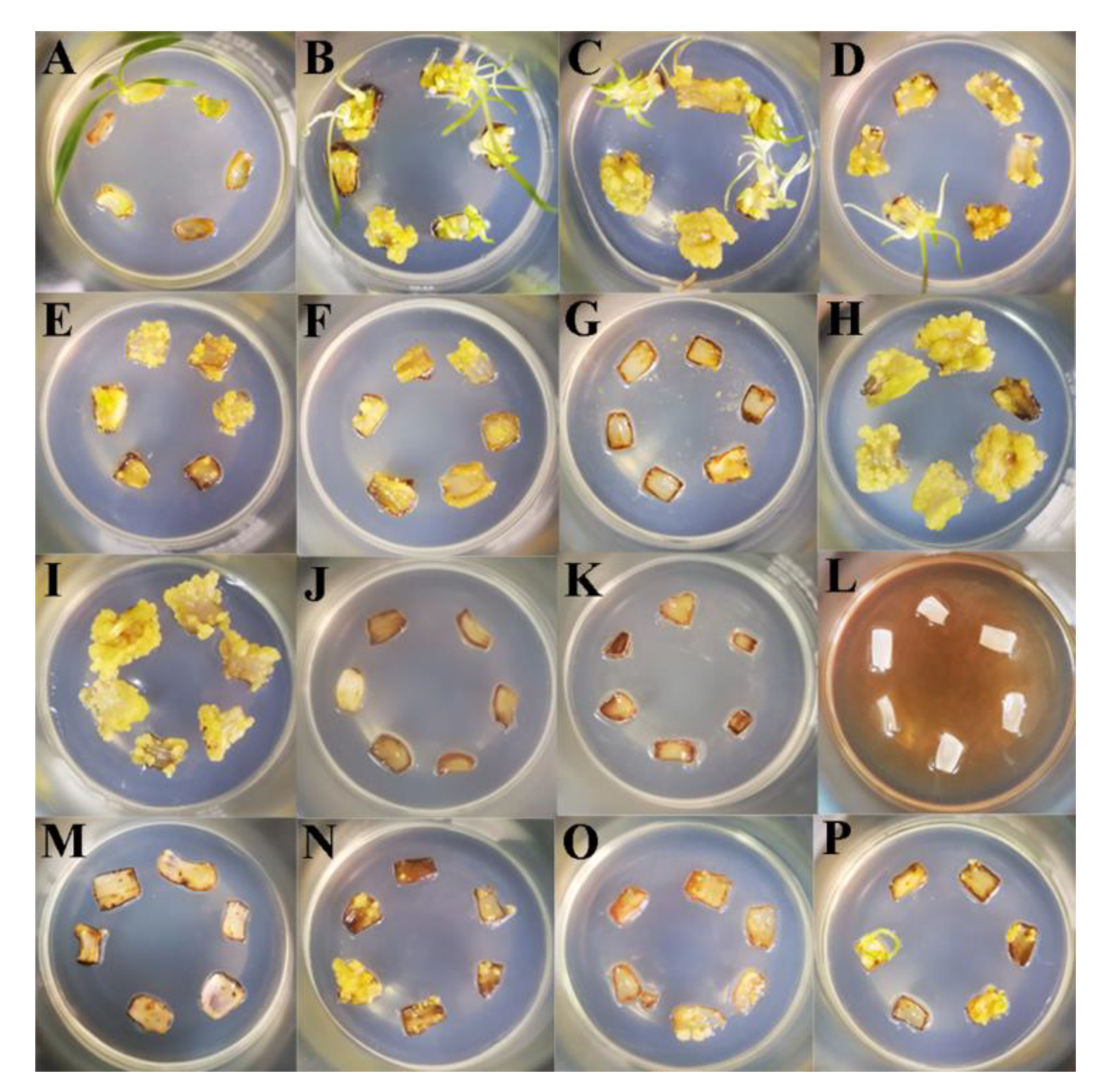
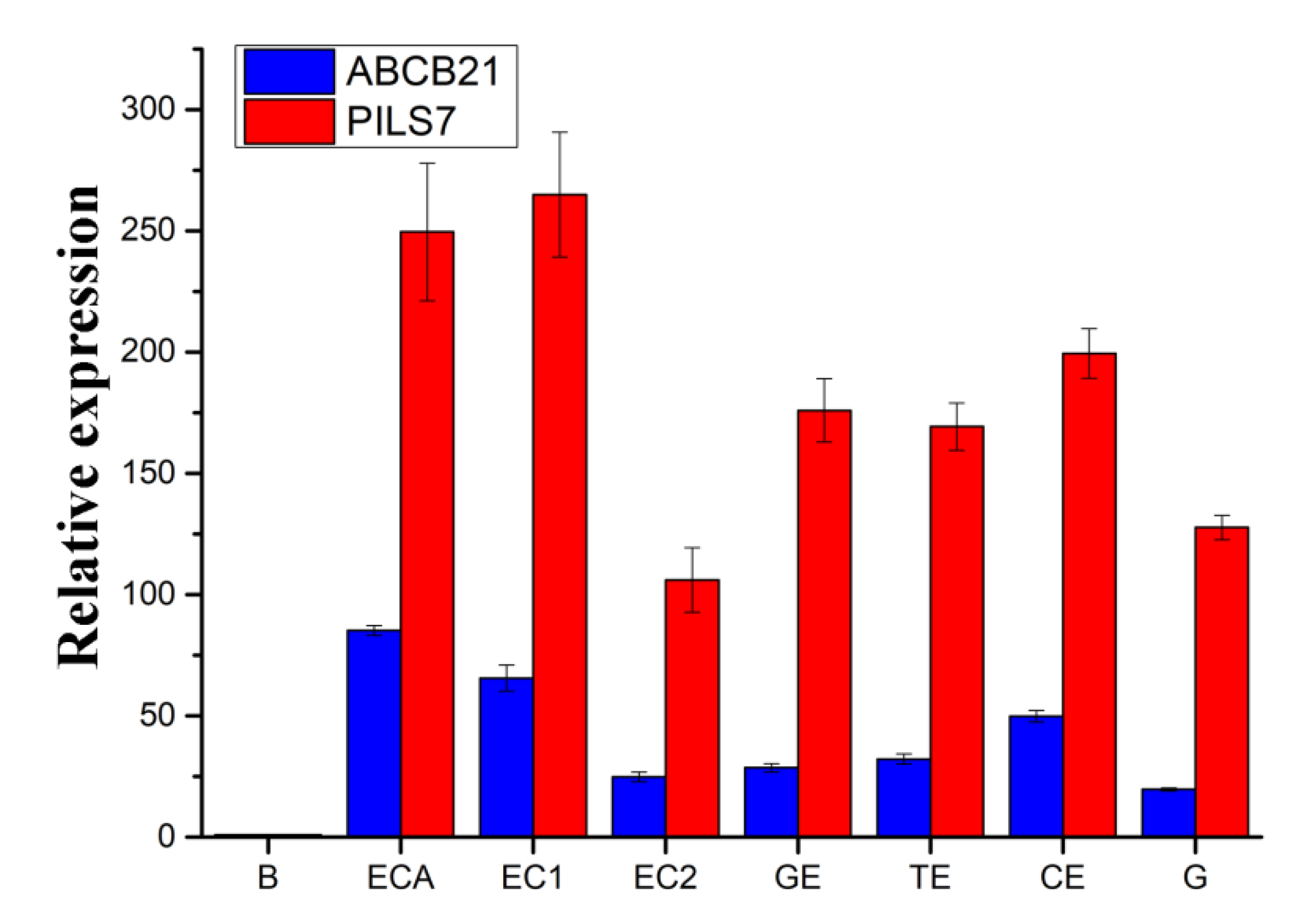
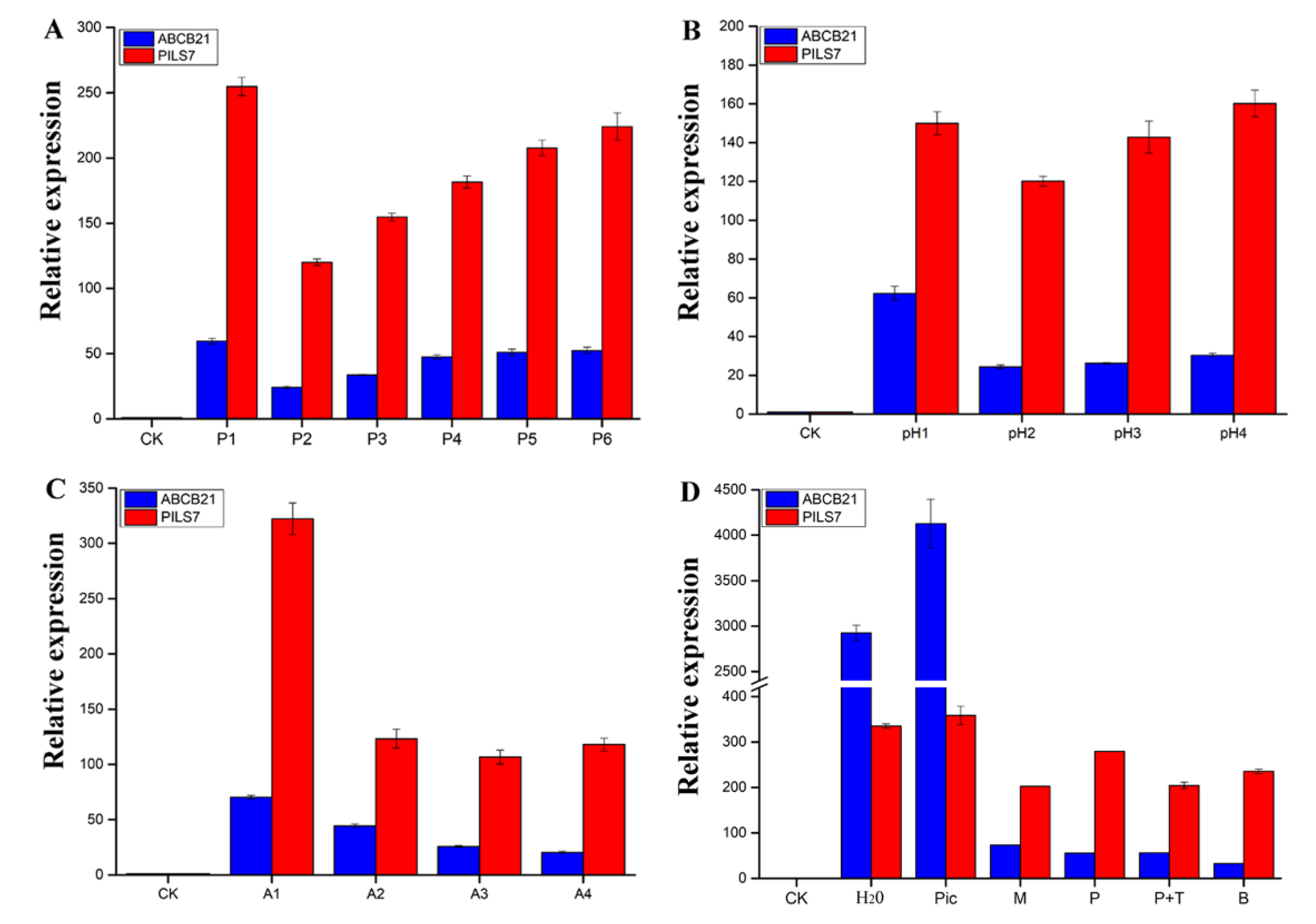
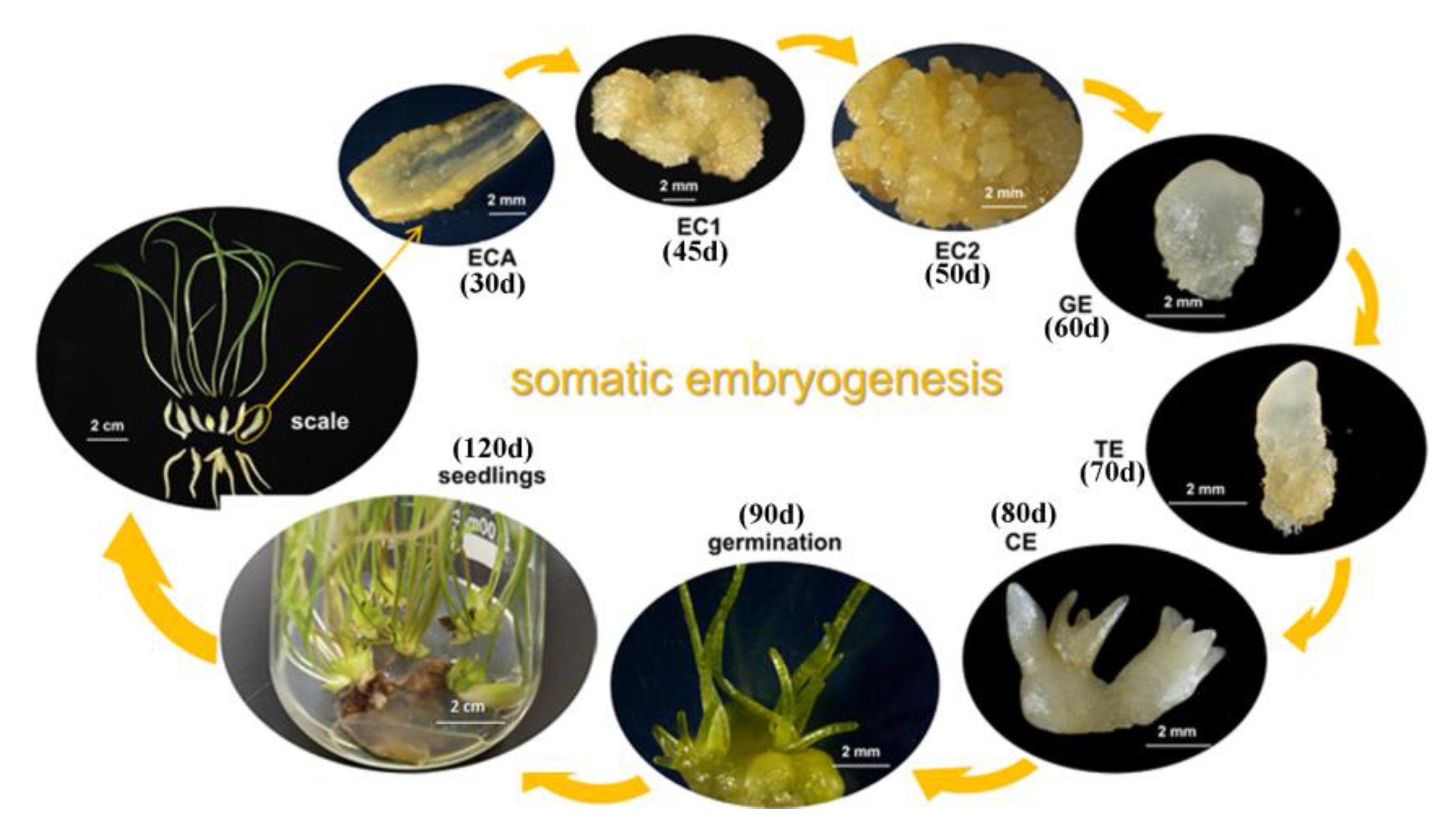
2.1. Effects of External Environmental Factors on SE
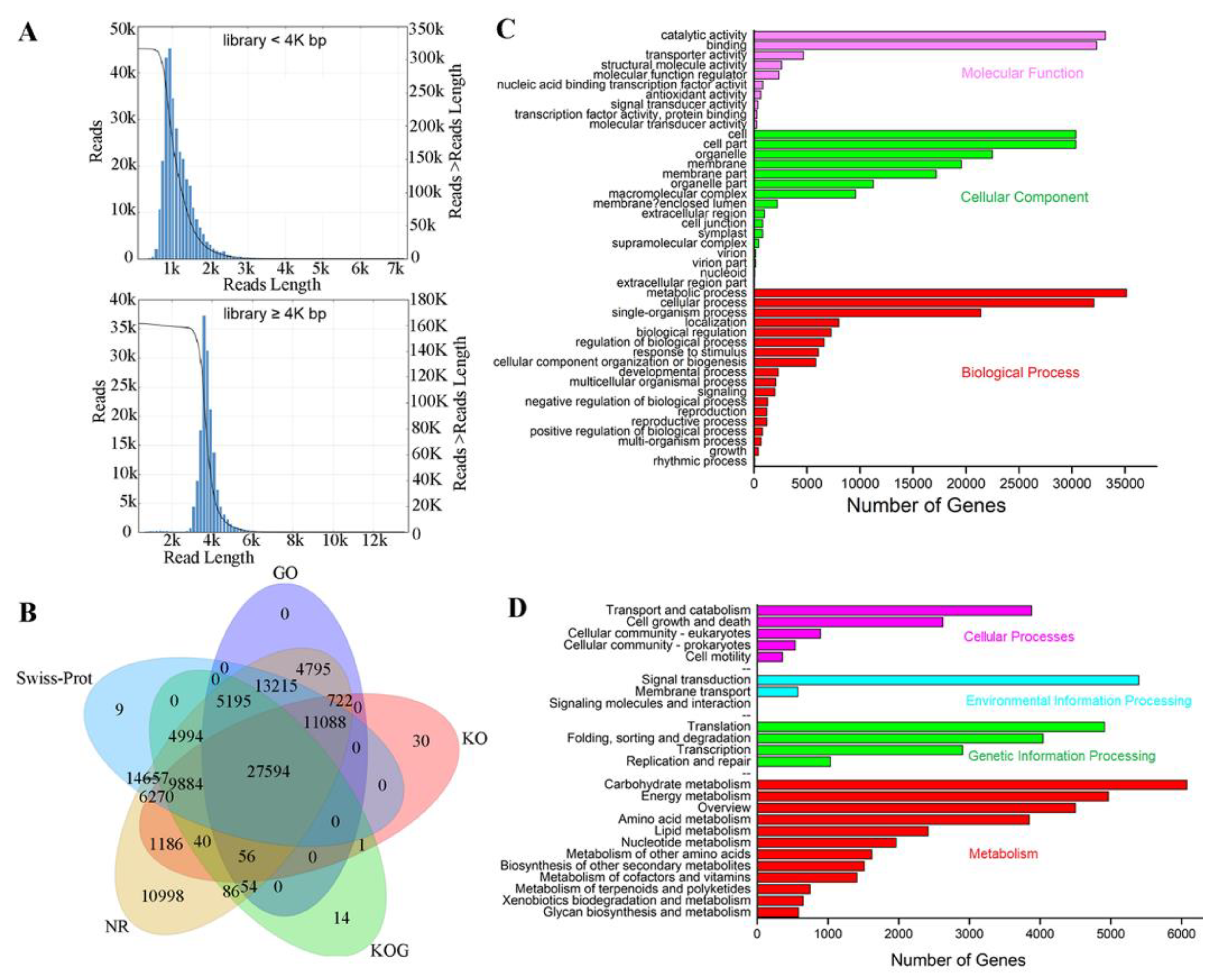
2.2. Comparison of Transcripts between Iso-seq and RNA-seq
2.3. Functional Annotation of Iso-seq Transcripts
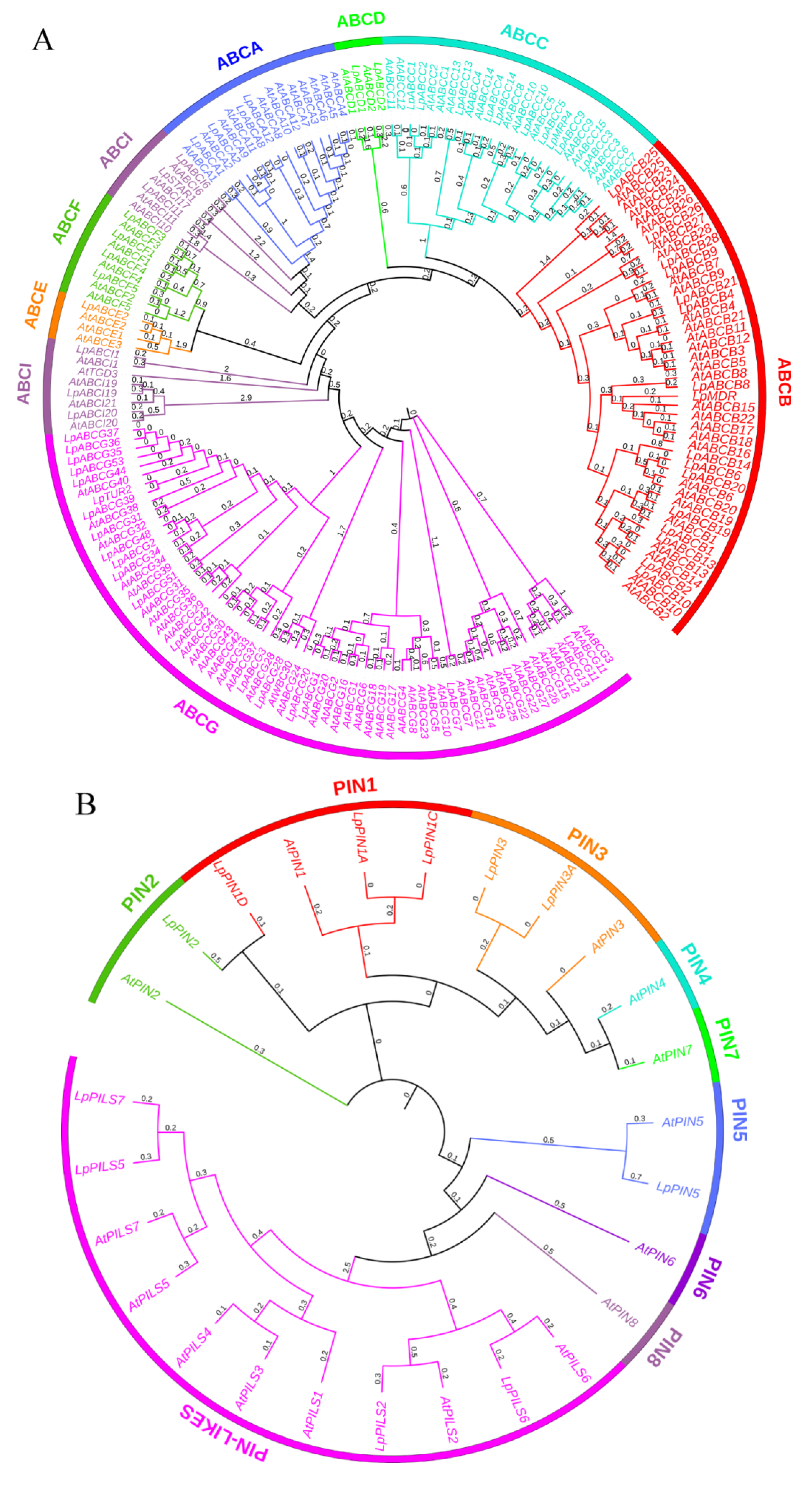
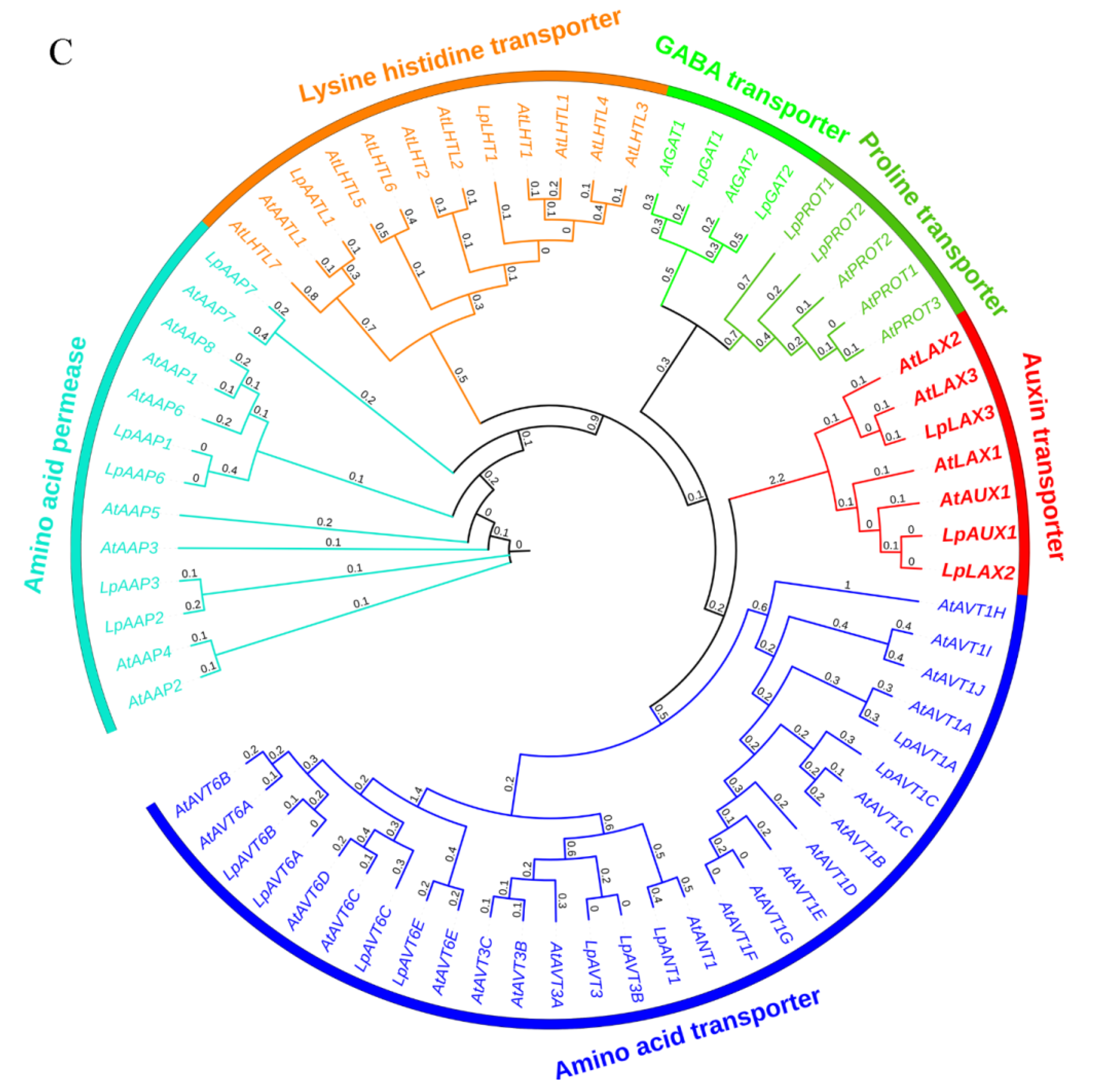
2.4. Identification and Analysis of Family Members Involved in Polar Auxin Transport
2.5. Genetic Analysis of Members of the Polar Auxin Transport Family
2.6. Conserved Domain Analysis of Members of the Polar Auxin Transport Family
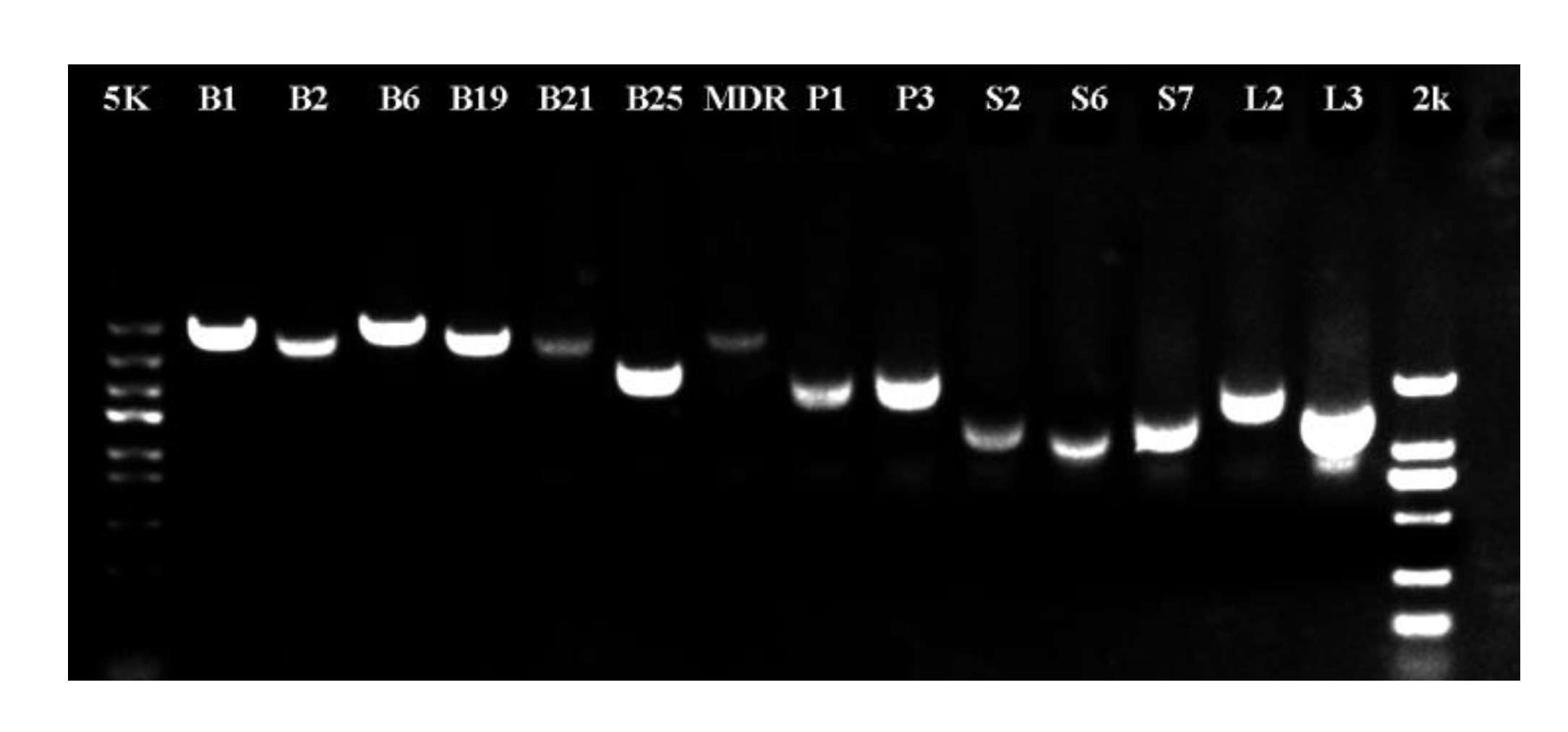
2.7. Cloning and Verification of Genes Involved in Polar Auxin Transport
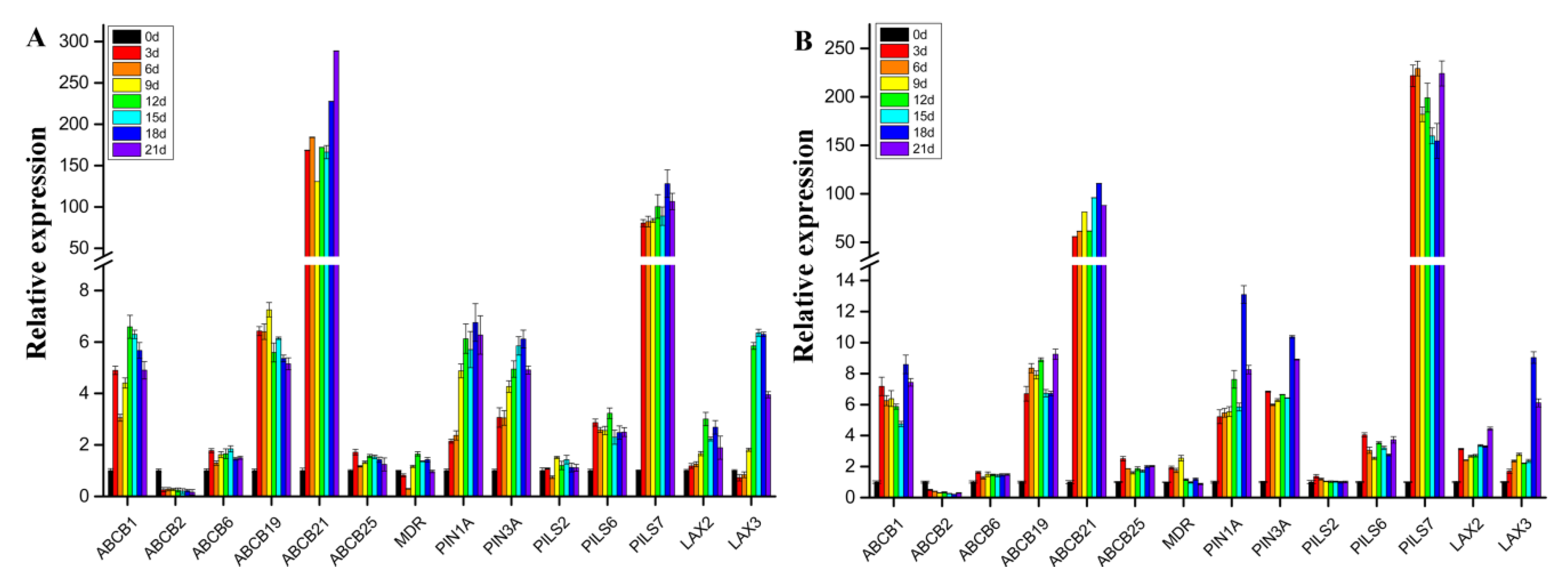
2.8. Expression Pattern Analysis of Genes Involved in Polar Auxin Transport
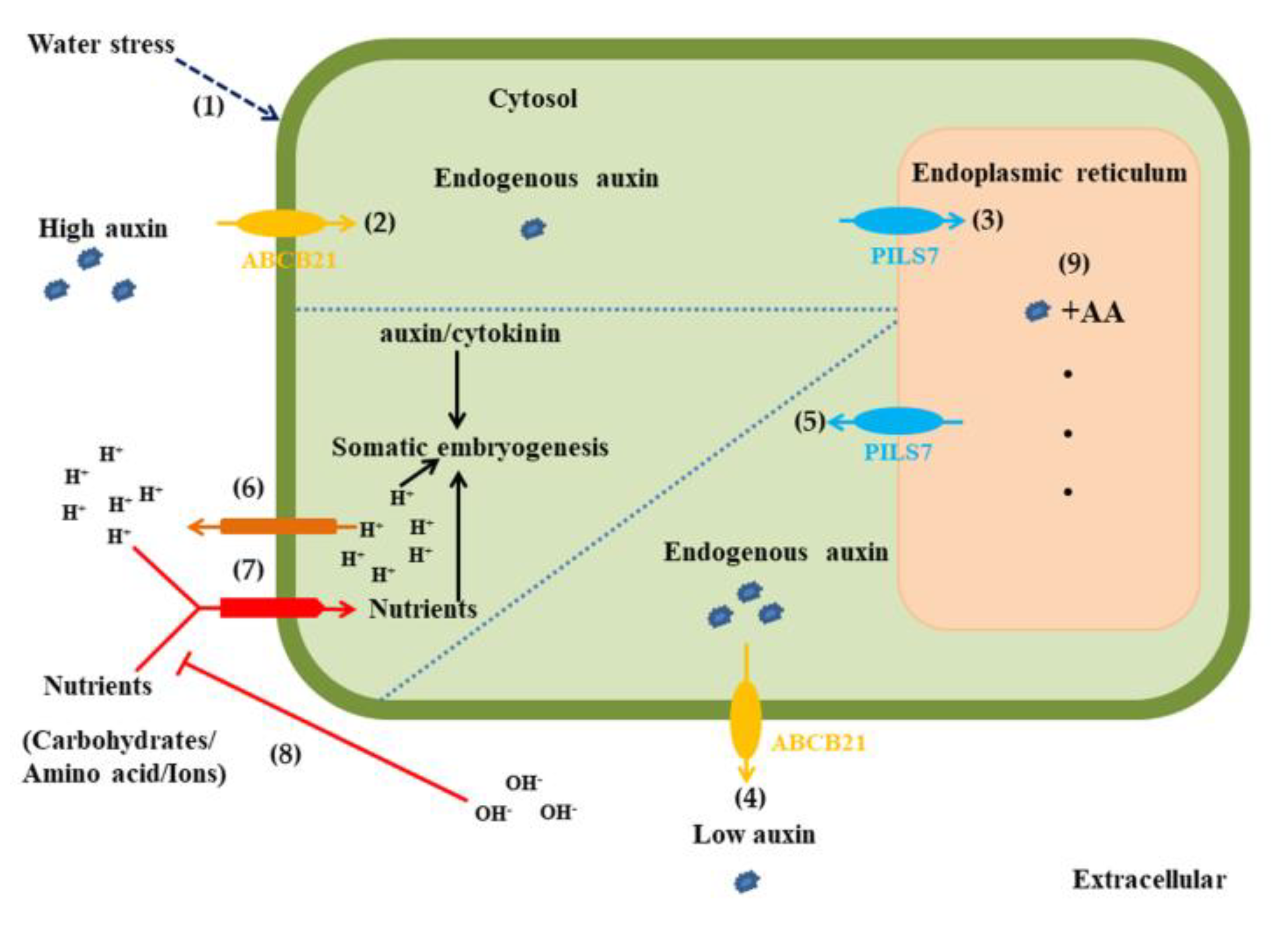
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Total RNA Extraction
4.2. PacBio Library Construction and Sequencing
4.3. Obtaining Full-Length Transcripts and Gene Functional Annotations
4.4. Prediction of the ORFs of Full-Length Transcript Sequences
4.5. Identification and Analysis of Polar Auxin Transport Family Members
4.6. Cloning and Expression Pattern Analysis of Genes Involved in Polar Auxin Transport
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MS (Culture medium) | Murashige-Skoog |
| PIC (IAA analogue) | Picloram (Sigma-Aldrich, USA) |
| 6-BA (Cytokinin) | 6-Benzylaminopurine (Tiangen, Beijing, China) |
| TIBA (Auxin transport inhibitors) | 2,3,5-triiodobenzoic acid (Sigma-Aldrich, USA) |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | PIC (mg/L) | 6-BA (mg/L) | TIBA (mg/L) | MS | pH | Duration (days) | Light/Dark |
|---|---|---|---|---|---|---|---|
| D1-D7 | 1.0 | √ | 5.8 | 3, 6, 9, 12, 15, 18, 21 | Dark | ||
| L1-L7 | 1.0 | √ | 5.8 | 3, 6, 9, 12, 15, 18, 21 | Light | ||
| M | √ | 5.8 | 3, 50 | Light | |||
| P | 1.0 | √ | 5.8 | 3, 50 | Light | ||
| B | 1.5 | √ | 5.8 | 3, 50 | Light | ||
| P+T | 1.0 | 1.0 | √ | 5.8 | 3, 50 | Light | |
| P1 | 0.5 | √ | 5.8 | 3, 50 | Light | ||
| P2 | 1.0 | √ | 5.8 | 3, 50 | Light | ||
| P3 | 2.0 | √ | 5.8 | 3, 50 | Light | ||
| P4 | 3.0 | √ | 5.8 | 3, 50 | Light | ||
| P5 | 4.0 | √ | 5.8 | 3, 50 | Light | ||
| P6 | 5.0 | √ | 5.8 | 3, 50 | Light | ||
| A1 | 1.0 | no macro | 5.8 | 3, 50 | Light | ||
| A2 | 1.0 | no micro | 5.8 | 3, 50 | Light | ||
| A3 | 1.0 | no iron salts | 5.8 | 3, 50 | Light | ||
| A4 | 1.0 | no organic | 5.8 | 3, 50 | Light | ||
| pH1–pH5 | 1.0 | √ | 4.8/5.8/6.8/7.8/9.8 | 3, 50 | Light |
References
- Nic-Can, G.I.; Lopez-Torres, A.; Barredo-Pool, F.; Wrobel, K.; Loyola-Vargas, V.M.; Rojas-Herrera, R.; De-la-Pena, C. New insights into somatic embryogenesis: Leafy cotyledon1, baby boom1 and WUSCHEL-related homeobox4 are epigenetically regulated in Coffea canephora. PLoS ONE 2013, 8, e72160. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, J.L. Somatic embryogenesis: A model for early development in higher plants. Plant Cell 1993, 5, 1411–1423. [Google Scholar] [CrossRef] [Green Version]
