Differences in Mitochondrial Membrane Potential Identify Distinct Populations of Human Cardiac Mesenchymal Progenitor Cells

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
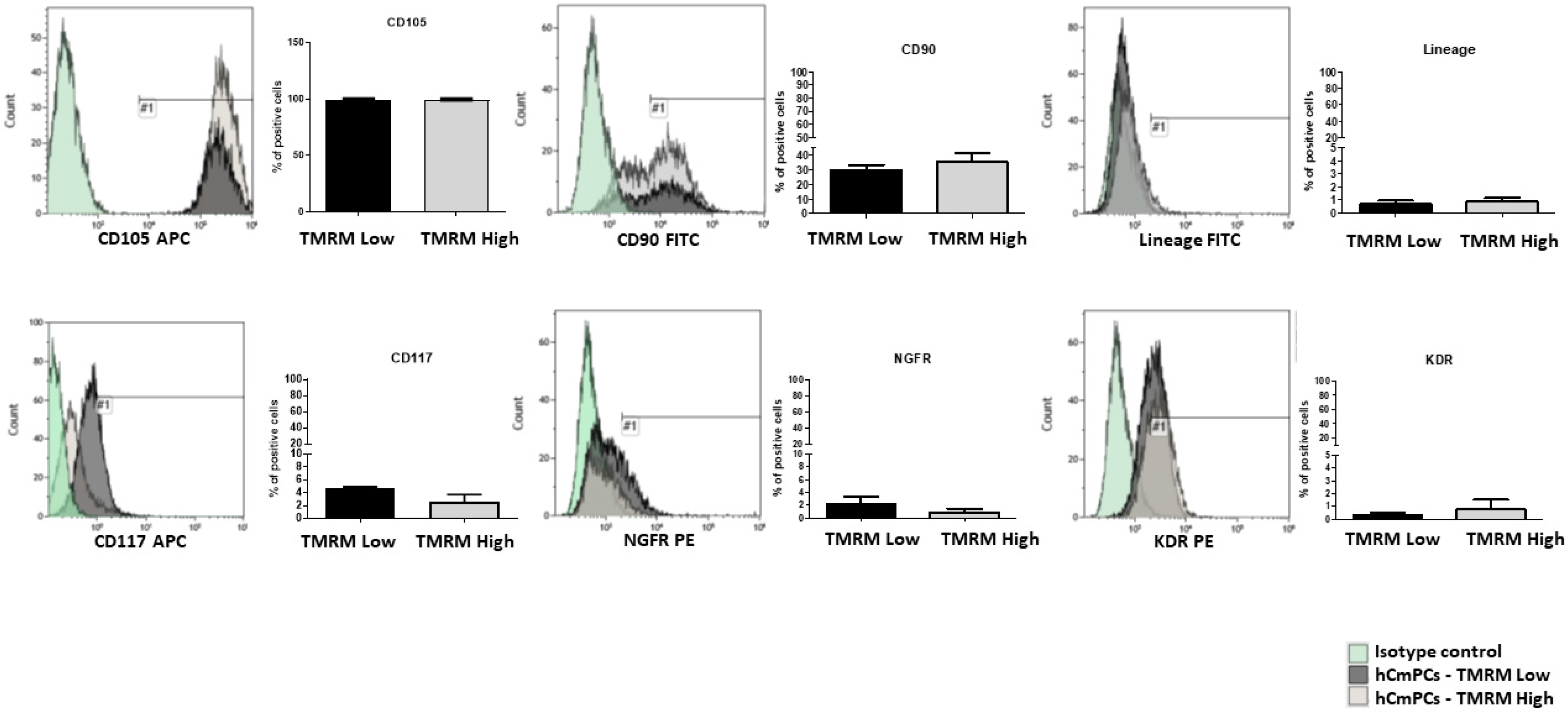
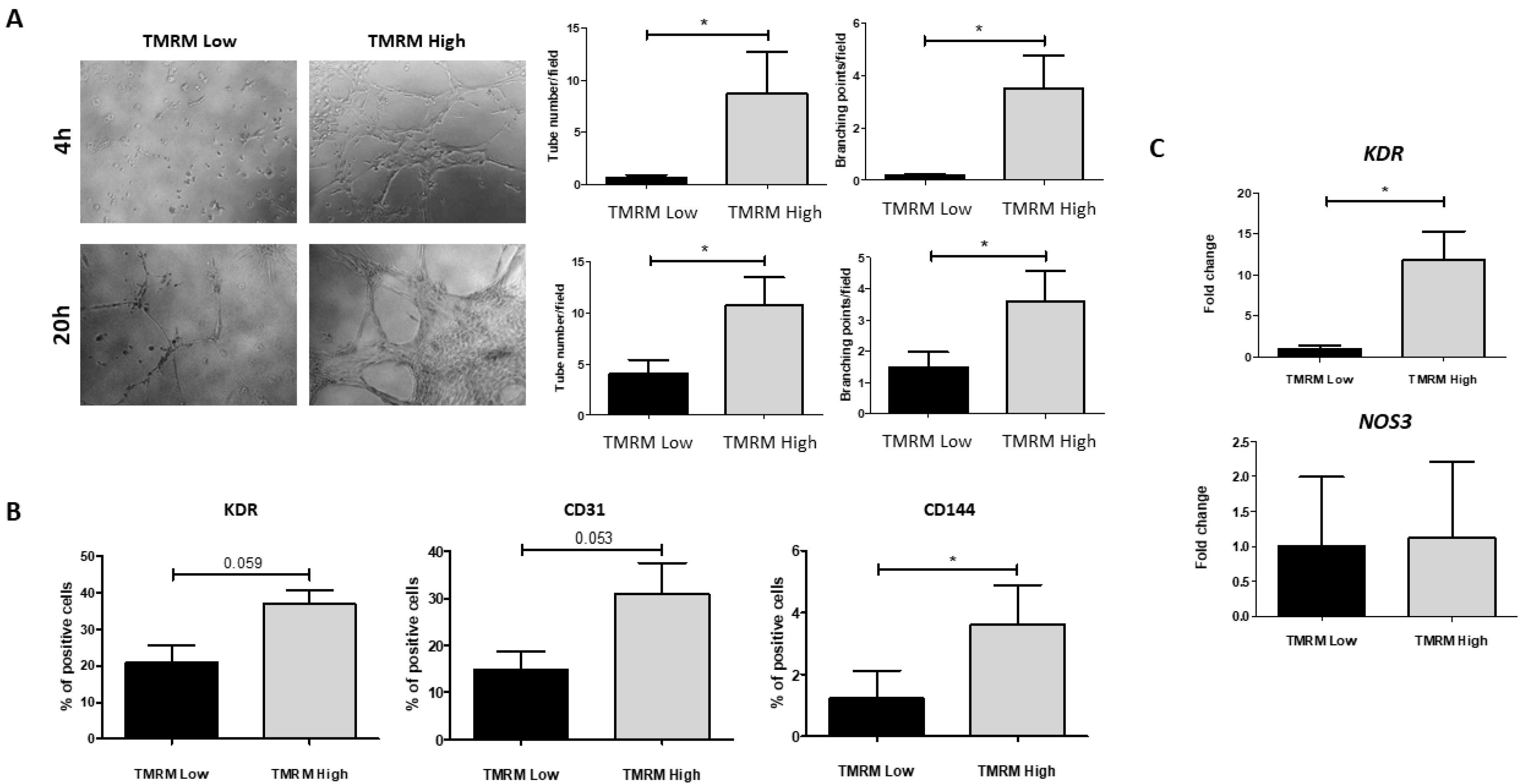
2.1. Surface Marker Expression
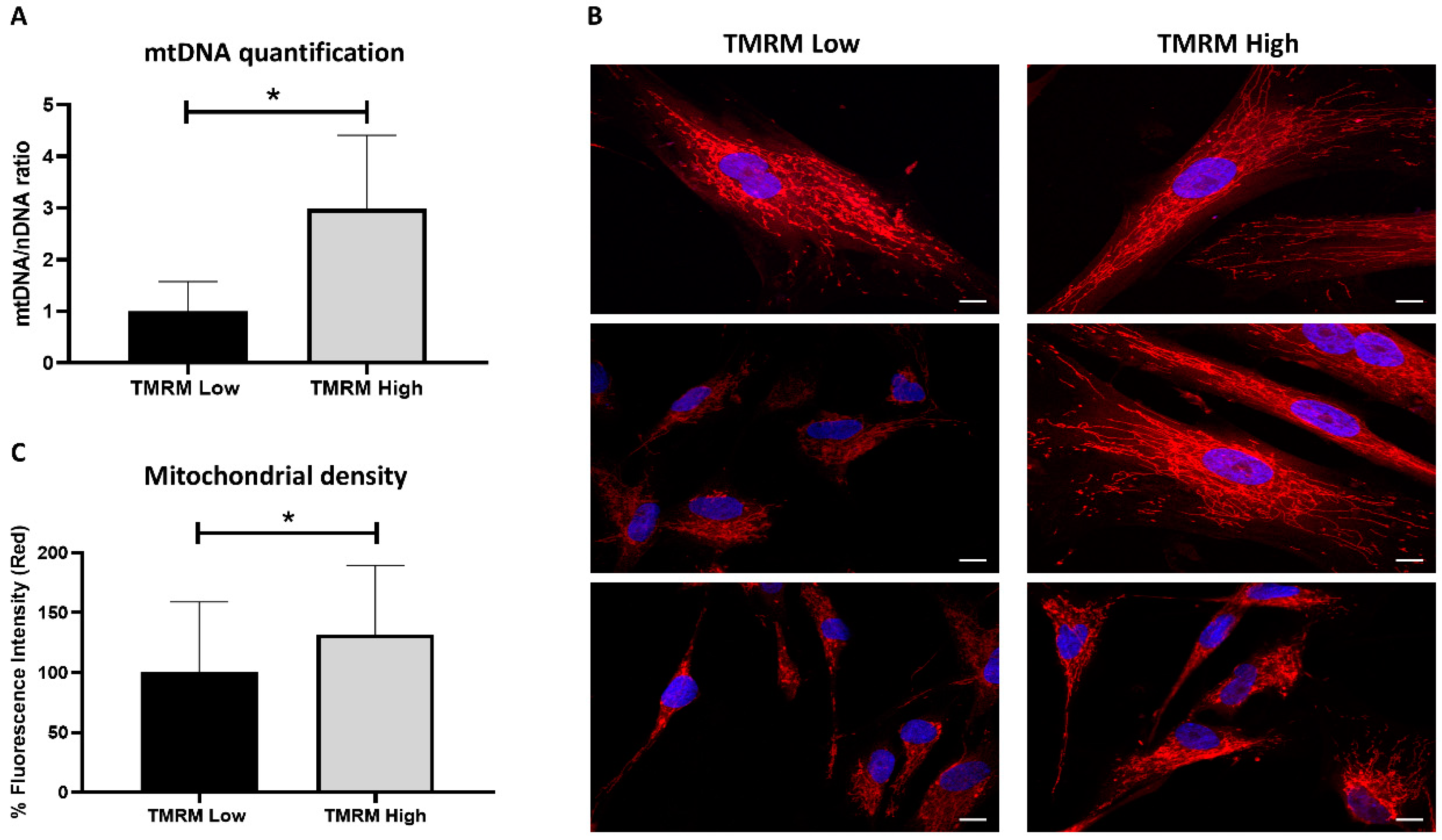
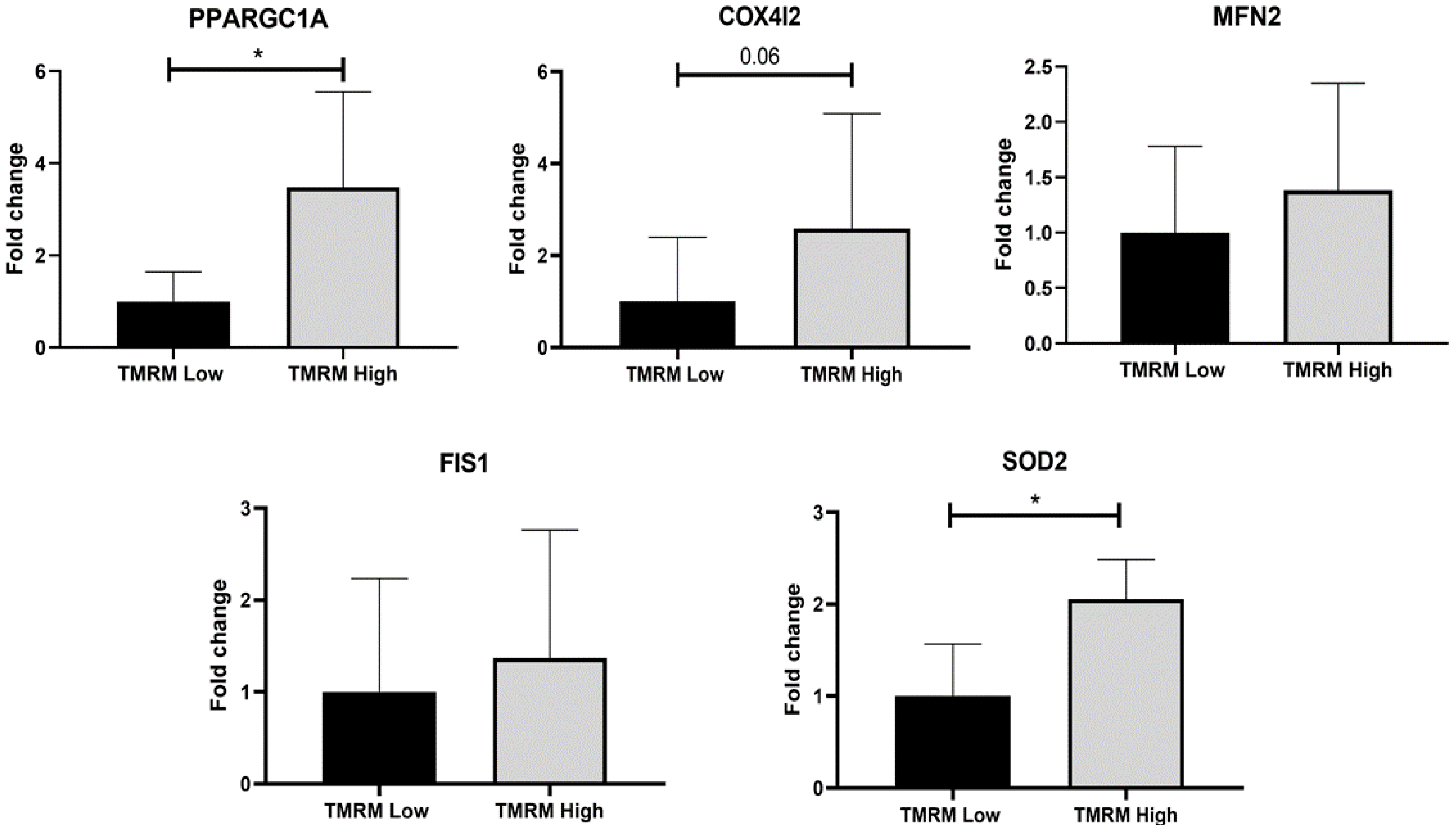
2.2. Energy Metabolism
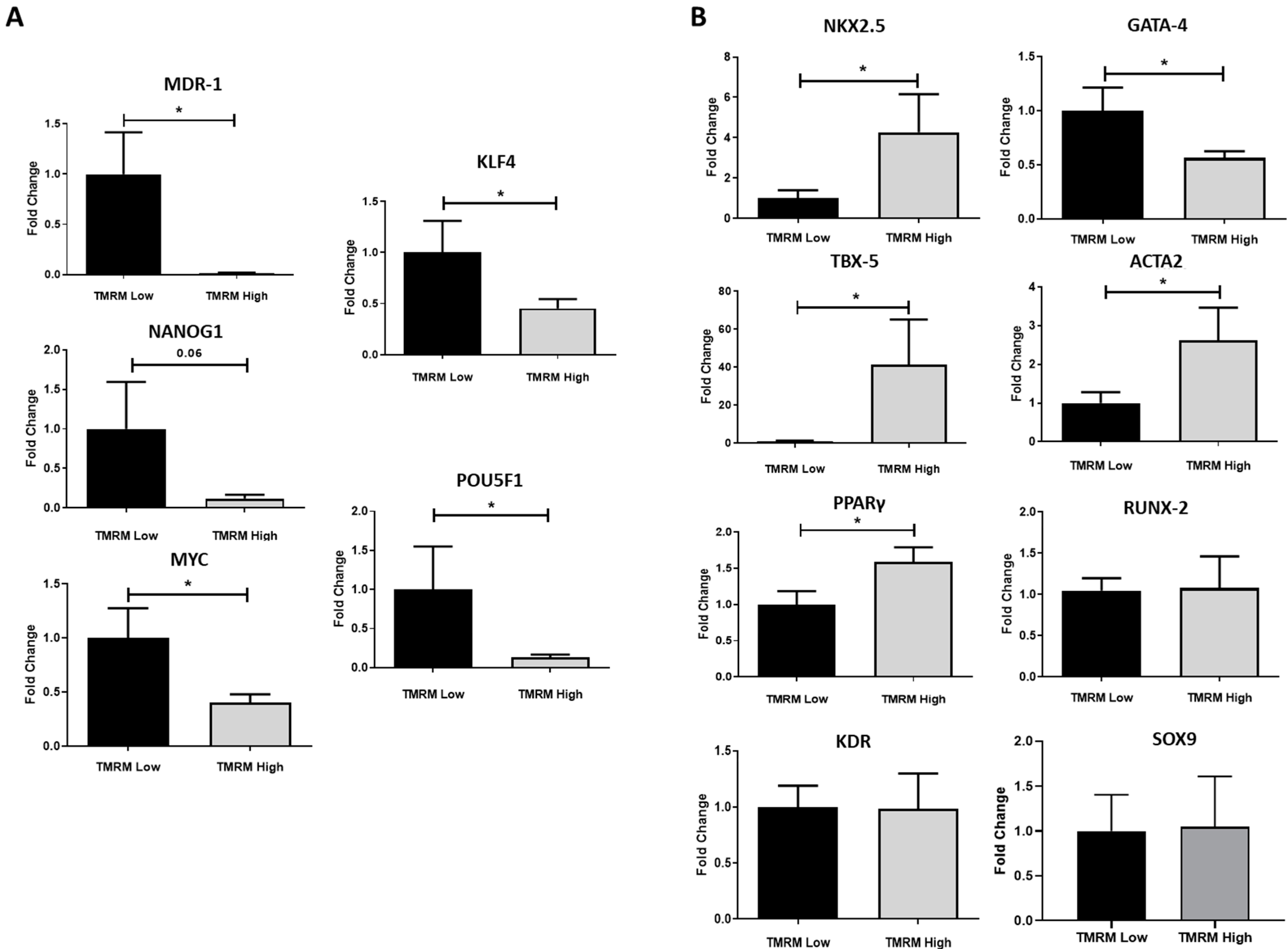
2.3. Gene Expression in Basal Conditions
2.4. Multilineage Cardiovascular and Mesenchymal Commitment
2.4.1. Endothelial Differentiation
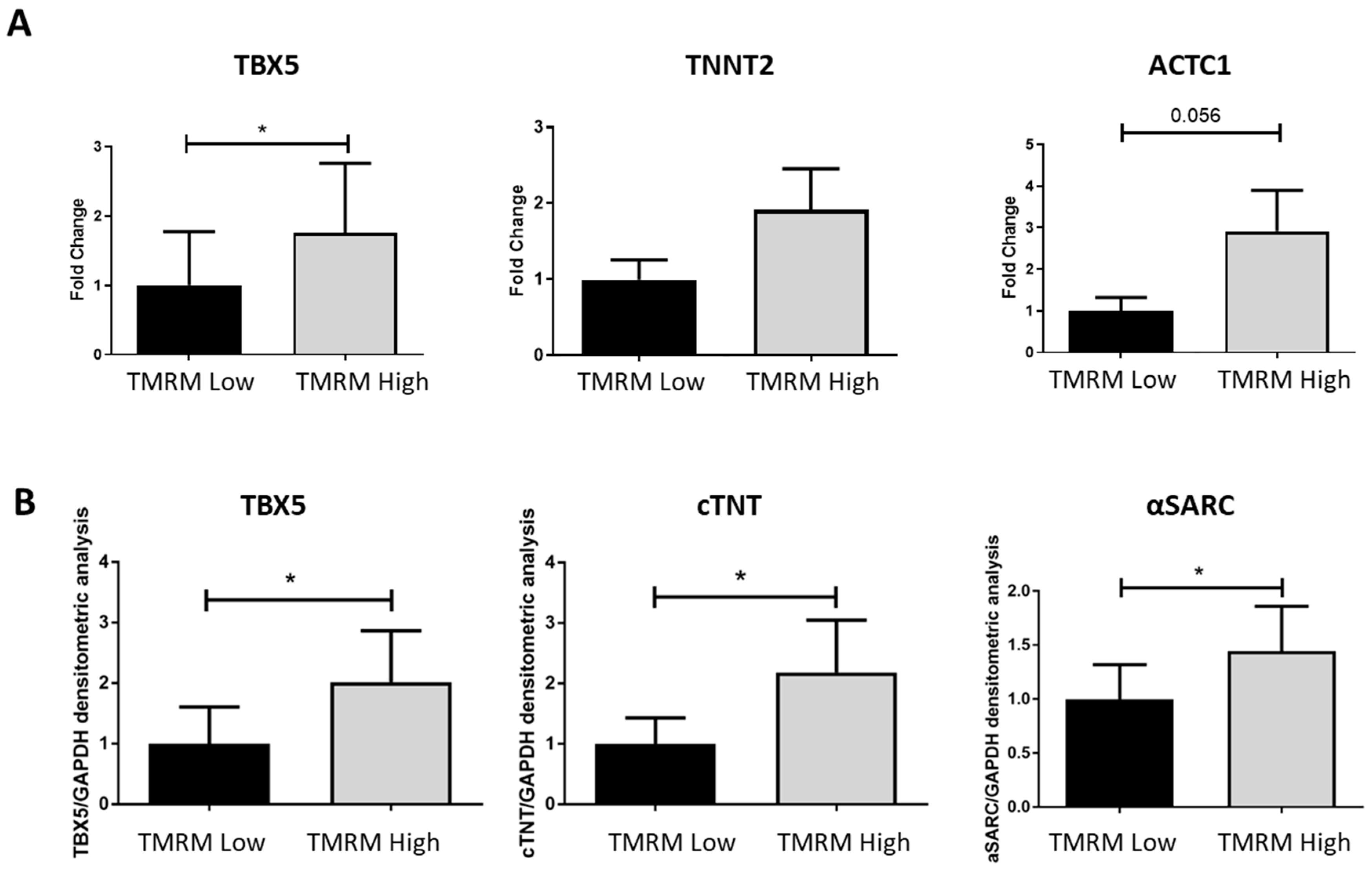
2.4.2. Cardiomyogenic Differentiation
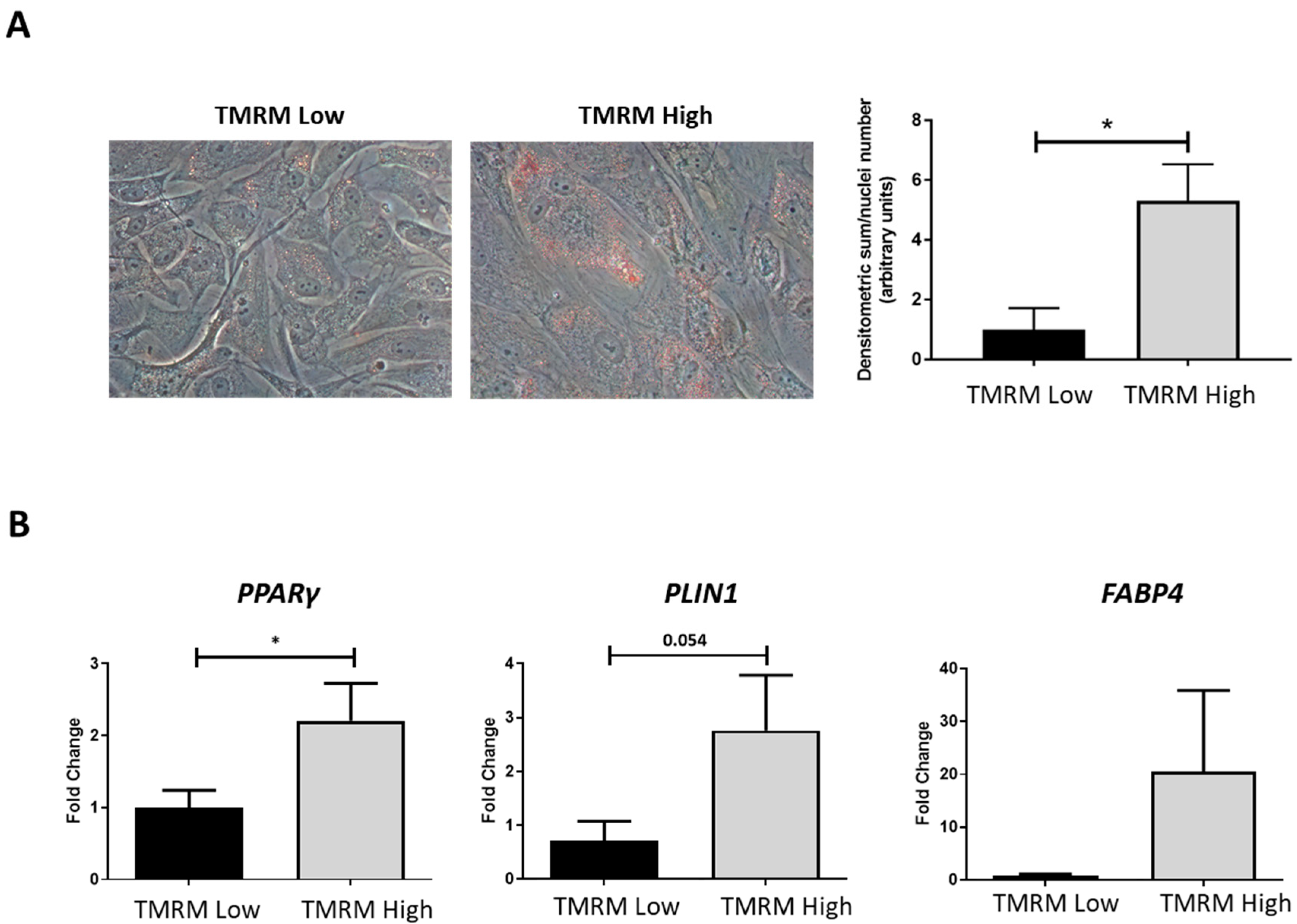
2.4.3. Adipogenic Differentiation
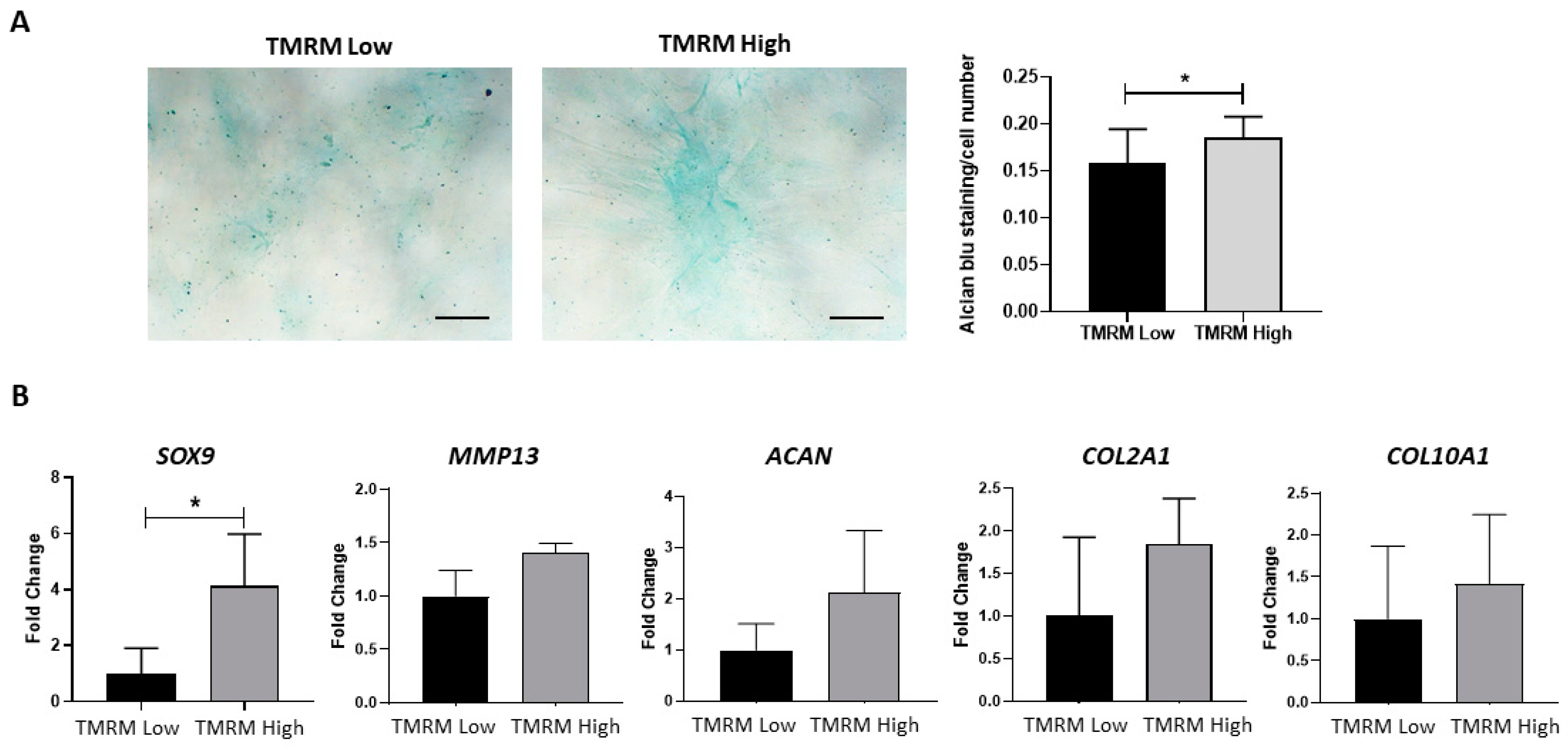
2.4.4. Chondrogenic Differentiation
2.4.5. Osteogenic Differentiation
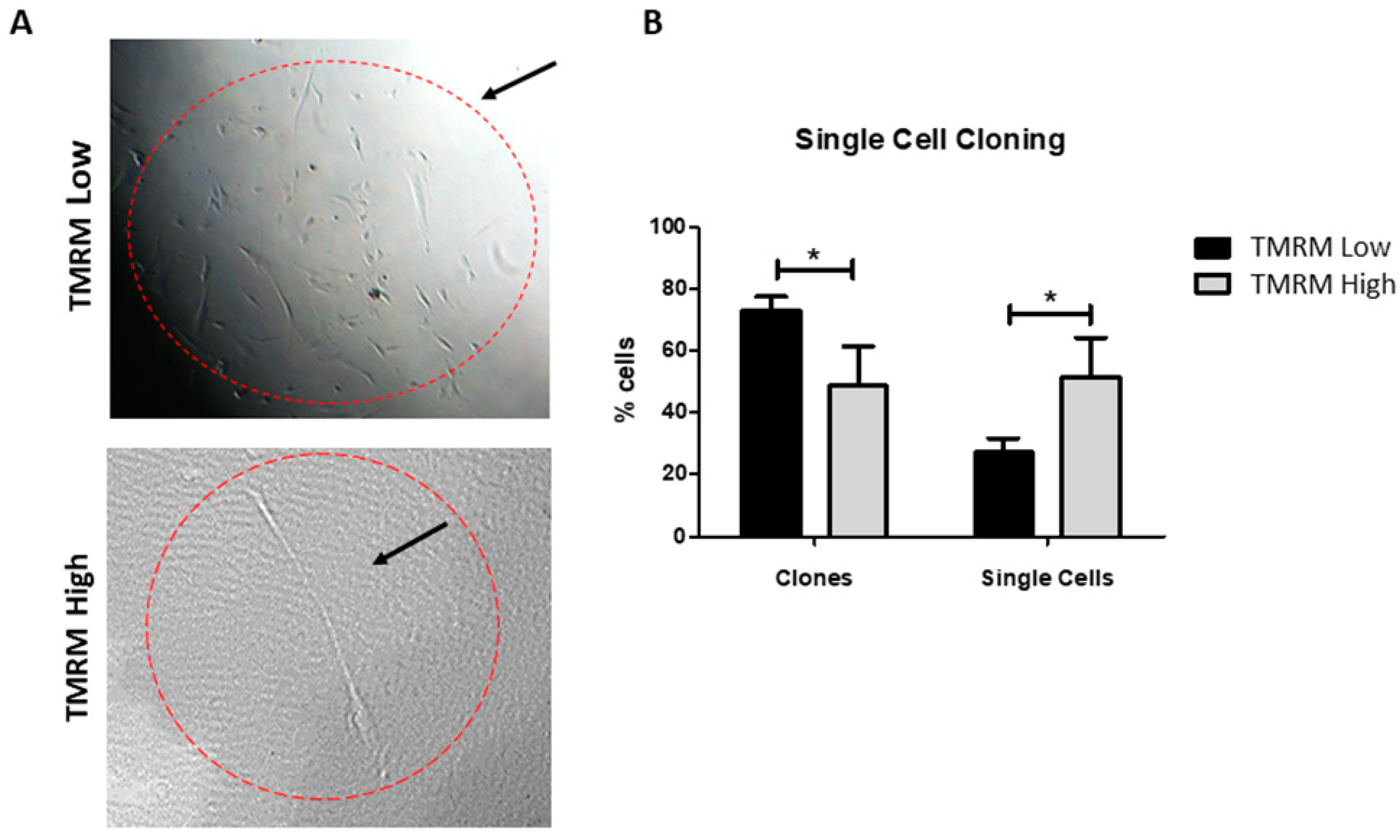
2.5. Single Cell Cloning
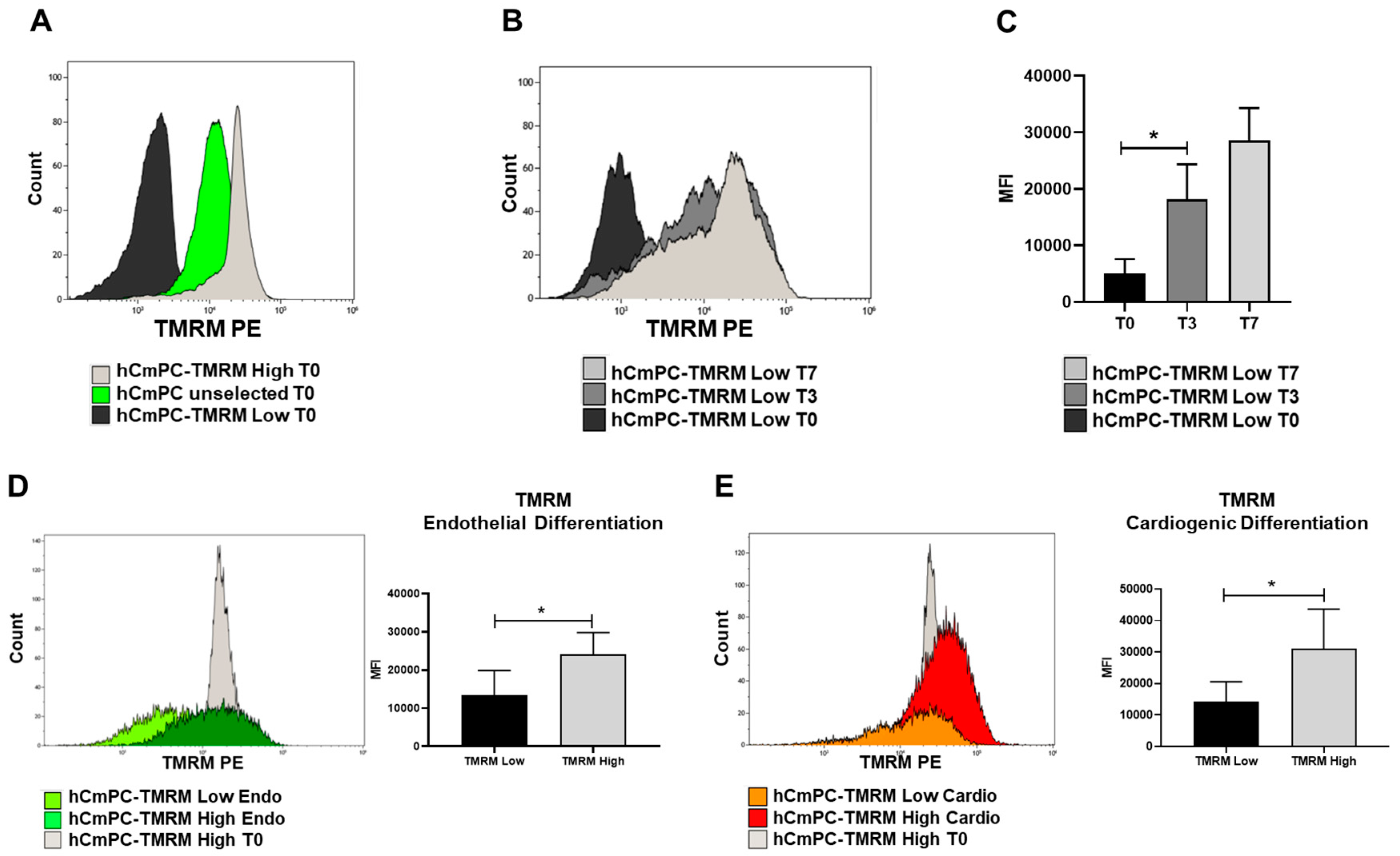
2.6. TMRM Transition in Culture and During Differentiation
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Cell Culture and TMRM Sorting
4.3. FACS Analysis
4.4. Seahorse Extracellular Flux (XF) Cell Mito Stress Test
4.5. Mitochondrial DNA Quantification
4.6. Mitochondrial Network
4.7. Gene Expression Analysis
4.8. Single Cell Cloning
4.9. Endothelial Differentiation
4.10. Cultrex Assays
4.11. Cardiac Differentiation
4.12. Adipogenic Differentiation
4.13. Oil Red O Staining
4.14. Chondrogenic Differentiation
4.15. Alcian Blue Staining
4.16. Osteogenic Differentiation
4.17. von Kossa Staining
4.18. Crystal Violet Staining
4.19. Western Blot
4.20. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACAN | Aggrecan |
| ATP | Adenosine triphosphate |
| B2M | β2 microglobulin |
| BM | Bone marrow |
| c-Myc | MYC |
| COL2A1 | Collagen type II alpha 1 chain |
| COL10A1 | Collagen type X alpha 1 chain |
| COX4I2 | Cytochrome c oxidase subunit 4I2 |
| CPCs | Cardiac progenitor cells |
| CSCs | Cardiac stem cells |
| cTNT | Cardiac troponin T |
| ECAR | Extracellular acidification rate |
| ESCs | Embryonic stem cells |
| FABP4 | Fatty acid-binding protein 4 |
| FACS | Fluorescent activated cell sorting |
| FCCP | Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone |
| FIS1 | Mitochondrial fission 1 protein |
| GATA4 | GATA Binding protein 4 |
| hCmPC | Human cardiac mesenchymal progenitor cells |
| iPSCs | Induced pluripotent stem cells |
| KDR | kinase insert domain receptor |
| KLF4 | Krüppel like factor 4 |
| MDR-1 | Multidrug-resistance |
| MFN2 | Mitofusin-2 |
