Microbes as Medicines: Harnessing the Power of Bacteria in Advancing Cancer Treatment





Abstract
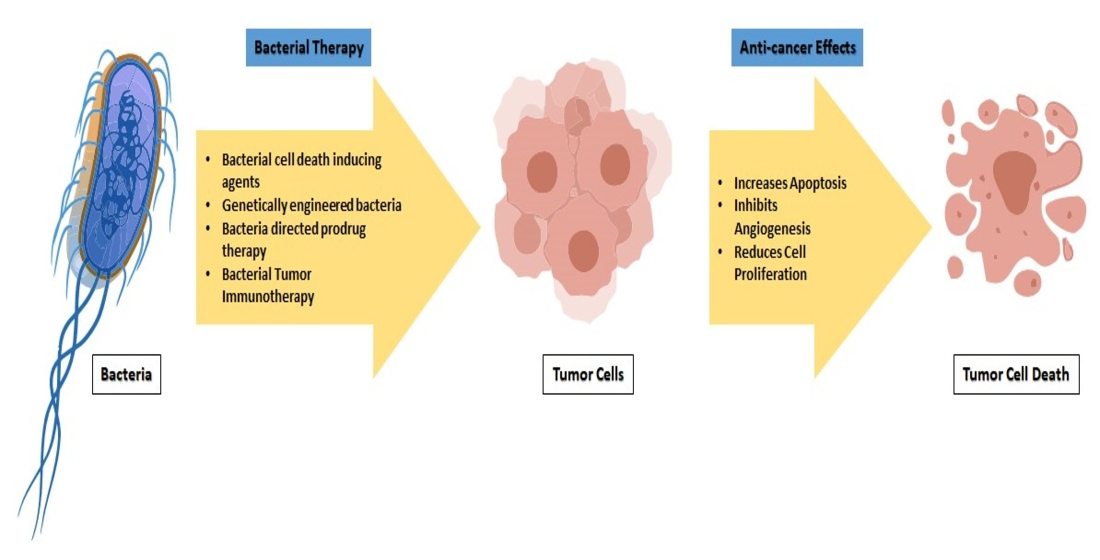
:
1. Introduction
2. Mechanism of Tumor Suppression
2.1. Bacterial Cell Death Inducing Agents
2.1.1. Bacterial Toxins
2.1.2. Bacterial Enzymes
2.1.3. Bacteriocins
2.1.4. Biosurfactants
2.2. Genetically Engineered Bacteria for Anti-Cancer Therapy
2.2.1. Virulence Attenuation
2.2.2. Tumor Targeting Enhancement
2.2.3. Bacterial Vectors for Anti-Cancer Cargo
2.2.4. Drug Expression Strategies
2.3. Bacterial Directed Enzyme Prodrug Therapy (BDEPT)
2.4. Anti-Angiogenic Therapy
2.5. Bacteria Tumor Immunotherapy
2.6. Bacteria Quorum Sensing for Tumor Targeting
2.7. Biofilm Based Anti-Cancer Therapy
3. Conventional Cancer Therapy in Combination with Bacteriotherapy
4. Challenges in Bacterial Cancer Therapy
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMP | Anti-microbial peptides |
| BDEPT | Bacterial directed enzyme prodrug therapy |
| CD | Cytosine Deaminase |
| CD8+ | Cytotoxic T-Lymphocytes |
| CHOP | Cyclophosphamide, doxorubicin, vincristine, and prednisone chemotherapy |
| ClyA | Cytolysin A |
| DT | Diphtheria Toxin |
| EDF | Extracellular death factor |
| EGFR | Epidermal growth factor receptor |
| FasL | Fas ligand |
| FNR | Fumarate and nitrate reduction regulator |
| HPRG | Histidine-proline-rich glycoprotein |
| HSV-TK | Herpes simplex virus thymidine kinase |
| IFN-γ | Interferon gamma |
| IPAF | Ice protease activating factor |
| L-ASNase | L-Asparaginase |
| LPS | Lipopolysaccharide |
| NK | Natural Killer cells |
| NLRP3 | Nucleotide binding domain, leucine rich containing, pyrin domain |
| O-DDHSL | N-3-oxo-dodecanoyl-L-homoserine lactone |
| QS | Quorum sensing |
| RGD | Arg-Gly-Asp |
| SLO | Streptolysin O |
| T3SS | Endostatin Type III secretion system |
| TME | Tumor microenvironment |
| TNF–α | Tumor necrosis factor–α |
| TLR | Toll like receptor |
| TRAIL | TNF–related apoptosis inducing ligands |
| T-regs | Regulatory T cells |
| VEGF | Vascular endothelial growth factor |
| 5-FC | 5-Flurocytosine |
| 5-FU | 5-Fluorouracil |
References
- Duong, M.T.Q.; Qin, Y.; You, S.H.; Min, J.J. Bacteria-cancer interactions: Bacteria-based cancer therapy. Exp. Mol. Med. 2019, 51, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Broadway, K.M.; Scharf, B.E. Salmonella Typhimurium as an Anticancer Therapy: Recent Advances and Perspectives. Curr. Clin. Microbiol. Rep. 2019, 6, 225–239. [Google Scholar] [CrossRef]
- Redelman-Sidi, G. Microbial Agents to Treat Cancer, 4th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; ISBN 9780128117378. [Google Scholar]
- Forbes, N.S.; Coffin, R.S.; Deng, L.; Evgin, L.; Fiering, S.; Giacalone, M.; Gravekamp, C.; Gulley, J.L.; Gunn, H.; Hoffman, R.M.; et al. White paper on microbial anti-cancer therapy and prevention. J. Immunother. Cancer 2018, 6, 1–24. [Google Scholar] [CrossRef]
- Nallar, S.C.; Xu, D.Q.; Kalvakolanu, D.V. Bacteria and genetically modified bacteria as cancer therapeutics: Current advances and challenges. Cytokine 2017, 89, 160–172. [Google Scholar] [CrossRef]
- Punj, V.; Saint-Dic, D.; Daghfal, S.; Kanwar, J.R. Microbial-based therapy of cancer: A new twist to age old practice. Cancer Biol. Ther. 2004, 3, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Staedtke, V.; Bai, R.Y.; Sun, W.; Huang, J.; Kibler, K.K.; Tyler, B.M.; Gallia, G.L.; Kinzler, K.; Vogelstein, B.; Zhou, S.; et al. Clostridium novyi-NT can cause regression of orthotopically implanted glioblastomas in rats. Oncotarget 2015, 6, 5536–5546. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Yu, B.; Cai, C.H.; Huang, J.D. Angiogenic inhibitors delivered by the type III secretion system of tumor-targeting Salmonella typhimurium safely shrink tumors in mice. AMB Express 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nemani, K.V.; Ennis, R.C.; Griswold, K.E.; Gimi, B. Magnetic nanoparticle hyperthermia induced cytosine deaminase expression in microencapsulated E coli for enzyme–Prodrug therapy. J. Biotechnol. 2015, 203, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yang, B.; Cheng, X.; Qiao, Y.; Tang, B.; Chen, G.; Wei, J.; Liu, X.; Cheng, W.; Du, P.; et al. Salmonella-mediated tumor-targeting TRAIL gene therapy significantly suppresses melanoma growth in mouse model. Cancer Sci. 2012, 103, 325–333. [Google Scholar] [CrossRef]
- Jiang, S.N.; Phan, T.X.; Nam, T.K.; Nguyen, V.H.; Kim, H.S.; Bom, H.S.; Choy, H.E.; Hong, Y.; Min, J.J. Inhibition of tumor growth and metastasis by a combination of escherichia coli-mediated cytolytic therapy and radiotherapy. Mol. Ther. 2010, 18, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Phan, T.X.; Nguyen, V.H.; van Dinh-Vu, H.; Zheng, J.H.; Yun, M.; Park, S.G.; Hong, Y.; Choy, H.E.; Szardenings, M.; et al. Salmonella typhimurium suppresses tumor growth via the pro-inflammatory cytokine interleukin-1β. Theranostics 2015, 5, 1328–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magaraci, M.S.; Veerakumar, A.; Qiao, P.; Amurthur, A.; Lee, J.Y.; Miller, J.S.; Goulian, M.; Sarkar, C.A. Engineering Escherichia coli for light-activated cytolysis of mammalian cells. ACS Synth. Biol. 2014, 3, 944–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardes, N.; Seruca, R.; Chakrabarty, A.M.; Fialho, A.M. Microbial-based therapy of Cancer Current progress and future prospects. Bioeng. Bugs 2010, 1, 178–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patyar, S.; Joshi, R.; Byrav, D.S.P.; Prakash, A.; Medhi, B.; Das, B.K. Bacteria in cancer therapy: A novel experimental strategy. J. Biomed. Sci. 2010, 17, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedighi, M.; Zahedi Bialvaei, A.; Hamblin, M.R.; Ohadi, E.; Asadi, A.; Halajzadeh, M.; Lohrasbi, V.; Mohammadzadeh, N.; Amiriani, T.; Krutova, M.; et al. Therapeutic bacteria to combat cancer; current advances, challenges, and opportunities. Cancer Med. 2019, 8, 3167–3181. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, S.; Piao, L.; Yuan, F. Radiotherapy combined with an engineered Salmonella typhimurium inhibits tumor growth in a mouse model of colon cancer. Exp. Anim. 2016, 65, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Proietti, S.; Nardicchi, V.; Porena, M.; Giannantoni, A. Botulinum toxin type-A toxin activity on prostate cancer cell lines. Urologia 2012, 79, 135–141. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Adamczak, A. Anticancer activity of bacterial proteins and peptides. Pharmaceutics 2018, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Bandala, C.; Perez-Santos, J.L.M.; Lara-Padilla, E.; Lopez, M.G.D.; Anaya-Ruiz, M. Effect of botulinum toxin A on proliferation and apoptosis in the T47D breast cancer cell line. Asian Pacific J. Cancer Prev. 2013, 14, 891–894. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.; Kim, K.S.; Kim, H.; Ko, K.C.; Song, J.J.; Choi, J.H.; Shin, M.; Min, J.J.; Jeong, J.H.; Choy, H.E. Anti-tumor activity of an immunotoxin (TGFα-PE38) delivered by attenuated Salmonella typhimurium. Oncotarget 2017, 8, 37550–37560. [Google Scholar] [CrossRef] [Green Version]
- Laliani, G.; Ghasemian Sorboni, S.; Lari, R.; Yaghoubi, A.; Soleimanpour, S.; Khazaei, M.; Hasanian, S.M.; Avan, A. Bacteria and cancer: Different sides of the same coin. Life Sci. 2020, 246, 117398. [Google Scholar] [CrossRef]
- Zam, W. Arginine enzymatic deprivation and diet restriction for cancer treatment. Brazilian J. Pharm. Sci. 2017, 53, 1–10. [Google Scholar] [CrossRef]
- Nguyen, H.A.; Su, Y.; Zhang, J.Y.; Antanasijevic, A.; Caffrey, M.; Schalk, A.M.; Liu, L.; Rondelli, D.; Oh, A.; Mahmud, D.L.; et al. A novel L-asparaginase with low L-glutaminase coactivity is highly efficacious against both T- and B-cell acute lymphoblastic Leukemias In Vivo. Cancer Res. 2018, 78, 1549–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemian, A.; Al-marzoqi, A.H.; Al-abodi, H.R.; Alghanimi, Y.K.; Kadhum, S.A.; Shokouhi Mostafavi, S.K.; Fattahi, A. Bacterial l-asparaginases for cancer therapy: Current knowledge and future perspectives. J. Cell. Physiol. 2019, 234, 19271–19279. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, A.; Czerniak, A.; Levy, T.; Amiur, S.; Gallula, J.; Matouk, I.; Abu-lail, R.; Sorin, V.; Birman, T.; de Groot, N.; et al. Development of targeted therapy for ovarian cancer mediated by a plasmid expressing diphtheria toxin under the control of H19 regulatory sequences. J. Transl. Med. 2009, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Scaiewicz, V.; Sorin, V.; Fellig, Y.; Birman, T.; Mizrahi, A.; Galula, J.; Abu-Lail, R.; Shneider, T.; Ohana, P.; Buscail, L.; et al. Use of H19 gene regulatory sequences in DNA-based therapy for pancreatic cancer. J. Oncol. 2010, 178174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasenpusch, G.; Pfeifer, C.; Aneja, M.K.; Wagner, K.; Reinhardt, D.; Gilon, M.; Ohana, P.; Hochberg, A.; Rudolph, C. Aerosolized bc-819 inhibits primary but not secondary lung cancer growth. PLoS ONE 2011, 6, e20760. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.S.; Park, S.O.; Yoon, A.R.; Yoo, J.Y.; Kim, M.K.; Yun, C.O.; Kim, C.W. Suicide cancer gene therapy using pore-forming toxin, streptolysin O. Mol. Cancer Ther. 2006, 5, 1610–1619. [Google Scholar] [CrossRef] [Green Version]
- Fotoohi-Ardakani, G.; Kheirollahi, M.; Zarei Jaliani, H.; Noorian, M.; Ansariniyia, H. Targeting MCF-7 Cell Line by Listeriolysin O Pore Forming Toxin Fusion with AHNP Targeted Peptide. Adv. Biomed. Res. 2019, 8, 33. [Google Scholar]
- Ishii, K.J.; Kawakami, K.; Gursel, I.; Conover, J.; Joshi, B.H.; Klinman, D.M.; Puri, R.K. Antitumor Therapy with Bacterial DNA and Toxin: Complete Regression of Established Tumor Induced by Liposomal CpG Oligodeoxynucleotides plus Interleukin-13 Cytotoxin. Clin. Cancer Res. 2003, 9, 6516–6522. [Google Scholar]
- Fiedler, T.; Strauss, M.; Hering, S.; Redanz, U.; William, D.; Rosche, Y.; Classen, C.F.; Kreikemeyer, B.; Linnebacher, M.; Maletzki, C. Arginine deprivation by arginine deiminase of Streptococcus pyogenes controls primary glioblastoma growth in vitro and in vivo. Cancer Biol. Ther. 2015, 16, 1047–1055. [Google Scholar] [CrossRef] [Green Version]
- Do, T.T.; Do, T.P.; Nguyen, T.N.; Nguyen, T.C.; Vu, T.T.P.; Nguyen, T.G.A.; Quang Khieu, D. Nanoliposomal L-Asparaginase and Its Antitumor Activities in Lewis Lung Carcinoma Tumor-Induced BALB/c Mice. Adv. Mater. Sci. Eng. 2019, 2019, 3534807. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, S.; Ghollasi, M.; Hosseini, H.M. The apoptotic impact of nisin as a potent bacteriocin on the colon cancer cells. Microb. Pathog. 2017, 111, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Arunmanee, W.; Ecoy, G.A.U.; Khine, H.E.E.; Duangkaew, M.; Prompetchara, E.; Chanvorachote, P.; Chaotham, C. Colicin N mediates apoptosis and suppresses integrin-modulated survival in human lung cancer cells. Molecules 2020, 25, 816. [Google Scholar] [CrossRef] [Green Version]