- Zeng, F.; Zhang, X.; Zhu, L.; Tu, L.; Guo, X.; Nie, Y. Isolation and characterization of genes associated to cotton somatic embryogenesis by suppression subtractive hybridization and macroarray. Plant Mol. Biol. 2006, 60, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Portillo, L.; Olmedilla, A.; Santacruz-Ruvalcaba, F. Cellular and molecular changes associated with somatic embryogenesis induction in Agave tequilana. Protoplasma 2012, 249, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Guo, H.; Zhang, L.; Fan, Y.; Fan, Y.; Zeng, F. SELTP-assembled battery drives totipotency of somatic plant cell. Plant Biotechnol. J. 2019, 17, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neelakandan, A.K.; Wang, K. Recent progress in the understanding of tissue culture-induced genome level changes in plants and potential applications. Plant Cell Rep. 2012, 31, 597–620. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cai, W.; Liu, Y.; Li, H.; Fu, L.; Liu, Z.; Xu, L.; Liu, H.; Xu, T.; Xiong, Y. Differential TOR activation and cell proliferation in Arabidopsis root and shoot apexes. Proc. Natl. Acad. Sci. USA 2017, 114, 2765–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vondráková, Z.; Eliášová, K.; Fischerová, L.; Vágner, M. The role of auxins in somatic embryogenesis of Abies alba. Cent. Eur. J. Biol. 2011, 6, 587–596. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, C.; Zhang, X.; Liu, C.; Wu, Z.; Yang, Z.; Zhou, K.; Yang, X.; Li, F. Transcriptome profiling reveals auxin and cytokinin regulating somatic embryogenesis in different sister lines of cotton cultivar CCRI24. J. Integr. Plant Biol. 2013, 55, 631–642. [Google Scholar] [CrossRef]
- Fujimura, T. Carrot somatic embryogenesis. A dream come true? Plant Biotechnol. Rep. 2014, 8, 23–28. [Google Scholar] [CrossRef]
- Loyola-Vargas, V.M.; Ochoa-Alejo, N. Somatic embryogenesis. an overview. In Somatic Embryogenesis: Fundamental Aspects and Applications; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–8. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodward, A.W. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, Y.; Men, S.Z.; Fischer, U.; Stepanova, A.N.; Alonso, J.M.; Ljung, K.; Grebe, M. Local auxin biosynthesis modulates gradient-directed planar polarity in Arabidopsis. Nat. Cell Biol. 2009, 11, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, M.J.; Marchant, A.; Green, H.G.; May, S.T.; Ward, S.P.; Millner, P.A.; Walker, A.R.; Schulz, B.; Feldmann, K.A. Arabidopsis AUX1 gene: A permease-like regulator of root gravitropism. Science 1996, 273, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Luschnig, C.; Gaxiola, R.A.; Grisafi, P.; Fink, G.R. EIR1, a root-specific protein involved in auxin transport, is required for gravitropism in Arabidopsis thaliana. Genes Dev. 1998, 12, 2175–2187. [Google Scholar] [CrossRef] [Green Version]
- Geisler, M.; Blakeslee, J.J.; Bouchard, R.; Lee, O.R.; Vincenzetti, V.; Bandyopadhyay, A.; Titapiwatanakun, B.; Peer, W.A.; Bailly, A.; Richards, E.L.; et al. Cellular efflux of auxin catalyzed by the Arabidopsis MDR/PGP transporter AtPGP1. Plant J. 2005, 44, 179–194. [Google Scholar] [CrossRef]
- Petrasek, J.; Mravec, J.; Bouchard, R.; Blakeslee, J.J.; Abas, M.; Seifertova, D.; Wisniewska, J.; Tadele, Z.; Kubes, M.; Covanova, M.; et al. PIN proteins perform a rate-limiting function in cellular auxin efflux. Science 2006, 312, 914–918. [Google Scholar] [CrossRef] [Green Version]
- Zazimalova, E.; Murphy, A.S.; Yang, H.; Hoyerova, K.; Hosek, P. Auxin transporters—Why so many? Cold Spring Harb. Perspect. Biol. 2010, 2, a001552. [Google Scholar] [CrossRef] [Green Version]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef]