| MFIM | Mitofusin-2Mean fluorescence intensity |
| MP13 | Matrix metallopeptidase 13 |
| mtDNA | Mitochondrial Deoxyribonucleic Acid |
| ND5 | Mitochondrially encoded NADH, ubiquinone oxidoreductase core subunit 5 |
| nDNA | Nuclear Deoxyribonucleic Acid |
| NGFR | Nerve growth factor receptor |
| NKX2.5 | NK2 Homeobox 5 |
| NOS3 | Endothelial nitric oxide synthase |
| OCR | Oxygen consumption rate |
| Oct-4 | Octamer-binding transcription factor |
| Opa1 | Optic atrophy type 1 |
| OXPHOS | Oxidative phosphorylation |
| PCR | Polymerase chain reaction |
| PGC-1α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PLIN-1 | Perilipin 1 |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| ROS | Reactive oxygen species |
| RUNX-2 | Runt-related transcription factor 2 |
| SPP1 | Secreted Phosphoprotein 1 |
| SOD2 | Superoxide dismutase 2 |
| SOX9 | SRY-Box transcription factor 9 |
| TBX5 | T-Box transcription factor 5 |
| TMRM | Tetramethylrhodamine methyl ester |
| VEGFR-2 | Vascular endothelial growth factor receptor 2 |
| α-SARC | Alpha sarcomeric actin |
| α-SMA | Alpha-smooth muscle actin |
| Δψm | Mitochondrial membrane potential |
References
- Rurali, E.; Vinci, M.C.; Bassetti, B.; Barbagallo, V.; Pompilio, G.; Gambini, E. New Strategies to Enhance Myocardial Regeneration: Expectations and Challenges from Preclinical Evidence. Curr. Stem Cell Res. 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Colter, D.C.; Class, R.; DiGirolamo, C.M.; Prockop, D.J. Rapid expansion of recycling stem cells in cultures of plastic-adherent cells from human bone marrow. Proc. Natl. Acad. Sci. USA 2000, 97, 3213–3218. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J. Stromal mechanisms of bone marrow: Cloning in vitro and retransplantation in vivo. Haematol. Blood Transfus. 1980, 25, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Leri, A.; Kajstura, J.; Anversa, P.; Frishman, W.H. Myocardial regeneration and stem cell repair. Curr. Probl. Cardiol. 2008, 33, 91–153. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ramos, J.R. Neural cells derived from adult bone marrow and umbilical cord blood. J. Neurosci. Res. 2002, 69, 880–893. [Google Scholar] [CrossRef]
- Wakitani, S.; Imoto, K.; Yamamoto, T.; Saito, M.; Murata, N.; Yoneda, M. Human autologous culture expanded bone marrow mesenchymal cell transplantation for repair of cartilage defects in osteoarthritic knees. Osteoarthr. Cart. 2002, 10, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.A.; Kinsey, S.E.; English, A.; Jones, R.A.; Straszynski, L.; Meredith, D.M.; Markham, A.F.; Jack, A.; Emery, P.; McGonagle, D. Isolation and characterization of bone marrow multipotential mesenchymal progenitor cells. Arthritis Rheum. 2002, 46, 3349–3360. [Google Scholar] [CrossRef]
