A Sub-Clone of RAW264.7-Cells Form Osteoclast-Like Cells Capable of Bone Resorption Faster than Parental RAW264.7 through Increased De Novo Expression and Nuclear Translocation of NFATc1

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
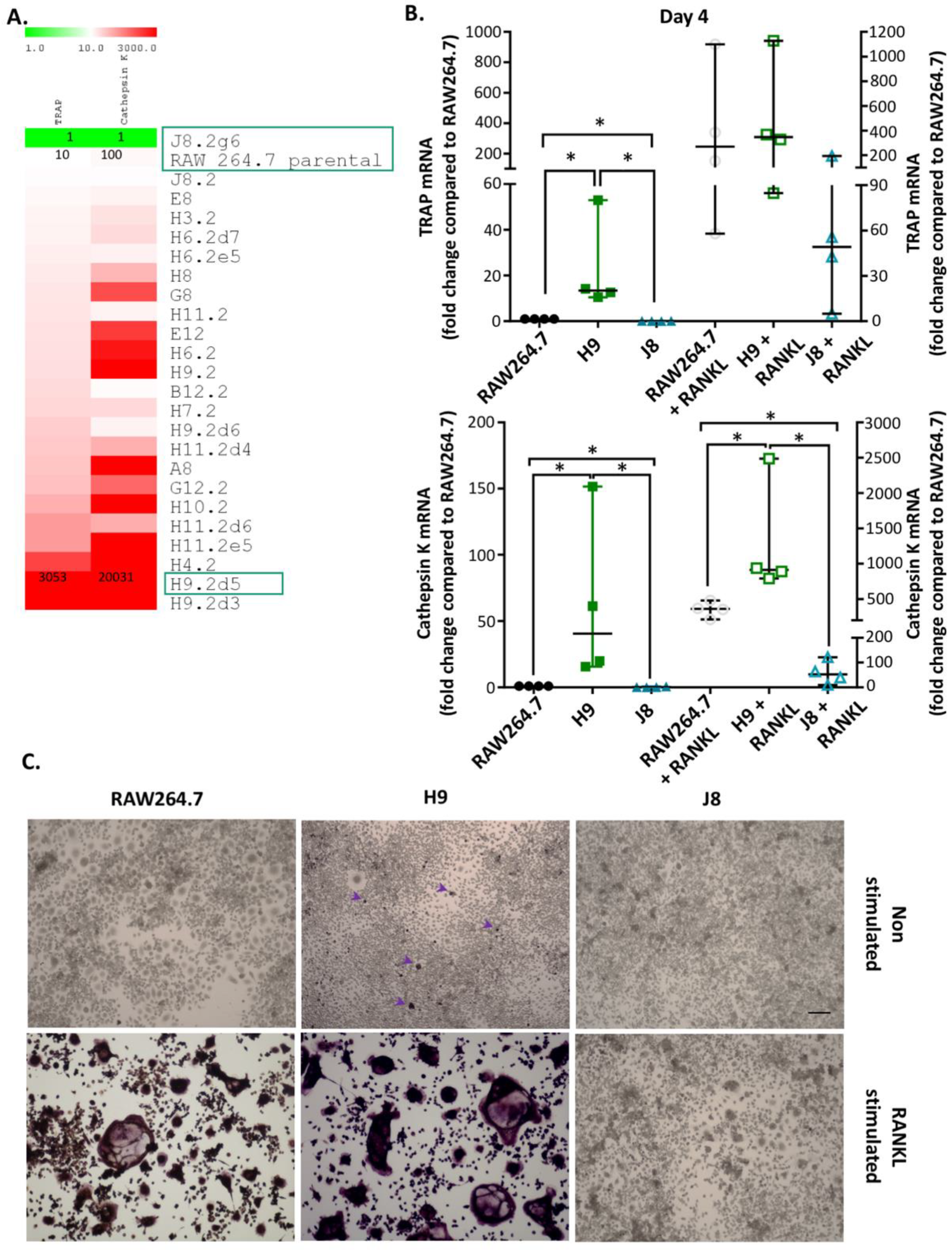
2.1. Selection of RAW264.7 Sub-Clones and Gene Expression of OC Markers, TRAP and Cathepsin K
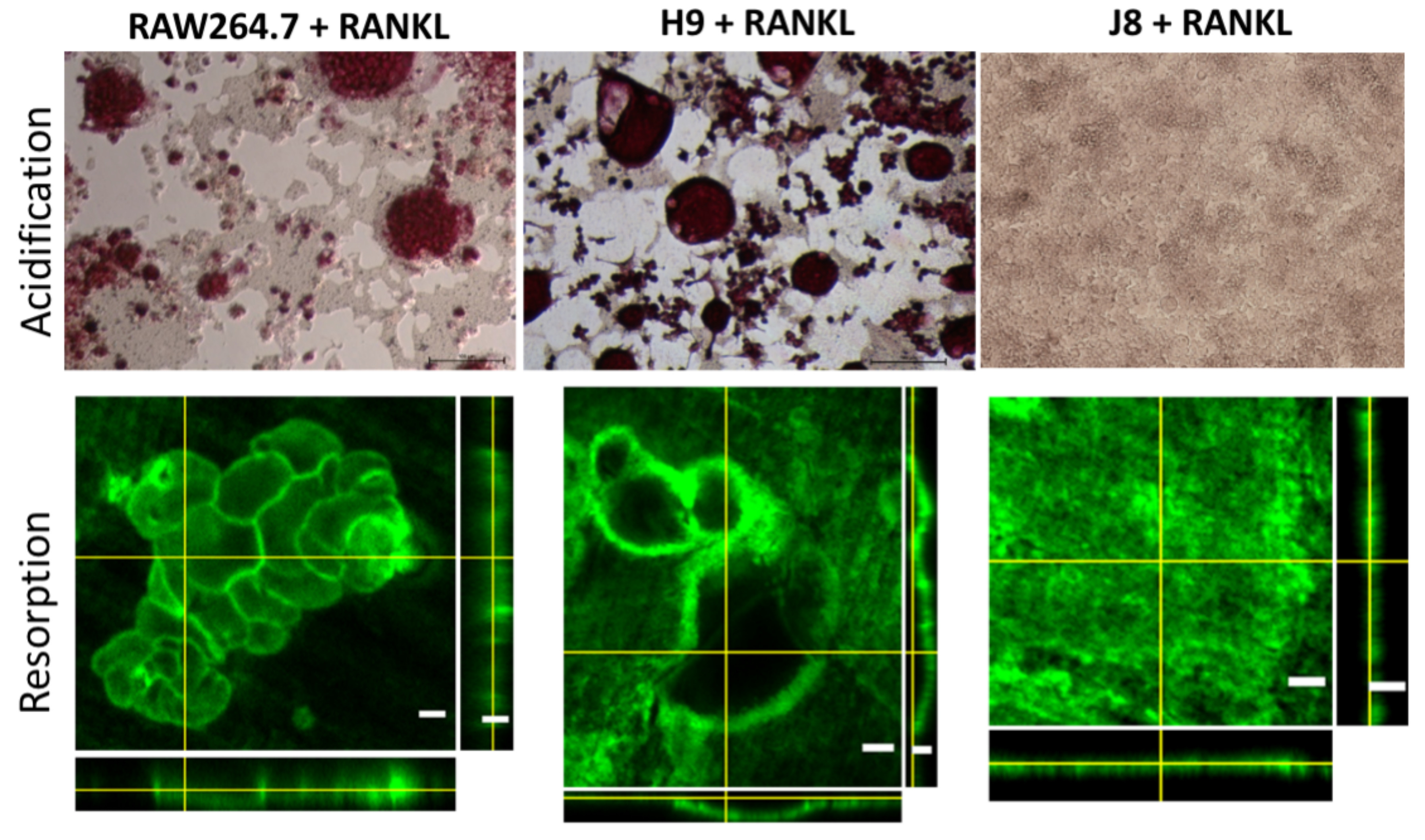
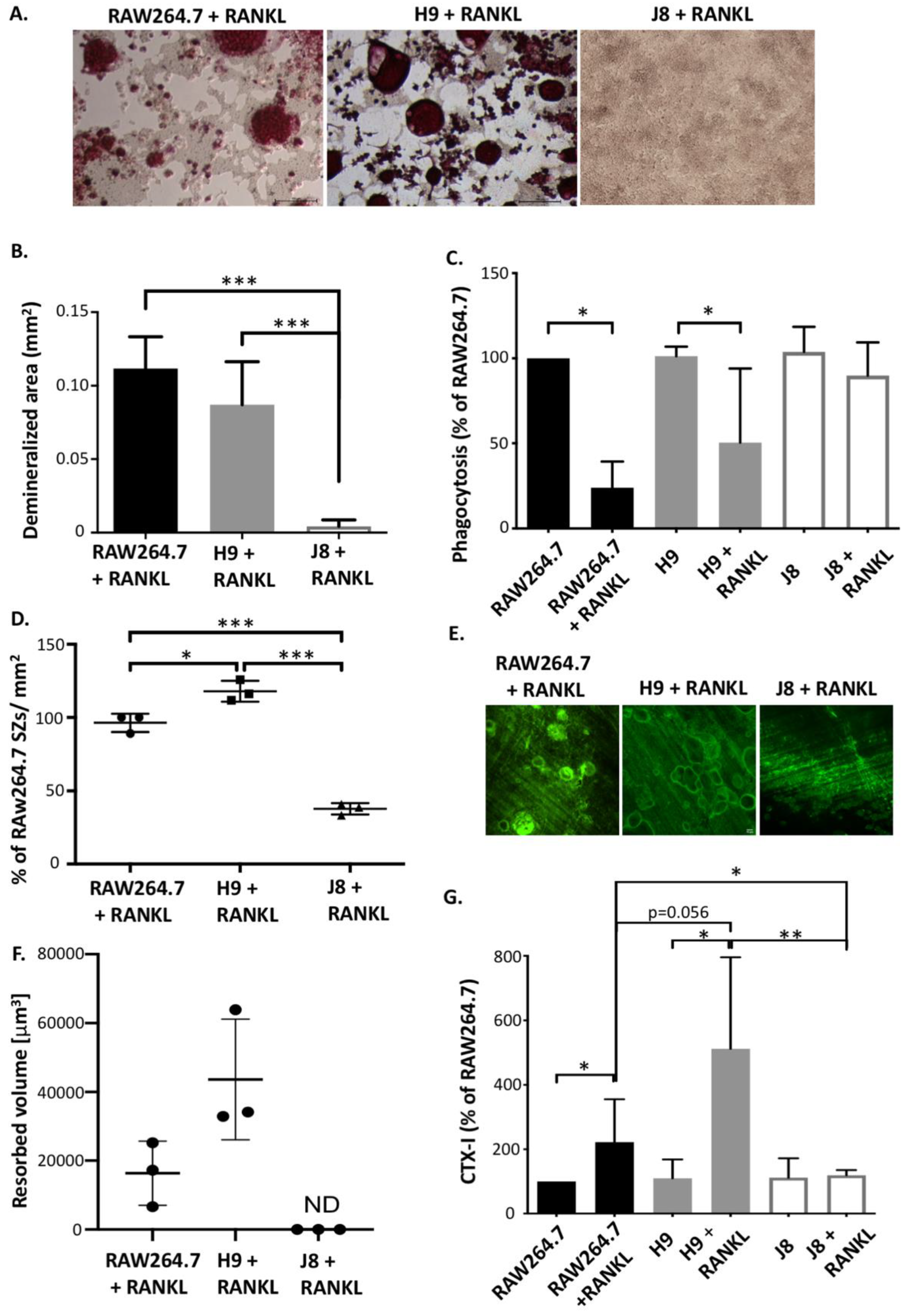
2.2. RANKL-Stimulated H9 and RAW264.7 Form Resorbing Osteoclast-Like Cells While J8 Does Not
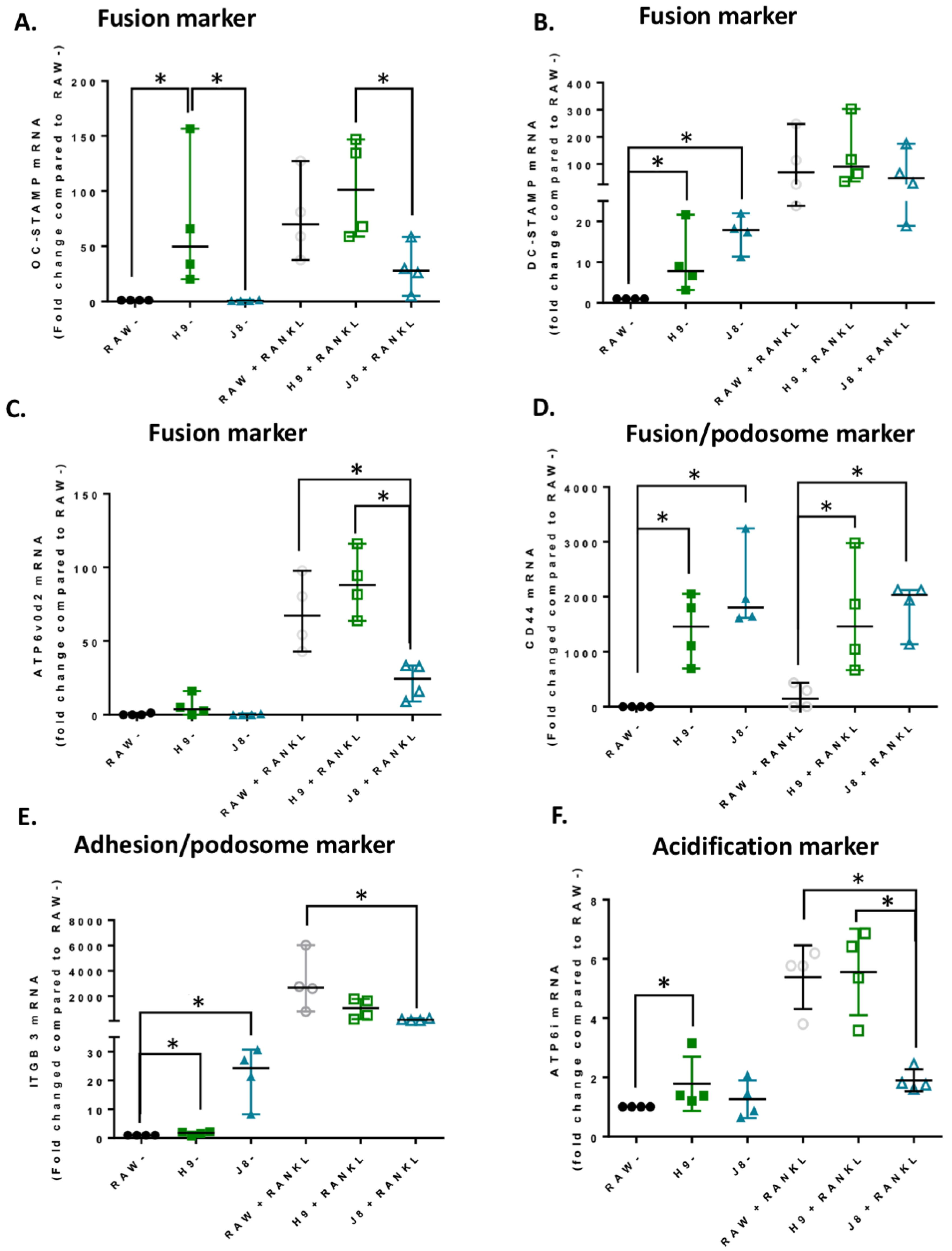
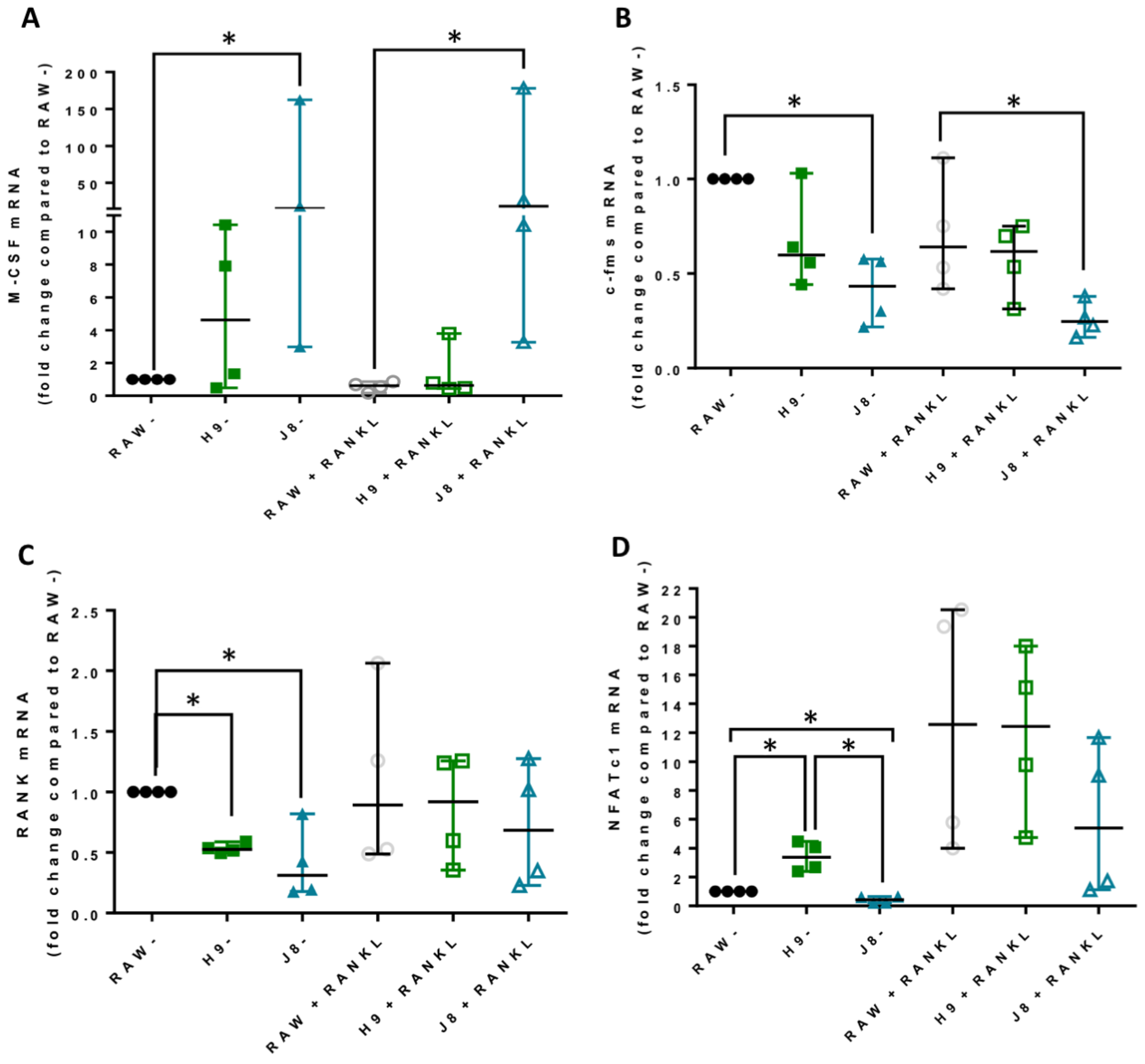
2.3. Gene Expression of Specific Osteoclast Markers Are Elevated in Unstimulated H9 Compared to RAW264.7
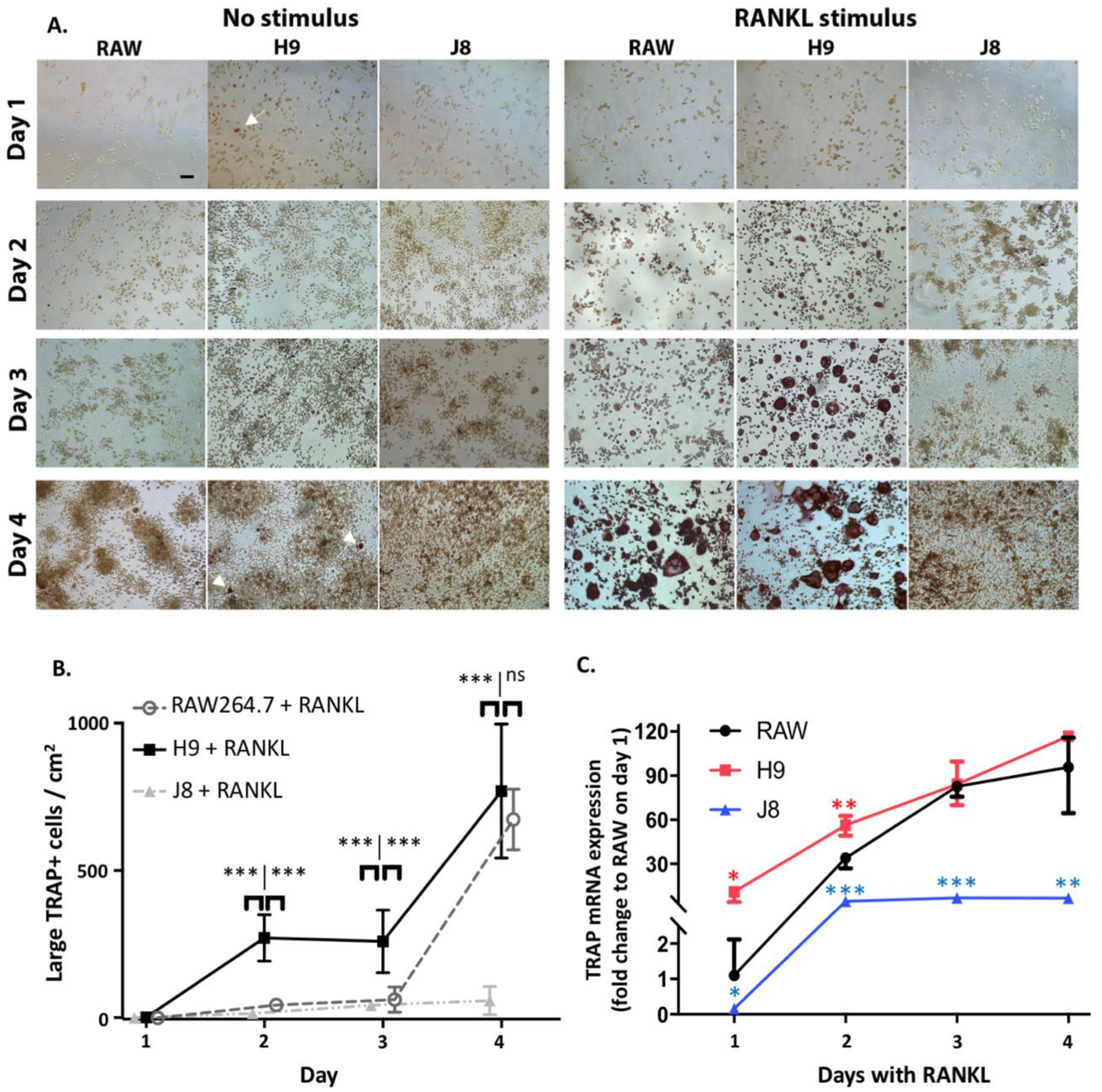
2.4. H9 Forms TRAP-Positive OCs Cells Faster than Parental RAW264.7 Due to Faster Increase in TRAP-Gene Expression
2.5. Unstimulated H9 Displays Higher Gene Expression of OC Transcription Factor NFATc1 Compared to RAW264.7
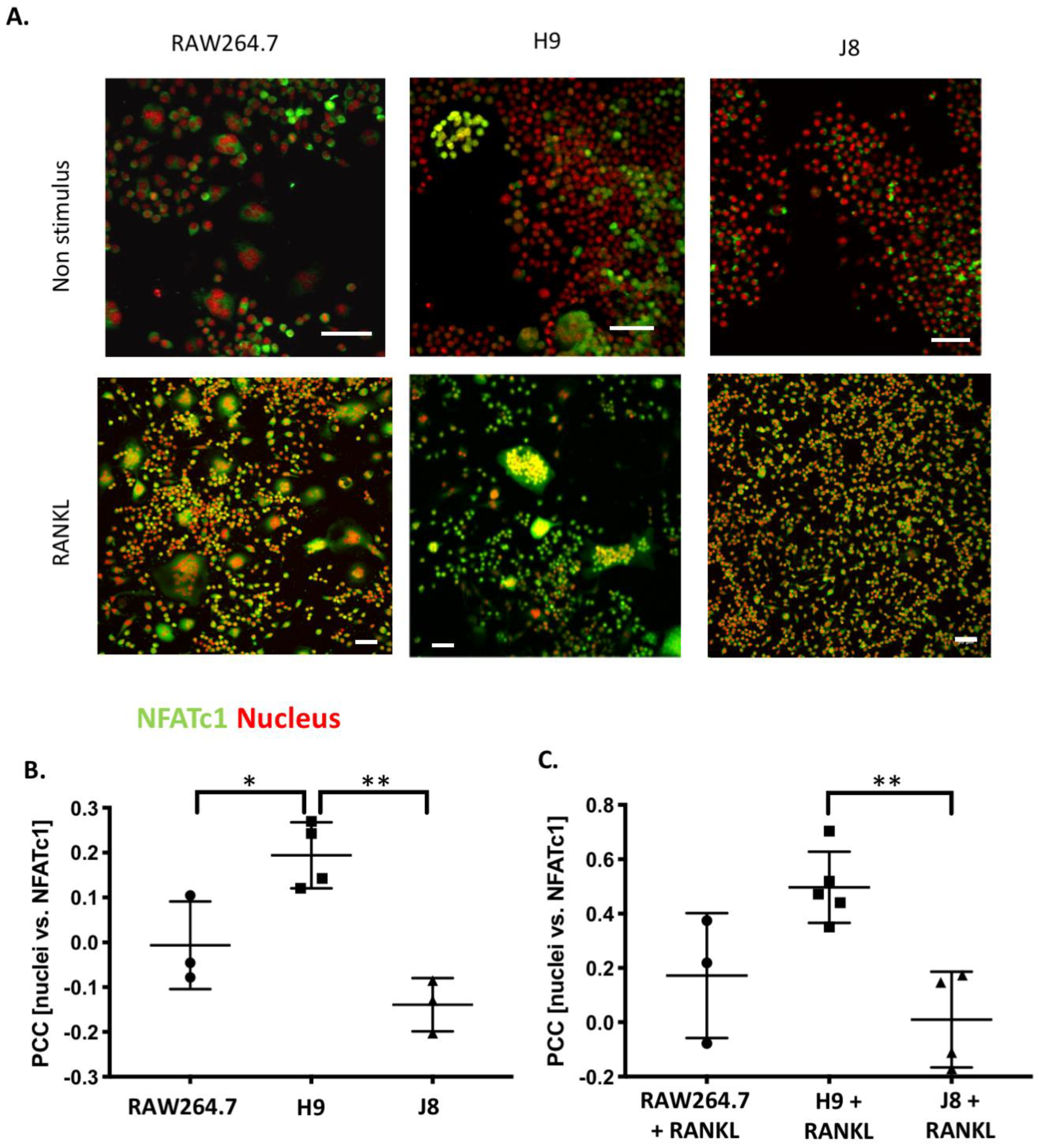
2.6. H9 Has Higher Degree of Nuclear Translocation of OC Transcription Factor NFATc1 than RAW264.7
3. Discussion and Conclusions
4. Material and Methods
4.1. Cell Culture
4.2. RAW264.7 Single Cell Cloning
4.3. RANKL Stimulation
4.4. RNA Purification and Reverse Transcription
4.5. Real Time qPCR
4.6. TRAP Staining of OC Cultures
4.7. Demineralization Assay
4.8. Phagocytosis Assay
4.9. Immunocytochemistry
4.10. Bone Chips