- Swarup, R.; Peret, B. AUX/LAX family of auxin influx carriers-an overview. Front. Plant Sci. 2012, 3, 225. [Google Scholar] [CrossRef] [Green Version]
- Barbez, E.; Kubes, M.; Rolcik, J.; Beziat, C.; Pencik, A.; Wang, B.; Rosquete, M.R.; Zhu, J.; Dobrev, P.I.; Lee, Y.; et al. A novel putative auxin carrier family regulates intracellular auxin homeostasis in plants. Nature 2012, 485, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Márquez-López, R.E.; Pérez-Hernández, C.; Ku-González, Á.; Galaz-Ávalos, R.M.; Loyola-Vargas, V.M. Localization and transport of indole-3-acetic acid during somatic embryogenesis in Coffea canephora. Protoplasma 2018, 255, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Klima, P.; Lankova, M.; Zazimalova, E. Inhibitors of plant hormone transport. Protoplasma 2016, 253, 1391–1404. [Google Scholar] [CrossRef]
- Teale, W.; Palme, K. Naphthylphthalamic acid and the mechanism of polar auxin transport. J. Exp. Bot. 2017, 69, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Wang, L.; Chen, J.; Liu, C.; Zeng, H.; Wang, H. Over-expression of KdSOC1 gene affected plantlet morphogenesis in Kalanchoe daigremontiana. Sci. Rep. 2017, 7, 5629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xue, B.; Gai, M.; Song, S.; Jia, N.; Sun, H. Small RNA and transcriptome sequencing reveal a potential miRNA-mediated interaction network that functions during somatic embryogenesis in Lilium pumilum DC. Fisch. Front. Plant Sci. 2017, 8, 566. [Google Scholar] [CrossRef] [PubMed]
- Bakhshaie, M.; Babalar, M.; Mirmasoumi, M.; Khalighi, A. Somatic embryogenesis and plant regeneration of Lilium ledebourii (Baker) Boiss., an endangered species. Plant Cell Tissue Organ Cult. 2010, 102, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.-W.; Jian, W.-T.; Lai, H.-C. Plant regeneration via somatic embryogenesis from suspension cell cultures of Lilium × formolongi Hort. using a bioreactor system. Vitr. Cell. Dev. Biol. Plant 2006, 42, 240–246. [Google Scholar] [CrossRef]
- Nhut, D.T.; Hanh, N.T.M.; Tuan, P.Q.; Tram, N.T.H.; Chinh, N.C.; Nguyen, N.H. Liquid culture as a positive condition to induce and enhance quality and quantity of somatic embryogenesis of Lilium longiflorum. Sci. Hortic. 2006, 110, 93–97. [Google Scholar] [CrossRef]
- KEdra, M.; Bach, A. Morphogenesis of Lilium martagon L. explants in callus culture. Acta Biol. Crac. Ser. Bot. 2005, 47, 65–73. [Google Scholar]
- Li, H.; Zhang, J.; Yang, Y.; Jia, N.; Wang, C.; Sun, H. miR171 and its target gene SCL6 contribute to embryogenic callus induction and torpedo-shaped embryo formation during somatic embryogenesis in two lily species. Plant Cell Tissue Organ Cult. 2017, 130, 591–600. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, J.; Wang, J.; Ren, Y.; Zhu, Y.; Sun, H. Dynamic changes of miR166s at both the transcriptional and post-transcriptional levels during somatic embryogenesis in Lilium. Sci. Hortic. 2020, 261, 108928. [Google Scholar] [CrossRef]