- Le Blanc, K.; Ringdén, O. Immunobiology of human mesenchymal stem cells and future use in hematopoietic stem cell transplantation. Biol. Blood Marrow Transpl. 2005, 11, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Rossini, A.; Frati, C.; Lagrasta, C.; Graiani, G.; Scopece, A.; Cavalli, S.; Musso, E.; Baccarin, M.; Di Segni, M.; Fagnoni, F.; et al. Human cardiac and bone marrow stromal cells exhibit distinctive properties related to their origin. Cardiovasc. Res. 2011, 89, 650–660. [Google Scholar] [CrossRef]
- Gambini, E.; Pompilio, G.; Biondi, A.; Alamanni, F.; Capogrossi, M.C.; Agrifoglio, M.; Pesce, M. C-kit+ cardiac progenitors exhibit mesenchymal markers and preferential cardiovascular commitment. Cardiovasc. Res. 2011, 89, 362–373. [Google Scholar] [CrossRef] [Green Version]
- Madonna, R.; Van Laake, L.W.; Davidson, S.M.; Engel, F.B.; Hausenloy, D.J.; Lecour, S.; Leor, J.; Perrino, C.; Schulz, R.; Ytrehus, K.; et al. Position Paper of the European Society of Cardiology Working Group Cellular Biology of the Heart: Cell-based therapies for myocardial repair and regeneration in ischemic heart disease and heart failure. Eur. Heart J. 2016, 37, 1789–1798. [Google Scholar] [CrossRef] [PubMed]
- Chimenti, I.; Smith, R.R.; Li, T.-S.; Gerstenblith, G.; Messina, E.; Giacomello, A.; Marbán, E. Relative Roles of Direct Regeneration Versus Paracrine Effects of Human Cardiosphere-Derived Cells Transplanted into Infarcted Mice. Circ. Res. 2010, 106, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Keith, M.C.L.; Bolli, R. “String Theory” of c-kit pos Cardiac Cells. Circ. Res. 2015, 116, 1216–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrella, F.; Spaggiari, L.; Acocella, F.; Barberis, M.; Bellomi, M.; Brizzola, S.; Donghi, S.; Giardina, G.; Giordano, R.; Guarize, J.; et al. Airway fistula closure after stem-cell infusion. N. Engl. J. Med. 2015, 372, 96–97. [Google Scholar] [CrossRef]
- La Francesca, S.; Aho, J.M.; Barron, M.R.; Blanco, E.W.; Soliman, S.; Kalenjian, L.; Hanson, A.D.; Todorova, E.; Marsh, M.; Burnette, K.; et al. Long-term regeneration and remodeling of the pig esophagus after circumferential resection using a retrievable synthetic scaffold carrying autologous cells. Sci. Rep. 2018, 8, 4123. [Google Scholar] [CrossRef]