4.11. Bisphosphonate Staining and Resorption Pit Imaging
4.12. Measurement of Type I Collagen Degradation Marker (CTX-I)
4.13. Transmission Electron Microscopy (TEM)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cappariello, A.; Maurizi, A.; Veeriah, V.; Teti, A. The Great Beauty of the osteoclast. Arch. Biochem. Biophys. 2014, 558, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.F.; Aliprantis, A.O. Osteoclasts: More than ‘bone eaters’. Trends Mol. Med. 2014, 20, 449–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikawa, Y.; Quinn, J.M.; Sabokbar, A.; McGee, J.O.; Athanasou, N.A. The human osteoclast precursor circulates in the monocyte fraction. Endocrinology 1996, 137, 4058–4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, J.M.W.; Neale, S.; Fujikawa, Y.; McGee, J.O.D.; Athanasou, N.A. Human osteoclast formation from blood monocytes, peritoneal macrophages, and bone marrow cells. Calcif. Tissue Int. 1998, 62, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.M.W.; Elliott, J.; Gillespie, M.T.; Martin, T.J. A Combination of Osteoclast Differentiation Factor and Macrophage-Colony Stimulating Factor Is Sufficient for both Human and Mouse Osteoclast Formation in Vitro. Endocrinology 1998, 139, 4424–4427. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, Y.P. Generation of mouse osteoclastogenic cell lines immortalized with SV40 large T antigen. J. Bone Miner. Res. 1998, 13, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama, H.; Masuda, R.; Inoue, N.; Yamamoto, K.; Kuriiwa, K.; Nakagawa, K.; Yoshida, K. Establishment and characterization of macrophage-like cell lines expressing osteoclast-specific markers. J. Bone Miner. Metab. 2001, 19, 220–227. [Google Scholar] [CrossRef]
- Di Ceglie, I.; van den Akker, G.G.H.; Ascone, G.; ten Harkel, B.; Häcker, H.; van de Loo, F.A.J.; Koenders, M.I.; van der Kraan, P.M.; de Vries, T.J.; Vogl, T.; et al. Genetic modification of ER-Hoxb8 osteoclast precursors using CRISPR/Cas9 as a novel way to allow studies on osteoclast biology. J. Leukoc. Biol. 2017, 101, 957–966. [Google Scholar] [CrossRef]
- Raschke, W.C.; Baird, S.; Ralph, P.; Nakoinz, I. Functional macrophage cell lines transformed by Abelson leukemia virus. Cell 1978, 15, 261–267. [Google Scholar] [CrossRef]
- Nakagawa, N.; Kinosaki, M.; Yamaguchi, K.; Shima, N.; Yasuda, H.; Yano, K.; Morinaga, T.; Higashio, K. RANK is the essential signaling receptor for osteoclast differentiation factor in osteoclastogenesis. Biochem. Biophys. Res. Commun. 1998, 253, 395–400. [Google Scholar] [CrossRef]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A.; et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Hayashi, S.; Kunisada, T.; Ogawa, M.; Nishikawa, S.; Okamura, H.; Sudo, T.; Shultz, L.D. The murine mutation osteopetrosis is in the coding region of the macrophage colony stimulating factor gene. Nature 1990, 345, 442–444. [Google Scholar] [CrossRef]