- Jenness, M.K.; Carraro, N.; Pritchard, C.A.; Murphy, A.S. The Arabidopsis ATP-BINDING CASSETTE transporter ABCB21 regulates auxin levels in cotyledons, the root pericycle, and leaves. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, C.; Wang, Y.; Valliyodan, B.; Nguyen, H.T. Comprehensive analysis of the soybean (Glycine max) GmLAX auxin transporter gene family. Front. Plant Sci. 2016, 7, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carraro, N.; Tisdale-Orr, T.E.; Clouse, R.M.; Knoller, A.S.; Spicer, R. Diversification and expression of the PIN, AUX/LAX, and ABCB families of putative auxin transporters in Populus. Front. Plant Sci. 2012, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Ma, T.; Wang, X.; Deng, Y.; Ma, H.; Zhang, R.; Zhao, J. OsAUX1 controls lateral root initiation in rice (Oryza sativa L.). Plant Cell Environ. 2015, 38, 2208–2222. [Google Scholar] [CrossRef]
- Olawuyi, O.; Ologidi, C. Genomic survey of ATP-binding cassette (ABC) transporters in Sorghum bicolor (L.) Moench. J. Plant Sci. Crop Prot. 2018, 1, 102. [Google Scholar] [CrossRef]
- Yu, C.; Dong, W.; Zhan, Y.; Huang, Z.A.; Li, Z.; Kim, I.S.; Zhang, C. Genome-wide identification and expression analysis of ClLAX, ClPIN and ClABCB genes families in Citrullus lanatus under various abiotic stresses and grafting. BMC Genet 2017, 18, 33. [Google Scholar] [CrossRef] [Green Version]
- Yue, R.; Tie, S.; Sun, T.; Zhang, L.; Yang, Y.; Qi, J.; Yan, S.; Han, X.; Wang, H.; Shen, C. Genome-wide identification and expression profiling analysis of ZmPIN, ZmPILS, ZmLAX and ZmABCB auxin transporter gene families in maize (Zea mays L.) under various abiotic stresses. PLoS ONE 2015, 10, e0118751. [Google Scholar] [CrossRef]
- Lan, P.; Li, W.; Schmidt, W. Complementary proteome and transcriptome profiling in phosphate-deficient Arabidopsis roots reveals multiple levels of gene regulation. Mol. Cell. Proteom. 2012, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oono, Y.; Kawahara, Y.; Yazawa, T.; Kanamori, H.; Kuramata, M.; Yamagata, H.; Hosokawa, S.; Minami, H.; Ishikawa, S.; Wu, J.Z.; et al. Diversity in the complexity of phosphate starvation transcriptomes among rice cultivars based on RNA-Seq profiles. Plant Mol. Biol. 2013, 83, 523–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.X.; Xu, C.Z.; Li, K.P.; Yan, S.; Qu, X.; Zhang, J.R. Phosphate starvation of maize inhibits lateral root formation and alters gene expression in the lateral root primordium zone. BMC Plant Biol. 2012, 12, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilgner, H.; Jahanbani, F.; Blauwkamp, T.; Moshrefi, A.; Jaeger, E.; Chen, F.; Harel, I.; Bustamante, C.D.; Rasmussen, M.; Snyder, M.P. Comprehensive transcriptome analysis using synthetic long-read sequencing reveals molecular co-association of distant splicing events. Nat. Biotechnol. 2015, 33, 736–742. [Google Scholar] [CrossRef]
- VanBuren, R.; Bryant, D.; Edger, P.P.; Tang, H.B.; Burgess, D.; Challabathula, D.; Spittle, K.; Hall, R.; Gu, J.; Lyons, E.; et al. Single-molecule sequencing of the desiccation-tolerant grass Oropetium thomaeum. Nature 2015, 527, 508–511. [Google Scholar] [CrossRef] [Green Version]
- Lan, T.Y.; Renner, T.; Ibarra-Laclette, E.; Farr, K.M.; Chang, T.H.; Cervantes-Perez, S.A.; Zheng, C.F.; Sankoff, D.; Tang, H.B.; Purbojati, R.W.; et al. Long-read sequencing uncovers the adaptive topography of a carnivorous plant genome. Proc. Natl. Acad. Sci. USA 2017, 114, E4435–E4441. [Google Scholar] [CrossRef] [Green Version]
- Ren, P.; Meng, Y.; Li, B.; Ma, X.; Si, E.; Lai, Y.; Wang, J.; Yao, L.; Yang, K.; Shang, X. Molecular mechanisms of acclimatization to phosphorus starvation and recovery underlying full-length transcriptome profiling in Barley (Hordeum vulgare L.). Front. Plant Sci. 2018, 9, 500. [Google Scholar] [CrossRef]