- Caplan, H.; Olson, S.D.; Kumar, A.; George, M.; Prabhakara, K.S.; Wenzel, P.; Bedi, S.; Toledano-Furman, N.E.; Triolo, F.; Kamhieh-Milz, J.; et al. Mesenchymal Stromal Cell Therapeutic Delivery: Translational Challenges to Clinical Application. Front. Immunol. 2019, 10, 1645. [Google Scholar] [CrossRef]
- Gambini, E.; Pesce, M.; Persico, L.; Bassetti, B.; Gambini, A.; Alamanni, F.; Agrifoglio, M.; Capogrossi, M.C.; Pompilio, G. Patient profile modulates cardiac c-kit+ progenitor cell availability and amplification potential. Transl. Res. 2012, 160, 363–373. [Google Scholar] [CrossRef]
- Prigione, A.; Lichtner, B.; Kuhl, H.; Struys, E.A.; Wamelink, M.; Lehrach, H.; Ralser, M.; Timmermann, B.; Adjaye, J. Human iPSCs Harbor Homoplasmic and Heteroplasmic Mitochondrial DNA Mutations While Maintaining hESC-Like Metabolic Reprogramming. Stem Cells 2011, 29, 1338–1348. [Google Scholar] [CrossRef] [Green Version]
- Teslaa, T.; Teitell, M.A. Pluripotent stem cell energy metabolism: An update. EMBO J. 2015, 34, 138–153. [Google Scholar] [CrossRef]
- Schieke, S.M.; Ma, M.; Cao, L.; McCoy, J.P.; Liu, C.; Hensel, N.F.; Barrett, A.J.; Boehm, M.; Finkel, T. Mitochondrial Metabolism Modulates Differentiation and Teratoma Formation Capacity in Mouse Embryonic Stem Cells. J. Biol. Chem. 2008, 283, 28506–28512. [Google Scholar] [CrossRef] [Green Version]
- Avitabile, D.; Magenta, A.; Lauri, A.; Gambini, E.; Spaltro, G.; Vinci, M.C. Metaboloepigenetics: The Emerging Network in Stem Cell Homeostasis Regulation. Curr. Stem Cell Res. 2016, 11, 352–369. [Google Scholar] [CrossRef] [PubMed]
- Kubli, D.A.; Sussman, M.A. Eat, breathe, ROS: Controlling stem cell fate through metabolism. Expert Rev. Cardiovasc. 2017, 15, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietilä, M.; Palomäki, S.; Lehtonen, S.; Ritamo, I.; Valmu, L.; Nystedt, J.; Laitinen, S.; Leskelä, H.-V.; Sormunen, R.; Pesälä, J.; et al. Mitochondrial Function and Energy Metabolism in Umbilical Cord Blood- and Bone Marrow-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2012, 21, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, M.; Liu, J.; Mehta, G.U.; Patel, S.J.; Roychoudhuri, R.; Crompton, J.G.; Klebanoff, C.A.; Ji, Y.; Li, P.; Yu, Z.; et al. Mitochondrial Membrane Potential Identifies Cells with Enhanced Stemness for Cellular Therapy. Cell Metab. 2016, 23, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Sanada, F.; Kim, J.; Czarna, A.; Chan, N.Y.