- Wiktor-Jedrzejczak, W.; Bartocci, A.; Ferrante, A.W.; Ahmed-Ansari, A.; Sell, K.W.; Pollard, J.W.; Stanley, E.R. Total absence of colony-stimulating factor 1 in the macrophage-deficient osteopetrotic (op/op) mouse. Proc. Natl. Acad. Sci. USA 1990, 87, 4828–4832. [Google Scholar] [CrossRef] [Green Version]
- Felix, R.; Cecchini, M.G.; Fleisch, H. Macrophage colony stimulating factor restores in vivo bone resorption in the op/op osteopetrotic mouse. Endocrinology 1990, 127, 2592–2594. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, N. Regulation of NFATc1 in Osteoclast Differentiation. J. Bone Metab. 2014, 21, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Takahasi, N.; Maeda, K.; Ishihara, A.; Uehara, S.; Kobayashi, Y. Regulatory Mechanism of Osteoclastogenesis by RANKL and Wnt Signals. Front. Biosci. 2011, 16, 1–13. [Google Scholar]
- Collin-Osdoby, P.; Osdoby, P. RANKL-Mediated Osteoclast Formation from Murine RAW 264.7 cells. In Bone Research Protocols; Helfrich, M.H., Ralston, S.H., Eds.; Springer: New York, NY, USA, 2012; pp. 187–202. [Google Scholar]
- Taciak, B.; Białasek, M.; Braniewska, A.; Sas, Z.; Sawicka, P.; Kiraga, Ł.; Rygiel, T.; Kro, M. Evaluation of phenotypic and functional stability of RAW 264. 7 cell line through serial passages. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Masters, J.R.W. Human cancer cell lines: Fact and fantasy. Nat. Rev. Mol. Cell Biol. 2000, 1, 233–236. [Google Scholar] [CrossRef]