- Xu, Z.C.; Peters, R.J.; Weirather, J.; Luo, H.M.; Liao, B.S.; Zhang, X.; Zhu, Y.J.; Ji, A.J.; Zhang, B.; Hu, S.N.; et al. Full-length transcriptome sequences and splice variants obtained by a combination of sequencing platforms applied to different root tissues of Salvia miltiorrhiza and tanshinone biosynthesis. Plant J. 2015, 82, 951–961. [Google Scholar] [CrossRef]
- Smeekens, S.; Ma, J.K.; Hanson, J.; Rolland, F. Sugar signals and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 274–279. [Google Scholar] [CrossRef]
- Businge, E.; Brackmann, K.; Moritz, T.; Egertsdotter, U. Metabolite profiling reveals clear metabolic changes during somatic embryo development of Norway spruce (Picea abies). Tree Physiol. 2012, 32, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Kube, M.; Drazna, N.; Konradova, H.; Lipavska, H. Robust carbohydrate dynamics based on sucrose resynthesis in developing Norway spruce somatic embryos at variable sugar supply. Vitr. Cell. Dev. Biol. Plant 2014, 50, 45–57. [Google Scholar] [CrossRef]
- Skylar, A.; Sung, F.; Hong, F.; Chory, J.; Wu, X. Metabolic sugar signal promotes Arabidopsis meristematic proliferation via G2. Dev. Biol. 2011, 351, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.S.; De Vega, J.J.; Miguel, C.M. Comprehensive assembly and analysis of the transcriptome of maritime pine developing embryos. BMC Plant Biol. 2018, 18, 379. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hammes, U.Z.; Taylor, C.G.; Schachtman, D.P.; Nielsen, E. High-affinity auxin transport by the AUX1 influx carrier protein. Curr. Biol. 2006, 16, 1123–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarup, K.; Benkova, E.; Swarup, R.; Casimiro, I.; Peret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S.; et al. The auxin influx carrier LAX3 promotes lateral root emergence. Nat. Cell Biol. 2008, 10, 946–954. [Google Scholar] [CrossRef]
- Peret, B.; Swarup, K.; Ferguson, A.; Seth, M.; Yang, Y.D.; Dhondt, S.; James, N.; Casimiro, I.; Perry, P.; Syed, A.; et al. AUX/LAX genes encode a family of auxin influx transporters that perform distinct functions during Arabidopsis development. Plant Cell 2012, 24, 2874–2885. [Google Scholar] [CrossRef] [Green Version]
- Pang, K.; Li, Y.; Liu, M.; Meng, Z.; Yu, Y. Inventory and general analysis of the ATP-binding cassette (ABC) gene superfamily in maize (Zea mays L.). Gene 2013, 526, 411–428. [Google Scholar] [CrossRef]
- Ma, J.E.; Jiang, H.Y.; Li, L.M.; Zhang, X.J.; Li, H.M.; Li, G.Y.; Mo, D.Y.; Chen, J.P. SMRT sequencing of the full-length transcriptome of the sunda pangolin (Manis javanica). Gene 2019, 692, 208–216. [Google Scholar] [CrossRef]
- Mikula, A.; Pozoga, M.; Tomiczak, K.; Rybczynski, J.J. Somatic embryogenesis in ferns: A new experimental system. Plant Cell Rep. 2015, 34, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, V.M.; Guevara, E.; Herrera, J.; Bangerth, F. Evolution of endogenous hormone concentration in embryogenic cultures of carrot during early expression of somatic embryogenesis. Plant Cell Rep. 2005, 23, 567–572. [Google Scholar] [CrossRef]