-K.; Signore, S.; Ogórek, B.; Isobe, K.; Wybieralska, E.; Borghetti, G.; Pesapane, A.; et al. c-Kit–Positive Cardiac Stem Cells Nested in Hypoxic Niches Are Activated by Stem Cell Factor Reversing the Aging Myopathy. Circ. Res. 2014, 114, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Vannini, N.; Girotra, M.; Naveiras, O.; Nikitin, G.; Campos, V.; Giger, S.; Roch, A.; Auwerx, J.; Lutolf, M.P. Specification of haematopoietic stem cell fate via modulation of mitochondrial activity. Nat. Commun. 2016, 7, 13125. [Google Scholar] [CrossRef]
- Rossi, J.M. Distinct mesodermal signals, including BMPs from the septum transversum mesenchyme, are required in combination for hepatogenesis from the endoderm. Genes Dev. 2001, 15, 1998–2009. [Google Scholar] [CrossRef] [Green Version]
- Birket, M.J.; Mummery, C.L. Pluripotent stem cell derived cardiovascular progenitors—A developmental perspective. Dev. Biol. 2015, 400, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Zhang, H.; Menzies, K.J.; Auwerx, J. The role of mitochondria in stem cell fate and aging. Development 2018, 145, dev143420. [Google Scholar] [CrossRef] [Green Version]
- Blazquez-Martinez, A.; Chiesa, M.; Arnalich, F.; Fernandez-Delgado, J.; Nistal, M.; De Miguel, M.P. c-Kit identifies a subpopulation of mesenchymal stem cells in adipose tissue with higher telomerase expression and differentiation potential. Differentiation 2014, 87, 147–160. [Google Scholar] [CrossRef] [PubMed]
- White, I.A.; Sanina, C.; Balkan, W.; Hare, J.M. Mesenchymal Stem Cells in Cardiology. Methods Mol. Biol. 2016, 1416, 55–87. [Google Scholar] [CrossRef] [Green Version]
- Aquila, I.; Cianflone, E.; Scalise, M.; Marino, F.; Mancuso, T.; Filardo, A.; Smith, A.J.; Cappetta, D.; De Angelis, A.; Urbanek, K.; et al. c-kit Haploinsufficiency impairs adult cardiac stem cell growth, myogenicity and myocardial regeneration. Cell Death Dis. 2019, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Vicinanza, C.; Aquila, I.; Cianflone, E.; Scalise, M.; Marino, F.; Mancuso, T.; Fumagalli, F.; Giovannone, E.D.; Cristiano, F.; Iaccino, E.; et al. Kitcre knock-in mice fail to fate-map cardiac stem cells. Nature 2018, 555, E1–E5. [Google Scholar] [CrossRef]
- Marino, F.; Scalise, M.; Cianflone, E.; Mancuso, T.; Aquila, I.; Agosti, V.; Torella, M.; Paolino, D.; Mollace, V.; Nadal-Ginard, B.; et al. Role of c-Kit in Myocardial Regeneration and Aging. Front. Endocrinol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.C.; Acsadi, G.; Brenner, C.A. Mitochondria: Determinants of Stem Cell Fate? Stem Cells Dev. 2009, 18, 803–806. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, T.; Brenner, C.; Bavister, B. Differentiation-related changes in mitochondrial properties as indicators of stem cell competence. J. Cell. Physiol. 2006, 208, 149–153. [Google Scholar] [CrossRef]