- Thompson, C.D.; Frazier-Jessen, M.R.; Rawat, R.; Nordan, R.P.; Brown, R.T. Evaluation of methods for transient transfection of a murine macrophage cell line, RAW 264.7. Biotechniques 1999, 27, 824–832. [Google Scholar] [CrossRef]
- Cuetara, B.L.V.; Crotti, T.N.; O’Donoghue, A.J.; McHugh, K.P. Cloning and characterization of osteoclast precursors from the RAW264.7 cell line. In Vitro. Cell Dev. Biol. Anim. 2000, 42, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, A.Y.; Tu, C.; Shen, S.; Xu, D.; Oursler, M.J.; Qu, J.; Yang, S. Comparative Characterization of Osteoclasts Derived From Murine Bone Marrow Macrophages and RAW 264.7 Cells Using Quantitative Proteomics. JBMR Plus 2018, 2, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Smith, W.; Hao, D. Overview of RAW264.7 for osteoclastogensis study: Phenotype and stimuli. J. Cell Mol. Med. 2019, 23, 3077–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgess, D.; Machuca-Gayet, I.; Blangy, A.; Jurdic, P. Podosome organization drives osteoclast-mediated bone resorption. Cell Adh. Migr. 2014, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabadel, A.; Banon-Rodrıguez, I.; Cluet, D.; Rudkin, B.B.; Wehrle-Haller, B.; Genot, E.; Pierre, J.; Anton, I.M.; Saltel, F. CD44 and B3 Integrin Organize Two Functionally Distinct Actin-based Domains in Osteoclasts. Mol. Biol. Cell 2008, 19, 2609–2619. [Google Scholar]