- Cleland, R.E.; Buckley, G.; Nowbar, S.; Lew, N.M.; Stinemetz, C.; Evans, M.L.; Rayle, D.L. The pH profile for acid-induced elongation of coleoptile and epicotyl sections is consistent with the acid-growth theory. Planta 1991, 186, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Rayle, D.L.; Cleland, R.E. The Acid Growth Theory of auxin-induced cell elongation is alive and well. Plant Physiol. 1992, 99, 1271–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heringer, A.S.; Barroso, T.; Macedo, A.F.; Santa-Catarina, C.; Souza, G.H.; Floh, E.I.; de Souza-Filho, G.A.; Silveira, V. Label-free quantitative proteomics of embryogenic and non-embryogenic callus during sugarcane somatic embryogenesis. PLoS ONE 2015, 10, e0127803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, R.S.; Vale Ede, M.; Heringer, A.S.; Santa-Catarina, C.; Silveira, V. Putrescine induces somatic embryo development and proteomic changes in embryogenic callus of sugarcane. J. Proteom. 2016, 130, 170–179. [Google Scholar] [CrossRef]
- Passamani, L.Z.; Bertolaz, A.A.; Ramos, A.C.; Santa-Catarina, C.; Thelen, J.J.; Silveira, V. Embryogenic competence acquisition in sugar cane callus is associated with differential H+-pump abundance and activity. J. Proteome Res. 2018, 17, 2767–2779. [Google Scholar] [CrossRef]
- Gorst, J.; Overall, R.L.; Wernicke, W. Ionic currents traversing cell clusters from carrot suspension cultures reveal perpetuation of morphogenetic potential as distinct from induction of embryogenesis. Cell Differ. 1987, 21, 101–109. [Google Scholar] [CrossRef]
- Feher, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tissue Organ Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Yang, H.; Murphy, A.S. Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J. 2009, 59, 179–191. [Google Scholar] [CrossRef]
- Kamimoto, Y.; Terasaka, K.; Hamamoto, M.; Takanashi, K.; Fukuda, S.; Shitan, N.; Sugiyama, A.; Suzuki, H.; Shibata, D.; Wang, B.; et al. Arabidopsis ABCB21 is a facultative auxin importer/exporter regulated by cytoplasmic auxin concentration. Plant Cell Physiol. 2012, 53, 2090–2100. [Google Scholar] [CrossRef] [Green Version]
- Pedroso, M.C.; Pais, M.S. A scanning electron microscopy and X-ray microanalysis study during induction of morphogenesis in Camellia japonica L. Plant Sci. 1992, 87, 99–108. [Google Scholar] [CrossRef]
- Pedroso, M.C.; Pais, M.S. Direct embryo formation in leaves of Camillia japonica L. Plant Cell Rep. 1993, 12, 639–643. [Google Scholar] [CrossRef]
- Pedroso, M.C.; Pais, M.S. Factors controlling somatic embryogenesis. Plant Cell Tissue Organ Cult. 1995, 43, 147–154. [Google Scholar] [CrossRef]
- Marimuthu, K.; Subbaraya, U.; Suthanthiram, B.; Marimuthu, S.S. Molecular analysis of somatic embryogenesis through proteomic approach and optimization of protocol in recalcitrant Musa spp. Physiol. Plant 2019, 167, 282–301. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Antoniadi, I.; Plackova, L.; Simonovik, B.; Dolezal, K.; Turnbull, C.; Ljung, K.; Novak, O. Cell-type-specific cytokinin distribution within the Arabidopsis primary root apex. Plant Cell 2015, 27, 1955–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, M.; Li, Y.; Zhang, Q.; Lindsey, K.; Daniell, H.; Jin, S.; Zhang, X. Multi-omics analyses reveal epigenomics basis for cotton somatic embryogenesis through successive regeneration acclimation process. Plant Biotechnol. J. 2019, 17, 435–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.J. Somatic embryogenesis in the Medicago truncatula model: Cellular and molecular mechanisms. Front. Plant Sci. 2019, 10, 267. [Google Scholar] [CrossRef]