- Paiva, A.D.; de Oliveira, M.D.; de Paula, S.O.; Baracat-Pereira, M.C.; Breukink, E.; Mantovani, H.C. Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiology 2012, 158, 2851–2858. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, J.H.; Lee, Y.J.; Lee, S.J.; Kim, Y. Surfactin suppresses TPA-induced breast cancer cell invasion through the inhibition of MMP-9 expression. Int. J. Oncol. 2013, 42, 287–296. [Google Scholar] [CrossRef]
- Kunda, N.K. Antimicrobial peptides as novel therapeutics for non-small cell lung cancer. Drug Discov. Today 2020, 25, 238–247. [Google Scholar] [CrossRef]
- Baindara, P.; Korpole, S.; Grover, V. Bacteriocins: Perspective for the development of novel anticancer drugs. Appl. Microbiol. Biotechnol. 2018, 102, 10393–10408. [Google Scholar] [CrossRef]
- Kaur, S.; Kaur, S. Bacteriocins as potential anticancer agents. Front. Pharmacol. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Baindara, P.; Gautam, A.; Raghava, G.P.S.; Korpole, S. Anticancer properties of a defensin like class IId bacteriocin Laterosporulin10. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactants produced by marine microorganisms with therapeutic applications. Mar. Drugs 2016, 14, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, C.; Gudiña, E.J.; Lima, C.F.; Rodrigues, L.R. Effects of biosurfactants on the viability and proliferation of human breast cancer cells. AMB Express 2014, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.S.; Ngai, S.C.; Goh, B.H.; Chan, K.G.; Lee, L.H.; Chuah, L.H. Anticancer activities of surfactin potential application of nanotechnology assisted surfactin delivery. Front. Pharmacol. 2017, 8, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Abdelli, F.; Jardak, M.; Elloumi, J.; Stien, D.; Cherif, S.; Mnif, S.; Aifa, S. Antibacterial, anti-adherent and cytotoxic activities of surfactin(s) from a lipolytic strain Bacillus safensis F4. Biodegradation 2019, 30, 287–300. [Google Scholar] [CrossRef]
- Karlapudi, A.P.; Venkateswarulu, T.C.; Srirama, K.; Kota, R.K.; Mikkili, I.; Kodali, V.P. Evaluation of anti-cancer, anti-microbial and anti-biofilm potential of biosurfactant extracted from an Acinetobacter M6 strain. J. King Saud Univ. Sci. 2020, 32, 223–227. [Google Scholar] [CrossRef]
- Baban, C.K.; Cronin, M.; O’Hanlon, D.; O’Sullivan, G.C.; Tangney, M. Bacteria as vectors for gene therapy of cancer. Bioeng. Bugs 2010, 1, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Gravekamp, C.; Bermudes, D.; Liu, K. Tumour-targeting bacteria engineered to fight cancer. Nat. Rev. Cancer 2018, 18, 727–743. [Google Scholar] [CrossRef]
- Jia, L.J.; Wei, D.P.; Sun, Q.M.; Huang, Y.; Wu, Q.; Hua, Z.C. Oral delivery of tumor-targeting Salmonella for cancer therapy in murine tumor models. Cancer Sci. 2007, 98, 1107–1112. [Google Scholar] [CrossRef]
- Toso, J.F.; Gill, V.J.; Hwu, P.; Marincola, F.M.; Restifo, N.P.; Schwartzentruber, D.J.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Stock, F.; et al. Phase I Study of the Intravenous Administration of Attenuated Salmonella typhimurium to Patients with Metastatic Melanoma. J. Clin. Oncol. 2002, 20, 142–152. [Google Scholar] [CrossRef]
- Kong, Q.; Six, D.A.; Liu, Q.; Gu, L.; Roland, K.L.; Raetz, C.R.H.; Curtiss, R. Palmitoylation state impacts induction of innate and acquired immunity by the Salmonella enterica serovar Typhimurium msbB mutant. Infect. Immun. 2011, 79, 5027–5038. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Park, S.H.; Tan, W.; Hong, S.H.; Shin, M.G.; Chung, I.J.; Hong, Y.; Bom, H.S.; et al. Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci. Transl. Med. 2017, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Geller, J.; Ma, H.; Yang, M.; Penman, S.; Hoffman, R.M. Monotherapy with a tumor-targeting mutant of Salmonella typhimurium cures orthotopic metastatic mouse models of human prostate cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 10170–10174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroshima, Y.; Zhang, Y.; Murakami, T.; Maawy, A.; Miwa, S.; Yamamoto, M.; Yano, S.; Sato, S.; Momiyama, M.; Mori, R.; et al. Efficacy of tumor-targeting Salmonella typhimurium A1-R in combination with anti-angiogenesis therapy on a pancreatic cancer patient-derived orthotopic xenograft (PDOX) and cellline mouse models. Oncotarget 2014, 5, 12346–12357. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Miwa, S.; Zhang, Y.; Zhao, M.; Yano, S.; Uehara, F.; Yamamoto, M.; Hiroshima, Y.; Toneri, M.; Bouvet, M.; et al. Intraperitoneal administration of tumor-targeting Salmonella typhimurium A1-R inhibits disseminated human ovarian cancer and extends survival in nude mice. Oncotarget 2015, 6, 11369–11377. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.; Husseiny, M.I.; Song, L.; Erdreich-Epstein, A.; Shackleford, G.M.; Seeger, R.C.; Jäckel, D.; Hensel, M.; Metelitsa, L.S. Novel cancer vaccine based on genes of Salmonella pathogenicity island 2. Int. J. Cancer 2010, 126, 2622–2634. [Google Scholar]