- Xing, L. Osteoclast fusion and regulation by RANKL-dependent and independent factors. World J. Orthop. 2012, 3, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Chen, W.; Liang, Y.; Li, E.; Stashenko, P. Atp6i-deficient mice exhibit severe osteopetrosis due to loss of osteoclast-mediated extracellular acidification. Nat. Genet. 1999, 23, 447–451. [Google Scholar] [CrossRef]
- Hume, D.A.; Yue, X.; Ross, I.L.; Favot, P.; Lichanska, A.; Ostrowski, M.C. Regulation of CSF-1 Receptor Expression. Mol. Reprod. Dev. 1997, 83, 46–53. [Google Scholar] [CrossRef]
- Destaing, O.; Saltel, F.; Geminard, J.C.; Jurdic, P.; Bard, F. Podosomes display actin turnover and dynamic self-organization in osteoclasts expressing actin-green fluorescent protein. Mol. Biol. Cell 2003, 14, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Pavlos, N.J.; Xu, J.; Riedel, D.; Yeoh, J.S.G.; Teitelbaum, S.L.; Papadimitriou, J.M.; Jahn, R.; Ross, F.P.; Zheng, M.H. Rab3D Regulates a Novel Vesicular Trafficking Pathway That Is Required for Osteoclastic Bone Resorption. Mol. Cell Biol. 2005, 25, 5253–5269. [Google Scholar] [CrossRef] [Green Version]
- Hollberg, K.; Marsell, R.; Norgård, M.; Larsson, T.; Jonsson, K.B.; Andersson, G. Osteoclast polarization is not required for degradation of bone matrix in rachitic FGF23 transgenic mice. Bone 2008, 42, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Hollberg, K.; Nordahl, J.; Hultenby, K.; Mengarelli-Widholm, S.; Andersson, G.; Reinholt, F.P. Polarization and secretion of cathepsin K precede tartrate-resistant acid phosphatase secretion to the ruffled border area during the activation of matrix-resorbing clasts. J. Bone Min. Metab. 