- Feher, A. Somatic embryogenesis—Stress-induced remodeling of plant cell fate. Bba-Gene Regul. Mech. 2015, 1849, 385–402. [Google Scholar] [CrossRef]
- Yang, X.Y.; Zhang, X.L. Regulation of somatic embryogenesis in higher plants. Crit. Rev. Plant Sci. 2010, 29, 36–57. [Google Scholar] [CrossRef]
- Leljak-Levanic, D.; Mihaljevic, S.; Bauer, N. Somatic and zygotic embryos share common developmental features at the onset of plant embryogenesis. Acta Physiol. Plant 2015, 37. [Google Scholar] [CrossRef]
- Feraru, E.; Feraru, M.I.; Barbez, E.; Waidmann, S.; Sun, L.; Gaidora, A.; Kleine-Vehn, J. PILS6 is a temperature-sensitive regulator of nuclear auxin input and organ growth in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2019, 116, 3893–3898. [Google Scholar] [CrossRef] [Green Version]
- Moncalean, P.; Garcia-Mendiguren, O.; Novak, O.; Strnad, M.; Goicoa, T.; Ugarte, M.D.; Montalban, I.A. Temperature and water availability during maturation affect the cytokinins and auxins profile of radiata pine somatic embryos. Front. Plant Sci. 2018, 9, 1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonga, J.M.; Klimaszewska, K.K.; von Aderkas, P. Recalcitrance in clonal propagation, in particular of conifers. Plant Cell Tissue Organ Cult. 2010, 100, 241–254. [Google Scholar] [CrossRef]
- Belin, C.; Megies, C.; Hauserova, E.; Lopez-Molina, L. Abscisic acid represses growth of the Arabidopsis embryonic axis after germination by enhancing auxin signaling. Plant Cell 2009, 21, 2253–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Yang, Y.N.; Xue, L.J.; Zou, M.J.; Liu, J.Y.; Chen, F.; Xue, H.W. Rice ABI5-Like1 regulates abscisic acid and auxin responses by affecting the expression of ABRE-containing genes. Plant Physiol. 2011, 156, 1397–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, F.Y.; Hu, L.S.; Yuan, D.J.; Xu, J.; Gao, W.H.; He, L.R.; Yang, X.Y.; Zhang, X.L. Comparative transcriptome analysis between somatic embryos (SEs) and zygotic embryos in cotton: Evidence for stress response functions in SE development. Plant Biotechnol. J. 2014, 12, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Sharp, R.E. Complexity and coordination of root growth at low water potentials: Recent advances from transcriptomic and proteomic analyses. Plant Cell Environ. 2010, 33, 590–603. [Google Scholar] [CrossRef]
- Xu, W.; Jia, L.; Shi, W.; Liang, J.; Zhou, F.; Li, Q.; Zhang, J. Abscisic acid accumulation modulates auxin transport in the root tip to enhance proton secretion for maintaining root growth under moderate water stress. New Phytol. 2013, 197, 139–150. [Google Scholar] [CrossRef]
- Pitts, R.J.; Cernac, A.; Estelle, M. Auxin and ethylene promote root hair elongation in Arabidopsis. Plant J. 1998, 16, 553–560. [Google Scholar] [CrossRef]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Jones, A.R.; Kramer, E.M.; Knox, K.; Swarup, R.; Bennett, M.J.; Lazarus, C.M.; Leyser, H.M.O.; Grierson, C.S. Auxin transport through non-hair cells sustains root-hair development. Nat. Cell Biol. 2009, 11, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staal, M.; De Cnodder, T.; Simon, D.; Vandenbussche, F.; Van Der Straeten, D.; Verbelen, J.P.; Elzenga, T.; Vissenberg, K. Apoplastic alkalinization is instrumental for the inhibition of cell elongation in the Arabidopsis root by the ethylene precursor 1-aminocyclopropane-1-carboxylic acid. Plant Physiol. 