- Park, S.H.; Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Kim, D.Y.; Szardenings, M.; Min, J.H.; Hong, Y.; Choy, H.E.; Min, J.J. RGD peptide cell-surface display enhances the targeting and therapeutic efficacy of attenuated salmonella-mediated cancer therapy. Theranostics 2016, 6, 1672–1682. [Google Scholar] [CrossRef] [Green Version]
- Massa, P.E.; Paniccia, A.; Monegal, A.; De Marco, A.; Rescigno, M. Salmonella engineered to express CD20-targeting antibodies and a drug-converting enzyme can eradicate human lymphomas. Blood 2013, 122, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Yoon, W.S.; Chae, Y.S.; Hong, J.; Park, Y.K. Antitumor therapeutic effects of a genetically engineered Salmonella typhimurium harboring TNF-α in mice. Appl. Microbiol. Biotechnol. 2011, 89, 1807–1819. [Google Scholar] [CrossRef]
- Ganai, S.; Arenas, R.B.; Forbes, N.S. Tumour-targeted delivery of TRAIL using Salmonella typhimurium enhances breast cancer survival in mice. Br. J. Cancer 2009, 101, 1683–1691. [Google Scholar] [CrossRef]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Inhibition of tumor growth using Salmonella expressing fas ligand. J. Natl. Cancer Inst. 2008, 100, 1113–1116. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, S.; Esmaeili, A.; Barzegari, A.; Rafi, M.A.; Omidi, Y. Bioengineered smart bacterial carriers for combinational targeted therapy of solid tumours. J. Drug Target. 2020, 28, 700–713. [Google Scholar] [CrossRef]
- Rooseboom, M.; Commandeur, J.N.M.; Vermeulen, N.P.E. Enzyme-Catalyzed Activation of Anticancer Prodrugs. Pharmacol. Rev. 2004, 56, 53–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guise, C.P.; Abbattista, M.R.; Tipparaju, S.R.; Lambie, N.K.; Su, J.; Li, D.; Wilson, W.R.; Dachs, G.U.; Patterson, A.V. Diflavin oxidoreductases activate the bioreductive prodrug PR-104A under hypoxia. Mol. Pharmacol. 2012, 81, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Endostatin gene therapy delivered by Salmonella choleraesuis in murine tumor models. J. Gene Med. 2004, 6, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Niethammer, A.G.; Xiang, R.; Becker, J.C.; Wodrich, H.; Pertl, U.; Karsten, G.; Eliceir, B.P.; Reisfeld, R.A. A DNA vaccine against VEGF receptor 2 prevents effective angiogenesis and inhibits tumor growth. Nat. Med. 2002, 8, 1369–1375. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, H.; Mizutani, M.; Mizutani, N.; Reisfeld, R.A.; Xiang, R. Transcription factor Fos-related antigen 1 is an effective target for a breast cancer vaccine. Proc. Natl. Acad. Sci. USA 2003, 100, 8850–8855. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, S.A. IL-2: The First Effective Immunotherapy for Human Cancer. J. Immunol. 2014, 192, 5451–5458. [Google Scholar] [CrossRef]
- Shahabi, V.; Reyes-Reyes, M.; Wallecha, A.; Rivera, S.; Paterson, Y.; MacIag, P. Development of a Listeria monocytogenes based vaccine against prostate cancer. Cancer Immunol. Immunother. 2008, 57, 1301–1313. [Google Scholar] [CrossRef]
- Roussel, L.; Rousseau, S. Exposure of airway epithelial cells to Pseudomonas aeruginosa biofilm-derived quorum sensing molecules decrease the activity of the anti-oxidant response element bound by NRF2. Biochem. Biophys. Res. Commun. 2017, 483, 829–833. [Google Scholar] [CrossRef]
- Miyake, K.; Yamamoto, S.; Iijima, S. Blocking adhesion of cancer cells to endothelial cell types by S. agalactiae type-specific polysaccharides. In Proceedings of the Cytotechnology; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; Volume 22, pp. 205–209. [Google Scholar]
- Van Dessel, N.; Swofford, C.A.; Forbes, N.S. Potent and tumor specific: Arming bacteria with therapeutic proteins. Ther. Deliv. 2015, 6, 385–399. [Google Scholar] [CrossRef] [Green Version]
- Ryan, R.M.; Green, J.; Williams, P.J.; Tazzyman, S.; Hunt, S.; Harmey, J.H.; Kehoe, S.C.; Lewis, C.E. Bacterial delivery of a novel cytolysin to hypoxic areas of solid tumors. Gene Ther. 2009, 16, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Min, S.Y.; Lim, H.D.; You, S.H.; Lim, D.; Jeong, J.H.; Kim, H.J.; Rhee, J.H.; Park, K.; Shin, M.; et al. Cell mass-dependent expression of an anticancer protein drug by tumor-targeted Salmonella. Oncotarget 2018, 9, 8548–8559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, N.S. Engineering the perfect (bacterial) cancer therapy. Nat. Rev. Cancer 2010, 10, 785–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehouritis, P.; Springer, C.; Tangney, M. Bacterial-directed enzyme prodrug therapy. J. Control. Release 2013, 170, 120–131. [Google Scholar] [CrossRef]
- Zhang, J.; Kale, V.; Chen, M. Gene-Directed Enzyme Prodrug Therapy. AAPS 2015, 17, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Lehouritis, P.; Hogan, G.; Tangney, M. Designer bacteria as intratumoural enzyme biofactories. Adv. Drug Deliv. Rev. 2017, 118, 8–23. [Google Scholar] [CrossRef]
- Chakrabarty, A.M.; Fialho, A.M. Microbial Infections and Cancer Therapy; Pan Stanford Publishing Pte. Ltd.: Singapore, 2019; ISBN 978-1-351-04190-4. [Google Scholar]
- Popczun, E.J.; Read, C.G.; Roske, C.W.; Lewis, N.S.; Schaak, R.E. Phase I/II Study of APS001F With Flucytosine and Maltose in Solid Tumors. Angew. Chemie Int. Ed. 2014, 53, 5427–5430. [Google Scholar] [CrossRef]