- Hill, B.G.; Benavides, G.A.; Lancaster, J.R.; Ballinger, S.; Dell’Italia, L.; Zhang, J.; Darley-Usmar, V.M. Integration of cellular bioenergetics with mitochondrial quality control and autophagy. Biol. Chem. 2012, 393, 1485–1512. [Google Scholar] [CrossRef] [Green Version]
- Mitra, K.; Lippincott-Schwartz, J. Analysis of Mitochondrial Dynamics and Functions Using Imaging Approaches. Curr. Protoc. Cell Biol. 2010, 46. [Google Scholar] [CrossRef] [Green Version]
- Mitra, K.; Wunder, C.; Roysam, B.; Lin, G.; Lippincott-Schwartz, J. A hyperfused mitochondrial state achieved at G1-S regulates cyclin E buildup and entry into S phase. Proc. Natl. Acad. Sci. USA 2009, 106, 11960–11965. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Detmer, S.A.; Ewald, A.J.; Griffin, E.E.; Fraser, S.E.; Chan, D.C. Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion and are essential for embryonic development. J. Cell Biol. 2003, 160, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Legros, F. Organization and dynamics of human mitochondrial DNA. J. Cell Sci. 2004, 117, 2653–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauvanet, C.; Duvezin-Caubet, S.; Salin, B.; David, C.; Massoni-Laporte, A.; di Rago, J.-P.; Rojo, M. Mitochondrial DNA Mutations Provoke Dominant Inhibition of Mitochondrial Inner Membrane Fusion. PLoS ONE 2012, 7, e49639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-T.; Shih, Y.-R.V.; Kuo, T.K.; Lee, O.K.; Wei, Y.-H. Coordinated Changes of Mitochondrial Biogenesis and Antioxidant Enzymes During Osteogenic Differentiation of Human Mesenchymal Stem Cells. Stem Cells 2008, 26, 960–968. [Google Scholar] [CrossRef]
- Chen, C.-T.; Hsu, S.-H.; Wei, Y.-H. Mitochondrial bioenergetic function and metabolic plasticity in stem cell differentiation and cellular reprogramming. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 571–576. [Google Scholar] [CrossRef]
- Zhang, Y.; Marsboom, G.; Toth, P.T.; Rehman, J. Mitochondrial Respiration Regulates Adipogenic Differentiation of Human Mesenchymal Stem Cells. PLoS ONE 2013, 8, e77077. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, A.; Yeung, K.Y.; Donoghue, J.; Baker, M.J.; Kelly, R.D.; McKenzie, M.; Johns, T.G.; St John, J.C. The regulation of mitochondrial DNA copy number in glioblastoma cells. Cell Death Differ. 2013, 20, 1644–1653. [Google Scholar] [CrossRef] [Green Version]