2011, 155, 2049–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.; Neumann, P.M. The spatially variable inhibition by water deficit of maize root growth correlates with altered profiles of proton flux and cell wall pH. Plant Physiol. 2004, 135, 2291–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feher, A. Callus, dedifferentiation, totipotency, somatic embryogenesis: What these terms mean in the era of molecular plant biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Li, X.; Cheng, J.; Zhang, J.; Teixeira da Silva, J.A.; Wang, C.; Sun, H. Validation of reference genes for accurate normalization of gene expression in Lilium davidii var. unicolor for real time quantitative PCR. PLoS ONE 2015, 10, e0141323. [Google Scholar] [CrossRef]












| Treatment | PIC Treatment Duration(days) | Culture Duration in MS(days) | Inductivity of Callus (No.) | Inductivity of Buds (No.) | No. of Roots |
|---|---|---|---|---|---|
| Light culture | 3 | 47 | 80.0% (24) | 50.0% (47) | 18 |
| 6 | 44 | 90.0% (27) | 50.0% (41) | 25 | |
| 9 | 41 | 96.7% (29) | 36.7% (39) | 18 | |
| 12 | 38 | 100% (30) | 30.0% (29) | 20 | |
| 15 | 35 | 100% (30) | 26.7% (20) | 5 | |
| 18 | 32 | 100% (30) | 26.7% (20) | 19 | |
| 21 | 29 | 100% (30) | 20.0% (15) | 15 | |
| 50 | 0 | 100% (30) | 13.3% (4) | 0 | |
| Darkness culture | 3 | 47 | 33.3% (10) | 10.0% (8) | 12 |
| 6 | 44 | 43.3% (13) | 10.0% (4) | 2 | |
| 9 | 41 | 66.7% (20) | 10.0% (10) | 5 | |
| 12 | 38 | 80.0% (24) | 16.7% (11) | 15 | |
| 15 | 35 | 96.7% (29) | 13.3% (6) | 18 | |
| 18 | 32 | 100% (30) | 10.0% (9) | 23 | |
| 21 | 29 | 100% (30) | 3.3% (1) | 26 | |
| 50 | 0 | 100% (30) | 0 | 0 |
| Treatment | M | P1 | P2 | P3 | P4 | P5 | P6 | pH1 | pH2 | pH3 | pH4 | pH5 | A1 | A2 | A3 | A4 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of callus | 2 | 15 | 18 | 15 | 9 | 7 | 3 | 17 | 18 | 2 | 0 | 0 | 2 | 15 | 12 | 12 |
| Inductivity/% | 11.1 | 83.3 | 100 | 83.3 | 50.0 | 38.9 | 16.7 | 94.4 | 100 | 11.1 | 0 | 0 | 11.1 | 83.3 | 66.7 | 66.7 |
| No. of buds | 3 | 28 | 16 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.; Wang, Z.; Ren, Y.; Sun, H. Full-Length Transcriptome Analysis of the ABCB, PIN/PIN-LIKES, and AUX/LAX Families Involved in Somatic Embryogenesis of Lilium pumilum DC. Fisch. Int. J. Mol. Sci. 2020, 21, 453. https://doi.org/10.3390/ijms21020453
Song S, Wang Z, Ren Y, Sun H. Full-Length Transcriptome Analysis of the ABCB, PIN/PIN-LIKES, and AUX/LAX Families Involved in Somatic Embryogenesis of Lilium pumilum DC. Fisch. International Journal of Molecular Sciences. 2020; 21(2):453. https://doi.org/10.3390/ijms21020453
Chicago/Turabian StyleSong, Shengli, Zhiping Wang, Yamin Ren, and Hongmei Sun. 2020. "Full-Length Transcriptome Analysis of the ABCB, PIN/PIN-LIKES, and AUX/LAX Families Involved in Somatic Embryogenesis of Lilium pumilum DC. Fisch." International Journal of Molecular Sciences 21, no. 2: 453. https://doi.org/10.3390/ijms21020453
APA StyleSong, S., Wang, Z., Ren, Y., & Sun, H. (2020). Full-Length Transcriptome Analysis of the ABCB, PIN/PIN-LIKES, and AUX/LAX Families Involved in Somatic Embryogenesis of Lilium pumilum DC. Fisch. International Journal of Molecular Sciences, 21(2), 453. https://doi.org/10.3390/ijms21020453