- Afkhami-poostchi, A.; Mashreghi, M.; Iranshahi, M.; Matin, M.M. Use of a genetically engineered E. coli overexpressing β -glucuronidase accompanied by glycyrrhizic acid, a natural and anti-in fl ammatory agent, for directed treatment of colon carcinoma in a mouse model. Int. J. Pharm. 2020, 579, 119159. [Google Scholar] [CrossRef]
- Ho, C.L.; Tan, H.Q.; Chua, K.J.; Kang, A.; Lim, K.H.; Ling, K.L.; Yew, W.S.; Lee, Y.S.; Thiery, J.P.; Chang, M.W. Engineered commensal microbes for diet-mediated colorectal-cancer chemoprevention. Nat. Biomed. Eng. 2018, 2, 27–37. [Google Scholar] [CrossRef]
- Chan-hyams, J.V.E.; Copp, J.N.; Smaill, B.; Patterson, A.V. Evaluating the abilities of diverse nitroaromatic prodrug metabolites to exit a model Gram negative vector for bacterial-directed enzyme-prodrug therapy. Biochem. Pharmacol. 2018, 158, 192–200. [Google Scholar] [CrossRef]
- Lehouritis, P.; Stanton, M.; Mccarthy, F.O.; Jeavons, M.; Tangney, M. Activation of multiple chemotherapeutic prodrugs by the natural enzymolome of tumour-localised probiotic bacteria. J. Control. Release 2016, 222, 9–17. [Google Scholar] [CrossRef]
- Cummins, J.; Tangney, M. Bacteria and tumours: Causative agents or opportunistic inhabitants? Infect. Agent. Cancer 2013, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Gardlik, R.; Behuliak, M.; Palffy, R.; Celec, P.; Li, C.J. Gene therapy for cancer: Bacteria-mediated anti-angiogenesis therapy. Gene Ther. 2011, 18, 425–431. [Google Scholar] [CrossRef]
- Song, S.; Vuai, M.S.; Zhong, M. The role of bacteria in cancer therapy—Enemies in the past, but allies at present. Infect. Agent. Cancer 2018, 13, 1–7. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, X.; Cheng, W.; Chen, J.; Ma, C.; Yang, B.; Hua, Z.-C. Tumor-Specific Delivery of Histidine-Rich Glycoprotein Suppresses Tumor Growth and Metastasis by Anti-angiogenesis and Vessel Normalization. Curr. Gene Ther. 2014, 14, 75–85. [Google Scholar] [CrossRef]
- Chen, J.; Qiao, Y.; Tang, B.; Chen, G.; Liu, X.; Yang, B.; Wei, J.; Zhang, X.; Cheng, X.; Du, P.; et al. Modulation of Salmonella tumor-colonization and intratumoral anti-angiogenesis by triptolide and its mechanism. Theranostics 2017, 7, 2250–2260. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Kim, Y.; Lee, H.B.; Kwon, H.J. Antiangiogenic activity of the lipophilic antimicrobial peptides from an endophytic bacterial strain isolated from red pepper leaf. Mol. Cells 2015, 38, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Ji, G.; Li, Q.; Yang, Y.; Shui, L.; Shen, Y.; Yang, H. Bacterial particles retard tumor growth as a novel vascular disrupting agent. Biomed. Pharmacother. 2020, 122, 109757. [Google Scholar] [CrossRef]
- Talib, W.H.; Saleh, S. Propionibacterium acnes Augments Antitumor, Anti-Angiogenesis and Immunomodulatory Effects of Melatonin on Breast Cancer Implanted in Mice. PLoS ONE 2015, 10, e0124384. [Google Scholar] [CrossRef]
- Torres, W.; Lameda, V.; Olivar, L.C.; Navarro, C.; Fuenmayor, J.; Pérez, A.; Mindiola, A.; Rojas, M.; Martínez, M.S.; Velasco, M.; et al. Bacteria in cancer therapy: Beyond immunostimulation. J. Cancer Metastasis Treat. 2018, 4, 4. [Google Scholar] [CrossRef]
- Kucerova, P.; Cervinkova, M. Spontaneous regression of tumour and the role of microbial infection—Possibilities for cancer treatment. Anticancer. Drugs 2016, 27, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, W.; Chang, Y.; Kim, J.; Seok, Y.; Hye, J.; Min, S.; Park, S.; Yoo, Y.; Keun, Y.; Mo, B. ScienceDirect Application of genetically engineered Salmonella typhimurium for interferon-gamma e induced therapy against melanoma. Eur. J. Cancer 2017, 70, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Lansley, S.M.; Varano Della Vergiliana, J.F.; Cleaver, A.L.; Ren, S.H.; Segal, A.; Xu, M.Y.; Lee, Y.C.G. A commercially available preparation of Staphylococcus aureus bio-products potently inhibits tumour growth in a murine model of mesothelioma. Respirology 2014, 19, 1025–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, J.; Tang, B.; Zhang, X.; Hua, Z.-C. Systemic administration of attenuated Salmonella typhimurium in combination with interleukin-21 for cancer therapy. Mol. Clin. Oncol. 2013, 1, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.Y.; Park, H.T.; Dinh, N.T.H.; Choi, S.J.; Lee, J.; Kim, J.H.; Lee, S.W.; Gho, Y.S. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Declue, A.E.; Axiak-Bechtel, S.M.; Zhang, Y.; Saha, S.; Zhang, L.; Tung, D.; Bryan, J.N. Immune response to C. novyi-NT immunotherapy. Vet. Res. 2018, 49, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daille, R.; Waldschmitt, N.; Boneca, I.G.; Chamaillard, M.; Waldschmitt, N.; Yamazaki, T.; Isnard, C.; Poirier-colame, V.; Duong, C.P.M.; Flament, C.; et al. Enterococcus hirae and Barnesiella intestinihominis Facilitate Cyclophosphamide-Induced Therapeutic Immunomodulatory Effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [Green Version]
- Wynendaele, E.; Pauwels, E.; Van de Wiele, C.; Burvenich, C.; De Spiegeleer, B. The potential role of quorum-sensing peptides in oncology. Med. Hypotheses 2012, 78, 814–817. [Google Scholar] [CrossRef]