- Shock, L.S.; Thakkar, P.V.; Peterson, E.J.; Moran, R.G.; Taylor, S.M. DNA methyltransferase 1, cytosine methylation, and cytosine hydroxymethylation in mammalian mitochondria. Proc. Natl. Acad. Sci. USA 2011, 108, 3630–3635. [Google Scholar] [CrossRef] [Green Version]
- Demaria, M.; Giorgi, C.; Lebiedzinska, M.; Esposito, G.; D’Angeli, L.; Bartoli, A.; Gough, D.J.; Turkson, J.; Levy, D.E.; Watson, C.J.; et al. A STAT3-mediated metabolic switch is involved in tumour transformation and STAT3 addiction. Aging 2010, 2, 823–842. [Google Scholar] [CrossRef] [Green Version]
- Khacho, M.; Slack, R.S. Mitochondrial activity in the regulation of stem cell self-renewal and differentiation. Curr. Opin. Cell Biol. 2017, 49, 1–8. [Google Scholar] [CrossRef]
- Dai, D.-F.; Danoviz, M.E.; Wiczer, B.; Laflamme, M.A.; Tian, R. Mitochondrial Maturation in Human Pluripotent Stem Cell Derived Cardiomyocytes. Stem Cells Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Wolf, P.L.; Escudero, R.; Deutsch, R.; Jamieson, S.W.; Thistlethwaite, P.A. Early Expression of Angiogenesis Factors in Acute Myocardial Ischemia and Infarction. N. Engl. J. Med. 2000, 342, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Cencioni, C.; Atlante, S.; Savoia, M.; Martelli, F.; Farsetti, A.; Capogrossi, M.C.; Zeiher, A.M.; Gaetano, C.; Spallotta, F. The double life of cardiac mesenchymal cells: Epimetabolic sensors and therapeutic assets for heart regeneration. Pharmacol. Therapeut. 2017, 171, 43–55. [Google Scholar] [CrossRef]
- Gaspar, J.A.; Doss, M.X.; Hengstler, J.G.; Cadenas, C.; Hescheler, J.; Sachinidis, A. Unique Metabolic Features of Stem Cells, Cardiomyocytes, and Their Progenitors. Circ. Res. 2014, 114, 1346–1360. [Google Scholar] [CrossRef] [Green Version]
- Quiros, P.M.; Goyal, A.; Jha, P.; Auwerx, J. Analysis of mtDNA/nDNA Ratio in Mice. Curr. Protoc. Mouse Biol. 2017, 7, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Chopra, D.; Ray, L.; Dwivedi, A.; Tiwari, S.K.; Singh, J.; Singh, K.P.; Kushwaha, H.N.; Jahan, S.; Pandey, A.; Gupta, S.K.; et al. Photoprotective efficiency of PLGA-curcumin nanoparticles versus curcumin through the involvement of ERK/AKT pathway under ambient UV-R exposure in HaCaT cell line. Biomaterials 2016, 84, 25–41. [Google Scholar] [CrossRef]
- Pilato, C.A.; Stadiotti, I.; Maione, A.S.; Saverio, V.; Catto, V.; Tundo, F.; Dello Russo, A.; Tondo, C.; Pompilio, G.; Casella, M.; et al. Isolation and Characterization of Cardiac Mesenchymal Stromal Cells from Endomyocardial Bioptic Samples of Arrhythmogenic Cardiomyopathy Patients. J. Vis. Exp. 2018, 132, e57263. [Google Scholar] [CrossRef]