2005, 23, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Saftig, P.; Hunziker, E.; Wehmeyer, O.; Jones, S.; Boyde, A.; Rommerskirch, W.; Moritz, J.D.; Schu, P.; von Figura, K. Impaired osteoclastic bone resorption leads to osteopetrosis in cathepsin-K-deficient mice. Proc. Natl. Acad. Sci. USA 1998, 95, 13453–13458. [Google Scholar] [CrossRef] [Green Version]
- Lång, P.; Andersson, G. Differential expression of monomeric and proteolytically processed forms of tartrate-resistant acid phosphatase in rat tissues. Cell Mol. Life Sci. 2005, 62, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Lång, P.; van Harmelen, V.; Rydén, M.; Kaaman, M.; Parini, P.; Carneheim, C.; Cassady, A.I.; Hume, D.A.; Andersson, G.; Arner, P. Monomeric tartrate resistant acid phosphatase induces insulin sensitive obesity. PLoS ONE 2008, 3, e1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lång, P.; Lange, S.; Delbro, D.; Andersson, G. Induction and cellular expression of tartrate resistant acid phosphatase during dextran sodium sulphate induced colitis in rats. Histochem. Cell Biol. 2009, 132, 599–612. [Google Scholar] [CrossRef]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [Green Version]
- Abràmofff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ Part II. Biophotonics Int. 2005, 11, 36–43. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mira-Pascual, L.; Tran, A.N.; Andersson, G.; Näreoja, T.; Lång, P. A Sub-Clone of RAW264.7-Cells Form Osteoclast-Like Cells Capable of Bone Resorption Faster than Parental RAW264.7 through Increased De Novo Expression and Nuclear Translocation of NFATc1. Int. J. Mol. Sci. 2020, 21, 538. https://doi.org/10.3390/ijms21020538
Mira-Pascual L, Tran AN, Andersson G, Näreoja T, Lång P. A Sub-Clone of RAW264.7-Cells Form Osteoclast-Like Cells Capable of Bone Resorption Faster than Parental RAW264.7 through Increased De Novo Expression and Nuclear Translocation of NFATc1. International Journal of Molecular Sciences. 2020; 21(2):538. https://doi.org/10.3390/ijms21020538
Chicago/Turabian StyleMira-Pascual, Laia, Anh N. Tran, Göran Andersson, Tuomas Näreoja, and Pernilla Lång. 2020. "A Sub-Clone of RAW264.7-Cells Form Osteoclast-Like Cells Capable of Bone Resorption Faster than Parental RAW264.7 through Increased De Novo Expression and Nuclear Translocation of NFATc1" International Journal of Molecular Sciences 21, no. 2: 538. https://doi.org/10.3390/ijms21020538
APA StyleMira-Pascual, L., Tran, A. N., Andersson, G., Näreoja, T., & Lång, P. (2020). A Sub-Clone of RAW264.7-Cells Form Osteoclast-Like Cells Capable of Bone Resorption Faster than Parental RAW264.7 through Increased De Novo Expression and Nuclear Translocation of NFATc1. International Journal of Molecular Sciences, 21(2), 538. https://doi.org/10.3390/ijms21020538