- Wynendaele, E.; Verbeke, F.; D’Hondt, M.; Hendrix, A.; Van De Wiele, C.; Burvenich, C.; Peremans, K.; De Wever, O.; Bracke, M.; De Spiegeleer, B. Crosstalk between the microbiome and cancer cells by quorum sensing peptides. Peptides 2015, 64, 40–48. [Google Scholar] [CrossRef] [Green Version]
- De Spiegeleer, B.; Verbeke, F.; D’Hondt, M.; Hendrix, A.; Van DeWiele, C.; Burvenich, C.; Peremans, K.; DeWever, O.; Bracke, M.; Wynendaele, E. The quorum sensing peptides PhrG, CSP and EDF promote angiogenesis and invasion of breast cancer cells in vitro. PLoS ONE 2015, 10, e0119471. [Google Scholar] [CrossRef]
- Moghaddam, M.M. Quorum Sensing in Bacteria and a Glance on Pseudomonas aeruginosa. Clin. Microbiol. Open Access 2014, 3, 1–10. [Google Scholar] [CrossRef]
- Anderson, J.C.; Clarke, E.J.; Arkin, A.P.; Voigt, C.A.; Berkeley, E.O.L. Environmentally Controlled Invasion of Cancer Cells by Engineered Bacteria. J. Mol. Biol. 2006, 355, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Castro, S.; Coker, C.; Hinchliffe, T.E.; Arpaia, N.; Danino, T. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat. Med. 2019, 25, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Dougan, M.; Dougan, S.K. Programmable bacteria as cancer therapy. Nat. Med. 2019, 25, 1030–1031. [Google Scholar] [CrossRef] [PubMed]
- Ran, D.; Mao, J.; Zhan, C.; Xie, C.; Ruan, H.; Ying, M.; Zhou, J.; Lu, W.; Lu, W. D-Retroenantiomer of Quorum-Sensing Peptide-Modi fi ed Polymeric Micelles for Brain Tumor-Targeted Drug Delivery. ACS Appl. Mater. Interfaces 2017, 9, 25672–25682. [Google Scholar] [CrossRef]
- Kumar, A.S.; Bryan, J.N.; Kumar, S.R. Bacterial Quorum Sensing Molecule N-3-Oxo- Dodecanoyl-L-Homoserine Lactone Causes Direct Cytotoxicity and Reduced Cell Motility in Human Pancreatic Carcinoma Cells. PLoS ONE 2014, 9, e106480. [Google Scholar] [CrossRef]
- Balhouse, B.N.; Patterson, L.; Schmelz, E.M.; Slade, D.J.; Verbridge, S.S. N-(3-oxododecanoyl)-L-homoserine lactone interactions in the breast tumor microenvironment: Implications for breast cancer viability and proliferation in vitro. PLoS ONE 2017, 12, e0180372. [Google Scholar] [CrossRef] [Green Version]
- Nandakumar, N.; Dandela, R.; Gopas, J.; Meijler, M.M. Quorum sensing modulators exhibit cytotoxicity in Hodgkin’s lymphoma cells and interfere with NF-κB signaling. Bioorganic Med. Chem. Lett. 2017, 27, 2967–2973. [Google Scholar] [CrossRef]
- Rizzato, C.; Torres, J.; Kasamatsu, E.; Camorlinga-Ponce, M.; Bravo, M.M.; Canzian, F.; Kato, I. Potential role of biofilm formation in the development of digestive tract cancer with special reference to helicobacter pylori infection. Front. Microbiol. 2019, 10, 1–21. [Google Scholar] [CrossRef]
- Raskov, H.; Kragh, K.N.; Bjarnsholt, T.; Alamili, M.; Gögenur, I. Bacterial biofilm formation inside colonic crypts may accelerate colorectal carcinogenesis. Clin. Transl. Med. 2018, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Groizeleau, J.; Rybtke, M.; Andersen, J.B.; Berthelsen, J.; Liu, Y.; Yang, L.; Nielsen, T.E.; Kaever, V.; Givskov, M.; Tolker-nielsen, T.; et al. The anti-cancerous drug doxorubicin decreases the c-di-GMP content in Pseudomonas aeruginosa but promotes biofilm formation. Microbiology 2016, 162, 1797–1807. [Google Scholar] [CrossRef] [PubMed]
- Podlesek, Z.; Žgur Bertok, D. The DNA Damage Inducible SOS Response Is a Key Player in the Generation of Bacterial Persister Cells and Population Wide Tolerance. Front. Microbiol. 2020, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Weitao, T. Bacteria form biofilms against cancer metastasis Why we should treat idiopathic optic neuritis with high-dose and long-term corticosteroids in China. Med. Hypotheses 2009, 72, 477–478. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Khan, S.; Al-Shammari, E.; Patel, M.; Saeed, M.; Hadi, S. In pursuit of cancer metastasis therapy by bacteria and its biofilms: History or future. Med. Hypotheses 2017, 100, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Kumeria, T.; Maher, S.; Wang, Y.; Kaur, G.; Wang, L.; Erkelens, M.; Forward, P.; Lambert, M.F.; Evdokiou, A.; Losic, D. Naturally Derived Iron Oxide Nanowires from Bacteria for Magnetically Triggered Drug Release and Cancer Hyperthermia in 2D and 3D Culture Environments: Bacteria Biofilm to Potent Cancer Therapeutic. Biomacromolecules 2016, 17, 2726–2736. [Google Scholar] [CrossRef]
- Wang, Z.H.; Liu, J.M.; Li, C.Y.; Wang, D.; Lv, H.; Lv, S.W.; Zhao, N.; Ma, H.; Wang, S. Bacterial Biofilm Bioinspired Persistent Luminescence Nanoparticles with Gut-Oriented Drug Delivery for Colorectal Cancer Imaging and Chemotherapy. ACS Appl. Mater. Interfaces 2019, 11, 36409–36419. [Google Scholar] [CrossRef]
- Padma, V.V. An overview of targeted cancer therapy. BioMedicine 2015, 5, 19. [Google Scholar] [CrossRef]
- Damyanov, C.A.; Maslev, I.K.; Pavlov, V.S. Conventional Treatment of Cancer Realities and Problems. Ann. Complement. Altern. Med. 2018, 1, 1–9. [Google Scholar]
- Kizaka-Kondoh, S.; Inoue, M.; Harada, H.; Hiraoka, M. Tumor hypoxia: A target for selective cancer therapy. Cancer Sci. 2003, 94, 1021–1028. [Google Scholar] [CrossRef]
- Jia, L.J.; Wei, D.P.; Sun, Q.M.; Jin, G.H.; Li, S.F.; Huang, Y.; Hua, Z.C. Tumor-targeting Salmonella typhimurium improves cyclophosphamide chemotherapy at maximum tolerated dose and low-dose metronomic regimens in a murine melanoma model. Int. J. Cancer 2007, 121, 666–674. [Google Scholar] [CrossRef]