- McMahon, L.A.; Prendergast, P.J.; Campbell, V.A. A comparison of the involvement of p38, ERK1/2 and PI3K in growth factor-induced chondrogenic differentiation of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2008, 368, 990–995. [Google Scholar] [CrossRef]
- Gowran, A.; McKayed, K.; Kanichai, M.; White, C.; Hammadi, N.; Campbell, V. Tissue Engineering of Cartilage; Can Cannabinoids Help? Pharmaceuticals 2010, 3, 2970–2985. [Google Scholar] [CrossRef] [Green Version]
- Ullah, M.; Hamouda, H.; Stich, S.; Sittinger, M.; Ringe, J. A Reliable Protocol for the Isolation of Viable, Chondrogenically Differentiated Human Mesenchymal Stem Cells from High-Density Pellet Cultures. BioRes. Open Access 2012, 1, 297–305. [Google Scholar] [CrossRef] [Green Version]












© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambini, E.; Martinelli, I.; Stadiotti, I.; Vinci, M.C.; Scopece, A.; Eramo, L.; Sommariva, E.; Resta, J.; Benaouadi, S.; Cogliati, E.; et al. Differences in Mitochondrial Membrane Potential Identify Distinct Populations of Human Cardiac Mesenchymal Progenitor Cells. Int. J. Mol. Sci. 2020, 21, 7467. https://doi.org/10.3390/ijms21207467
Gambini E, Martinelli I, Stadiotti I, Vinci MC, Scopece A, Eramo L, Sommariva E, Resta J, Benaouadi S, Cogliati E, et al. Differences in Mitochondrial Membrane Potential Identify Distinct Populations of Human Cardiac Mesenchymal Progenitor Cells. International Journal of Molecular Sciences. 2020; 21(20):7467. https://doi.org/10.3390/ijms21207467
Chicago/Turabian StyleGambini, Elisa, Ilenia Martinelli, Ilaria Stadiotti, Maria Cristina Vinci, Alessandro Scopece, Luana Eramo, Elena Sommariva, Jessica Resta, Sabrina Benaouadi, Elisa Cogliati, and et al. 2020. "Differences in Mitochondrial Membrane Potential Identify Distinct Populations of Human Cardiac Mesenchymal Progenitor Cells" International Journal of Molecular Sciences 21, no. 20: 7467. https://doi.org/10.3390/ijms21207467
APA StyleGambini, E., Martinelli, I., Stadiotti, I., Vinci, M. C., Scopece, A., Eramo, L., Sommariva, E., Resta, J., Benaouadi, S., Cogliati, E., Paolin, A., Parini, A., Pompilio, G., & Savagner, F. (2020). Differences in Mitochondrial Membrane Potential Identify Distinct Populations of Human Cardiac Mesenchymal Progenitor Cells. International Journal of Molecular Sciences, 21(20), 7467. https://doi.org/10.3390/ijms21207467