- Bascuas, T.; Moreno, M.; Grille, S.; Chabalgoity, J.A. Salmonella immunotherapy improves the outcome of CHOP chemotherapy in non-Hodgkin lymphoma-bearing mice. Front. Immunol. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, L.H.; Bettegowda, C.; Agrawal, N.; Cheong, I.; Huso, D.; Frost, P.; Loganzo, F.; Greenberger, L.; Barkoczy, J.; Pettit, G.R.; et al. Targeting Vascular and Avascular Compartments of Tumors with C. novyi-NT and Anti-microtubule Agents. Cancer Biol. Ther. 2004, 3, 326–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltzman, D.; Augustin, L.; Leonard, A.; Mertensotto, M.; Schottel, J. Low dose chemotherapy combined with attenuated Salmonella decreases tumor burden and is less toxic than high dose chemotherapy in an autochthonous murine model of breast cancer. Surgery 2018, 163, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, M.; Du, L.; Zeng, J.; Yao, T.; Jin, Y. Paclitaxel-in-liposome-in-bacteria for inhalation treatment of primary lung cancer. Int. J. Pharm. 2020, 578, 119177. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Xie, Y.; Xu, S.; Xin, J.; Wang, J.; Han, T.; Ting, R.; Zhang, J.; An, F. Hypoxia-activated nanomedicines for effective cancer therapy. Eur. J. Med. Chem. 2020, 195, 112274. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, Y.; Qin, M.; Zhang, X.; Zhang, Z.; Sun, X.; Gu, Z. Bacteria-Driven Hypoxia Targeting for Combined Biotherapy and Photothermal Therapy. ACS Nano 2018, 12, 5995–6005. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.M.; El-Gebaly, R.H.; Hamad, A.M. Combination of bacteriolytic therapy with magnetic field for Ehrlich tumour treatment. Gen. Physiol. Biophys. 2017, 36, 259–271. [Google Scholar] [CrossRef]
- Park, W.; Cho, S.; Huang, X.; Larson, A.C.; Kim, D.-H. Branched Gold Nanoparticle Coating of Clostridium novyi-NT Spores for CT-Guided Intratumoral Injection. Small 2017, 13, 1602722. [Google Scholar] [CrossRef] [Green Version]
- Park, W.; Cho, S.; Kang, D.; Han, J.H.; Park, J.H.; Lee, B.; Lee, J.; Kim, D.H. Tumor Microenvironment Targeting Nano–Bio Emulsion for Synergistic Combinational X-Ray PDT with Oncolytic Bacteria Therapy. Adv. Healthc. Mater. 2020, 9, 1–8. [Google Scholar] [CrossRef]
- Teng, G.; Ju, Y.; Yang, Y.; Hua, H.U.; Chi, J.; Mu, X. Combined antitumor activity of the nitroreductase/CB1954 suicide gene system and γ–rays in HeLa cells in vitro. Mol. Med. Rep. 2016, 14, 5164–5170. [Google Scholar] [CrossRef] [Green Version]
- Curran, C.S.; Rasooly, A.; He, M.; Prickril, B.; Thurin, M.; Sharon, E. Report on the NCI microbial-based cancer therapy conference. Cancer Immunol. Res. 2018, 6, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, M.G.; Masner, M.; Ferreira, F.A.; Hoffman, R.M. Bacterial therapy of cancer: Promises, limitations, and insights for future directions. Front. Microbiol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treatment of Patients with Cancer with Genetically Modified Salmonella Typhimurium Bacteria—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT00004988 (accessed on 12 October 2020).
- VNP20009 in Treating Patients with Advanced Solid Tumors—Full Text View—ClinicalTrials.gov. Available online: https://www.clinicaltrials.gov/ct2/show/NCT00006254 (accessed on 12 October 2020).
- IL-2 Expressing, Attenuated Salmonella Typhimurium in Unresectable Hepatic Spread—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01099631 (accessed on 12 October 2020).
- VXM01 Phase I Dose Escalation Study in Patients with Locally Advanced, Inoperable and Stage IV Pancreatic Cancer—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01486329 (accessed on 12 October 2020).
- Safety & Immunogenicity of JNJ-64041809, a Live Attenuated Double-deleted Listeria Immunotherapy, in Participants with Metastatic Castration-resistant Prostate Cancer—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02625857 (accessed on 12 October 2020).
- Le, D.T.; Wang-Gillam, A.; Picozzi, V.; Greten, T.F.; Crocenzi, T.; Springett, G.; Morse, M.; Zeh, H.; Cohen, D.; Fine, R.L.; et al. Safety and survival with GVAX pancreas prime and Listeria monocytogenes-expressing mesothelin (CRS-207) boost vaccines for metastatic pancreatic cancer. J. Clin. Oncol. 2015, 33, 1325–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safety and Efficacy of Combination Listeria/GVAX Pancreas Vaccine in the Pancreatic Cancer Setting—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02004262 (accessed on 12 October 2020).
- Basu, P.; Mehta, A.; Jain, M.; Gupta, S.; Nagarkar, R.V.; John, S.; Petit, R. A Randomized Phase 2 Study of ADXS11-001 Listeria monocytogenes-Listeriolysin O Immunotherapy With or Without Cisplatin in Treatment of Advanced Cervical Cancer. Int. J. Gynecol. Cancer 2018, 28, 764–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Study of ADXS11-001 in Subjects with High Risk Locally Advanced Cervical Cancer—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02853604 (accessed on 12 October 2020).
- Safety Study of Intratumoral Injection of Clostridium Novyi-NT Spores to Treat Patients With Solid Tumors That Have Not Responded to Standard Therapies—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01924689 (accessed on 12 October 2020).
- Pembrolizumab with Intratumoral Injection of Clostridium Novyi-NT—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03435952 (accessed on 12 October 2020).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |



{kind=link}
{kind=link}
{kind=link}
| Bacterial Metabolites—Category | Bacterial Strain | Metabolite | Mechanism of Action | Biological Target: Cancer Cells/Cell Lines | References |
|---|---|---|---|---|---|
| Bacterial Toxins | Corynebacterium diphtheria | Diphtheria toxin (DT) 2 major fragments (DTA and DTB) | DTA is responsible for the cytotoxic enzymatic activity and inactivates the ADP-ribosylation of elongation factor 2; DTB facilitates cell entry by binding to surface receptors and subsequent translocation into cytoplasm by undergoing endocytosis | Ovarian cancer (SKOV-3), Pancreatic cancer (CRL-1687, CRL-2119, CRL-1997, and CRL-2547), Lung cancer (NCI-H460, NCI-H358, and A549) | [26,27,28] |
| Streptococcus | Streptolysin O (SLO) | SLO binds to cholesterol in the plasma membrane, oligomerizes to form aggregates which form large pores. This results in cytolysis and cell death | Embryonic kidney fibroblasts (293T, HEK 293 cell derivatives that harbor SV40 large T antigen) | [29] | |
| Listeria monocytogenes | Listeriolysin O | Binds to cholesterol binding receptors and induces pore formation in the cell membrane resulting in cytolysis. Induces apoptosis in T-cells by caspase mediated pathway | Breast cancer cells (MDA-MB-231 and MCF-7) | [30] | |
| Pseudomonas aeruginosa | Exotoxin A | Targeting with tumor-related antigens and induction of cytotoxic pathways | Head and neck cancer cell line (KCCT873) | [31] | |
| Bacterial Enzymes | Mycoplasma hominis or M. arginine | Arginine deiminase | Hydrolyzes arginine and deprives the tumor of arginine, essential for growth. This results in reduced tumor proliferation | Glioblastoma (HROG02, HROG05, HROG10) | [32] |
| Escherichia coli | L-Asparaginase | Catalyzes asparagine hydrolysis and reduces its blood concentration. This results in selective growth inhibition of malignant cells | Breast carcinoma (MCF-7), Hepatocellular carcinoma (HepG2), Lung carcinoma (SK-LU-1) | [33] | |
| Bacteriocins | Lactococcus lactis | Nisin A | Induces cell cycle arrest and apoptosis through activation of CHAC1 | Colon cancer (SW480) cells | [34] |
| Escherichia coli | Colicin | Binds to a specific receptor on the outer membrane and forms pores which leads to apoptosis | Lung cancer (H460, H292, and H23) cells | [35] | |
| Staphylococcus bovis HC5 | Bovicin | Binds to the cell membrane and disrupts cell membrane integrity by pore formation. It also induces the potassium efflux in target cells | Breast cancer cells (MCF7), Liver cancer cells (HepG2) | [36] | |
| Biosurfactants | Bacillus subtilis | Surfactin, a cyclic lipopeptide | Inhibits tumor cell invasion, migration, and colony formation | Breast carcinoma cells (MCF-7 and MDA-MB-231) | [37] |
| Treatment Strategy | Bacterial Strain | Gene/Drug | Mechanism of Action | Application | References |
|---|---|---|---|---|---|
| Prodrug Therapy | Salmonella | Thymidine kinase polypeptide Prodrug: Ganciclovir | Inhibits deoxyguanosine triphosphate, dGTP, incorporation into DNA | Melanoma | [63] |
| Prodrug Therapy Anti-Angiogenic Therapy | Escherichia Coli | Uridine phosphorylase Prodrug: Capecitabine | Impede thymidylate synthase enzyme | Colon, rectum, and head and neck cancers | [64] |
| S. choleraesuis | Endostatin | Increases infiltration of CD8(+) T cells | Melanoma, Bladder tumor, Hepatoma | [65] | |
| Anti-Angiogenic Therapy Anti-Angiogenic Therapy Immunotherapy | S. typhimurium SL7207 | VEGFR-2 | Upregulates vascular-endothelial growth factor receptor 2 (FLK-1) of proliferating endothelial cells in the tumor vasculature | Melanoma, Colon carcinoma, Lung carcinoma, | [66] |
| S. typhimurium RE88 | IL-18 | Activation of T, natural killer, and dendritic cells | Breast carcinoma | [67] | |
| Anti-Angiogenic Therapy Immunotherapy Quorum Sensing peptides for anti-tumor action | Serratia marcescens | Endotoxin | Releases pro-inflammatory cytokines, making the immune system eliminate or protect against multiple tumors | Melanoma, Leukemia, Lymphoma | [68] |
| Listeria monocytogenes | Listeriolysin O | Releases proinflammatory cytokines and increases expression of co-stimulant molecules in antigen presenting cells surfaces leading to maturation and activation of high affinity T cells | Prostate cancer | [69] | |
| Pseudomonas. aeruginosa | N-3-oxododecanoyl homoserine lactone (3OC12-HSL) | Inhibition by protein kinase | Cystic fibrosis | [70] | |
| Biofilms as Anti-Cancer Agents | Streptococcus agalactiae | Polysaccharides | Inhibit adhesion of cancer cells to endothelial cells | Colon cancer | [71] |
| Bacterial Strain | Gene/Strain | Tumor Model | Phase | Observation | Identifier (NCT Number) | Reference |
|---|---|---|---|---|---|---|
| Salmonella typhimurium | VNP20009 with HSV-TK | B16F10 melanomas | I | Dose-dependent suppression of tumor growth and prolonged survival | NCT00004988 | [136] |
| VNP20009 | Metastatic melanoma, metastatic renal cell carcinoma | I | Induced a dose-related increase in the circulation of proinflammatory cytokines, such as IL-1β, TNF-α, IL-6, and IL-12 | NCT00006254 | [137] | |
| χ4550 with IL-2 | Hepatoma, liver neoplasms | I | Consistent reduction in the mean number of hepatic metastases in fed animals | NCT01099631 | [138] | |
| Salmonella typhimurium | VXM01 | Pancreatic cancer | I | Reduction in tumor perfusion after vaccination | NCT01486329 | [139] |
| Listeria monocytogenes | JNJ-64041809 | CT26 Colon tumor, Prostate cancer | I | Breaking of self-tolerance and long-term survival | NCT02625857 | [140] |
| GVAX+ CRS-207, Drug: Cyclophosphamide | Metastatic pancreatic cancer | II | Extended survival for patients with pancreatic cancer, with minimal toxicity | NCT01417000 NCT02004262 | [141,142] | |
| ADXS11-001 | Cervical Cancer | II, III | Promising safety and efficacy results | NCT02853604 | [143,144] | |
| Clostridium | novyi-NT | Solid tumor malignancies | I | Reduced the tumor size | NCT01924689 | [145] |
| novyi-NT Drug: Pembrolizumab | Refractory advanced solid tumors | I | Ongoing | NCT03435952 | [146] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawant, S.S.; Patil, S.M.; Gupta, V.; Kunda, N.K. Microbes as Medicines: Harnessing the Power of Bacteria in Advancing Cancer Treatment. Int. J. Mol. Sci. 2020, 21, 7575. https://doi.org/10.3390/ijms21207575
Sawant SS, Patil SM, Gupta V, Kunda NK. Microbes as Medicines: Harnessing the Power of Bacteria in Advancing Cancer Treatment. International Journal of Molecular Sciences. 2020; 21(20):7575. https://doi.org/10.3390/ijms21207575
Chicago/Turabian StyleSawant, Shruti S., Suyash M. Patil, Vivek Gupta, and Nitesh K. Kunda. 2020. "Microbes as Medicines: Harnessing the Power of Bacteria in Advancing Cancer Treatment" International Journal of Molecular Sciences 21, no. 20: 7575. https://doi.org/10.3390/ijms21207575
APA StyleSawant, S. S., Patil, S. M., Gupta, V., & Kunda, N. K. (2020). Microbes as Medicines: Harnessing the Power of Bacteria in Advancing Cancer Treatment. International Journal of Molecular Sciences, 21(20), 7575. https://doi.org/10.3390/ijms21207575