Uncharacterized RNAs in Plasma of Alzheimer’s Patients Are Associated with Cognitive Impairment and Show a Potential Diagnostic Power

 ,
,  ,
, 
 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Transcriptome Analysis of Plasma Samples of AD Patients Compared to Unaffected Individuals
2.2. Validation of Microarray Results by Real-Time PCR
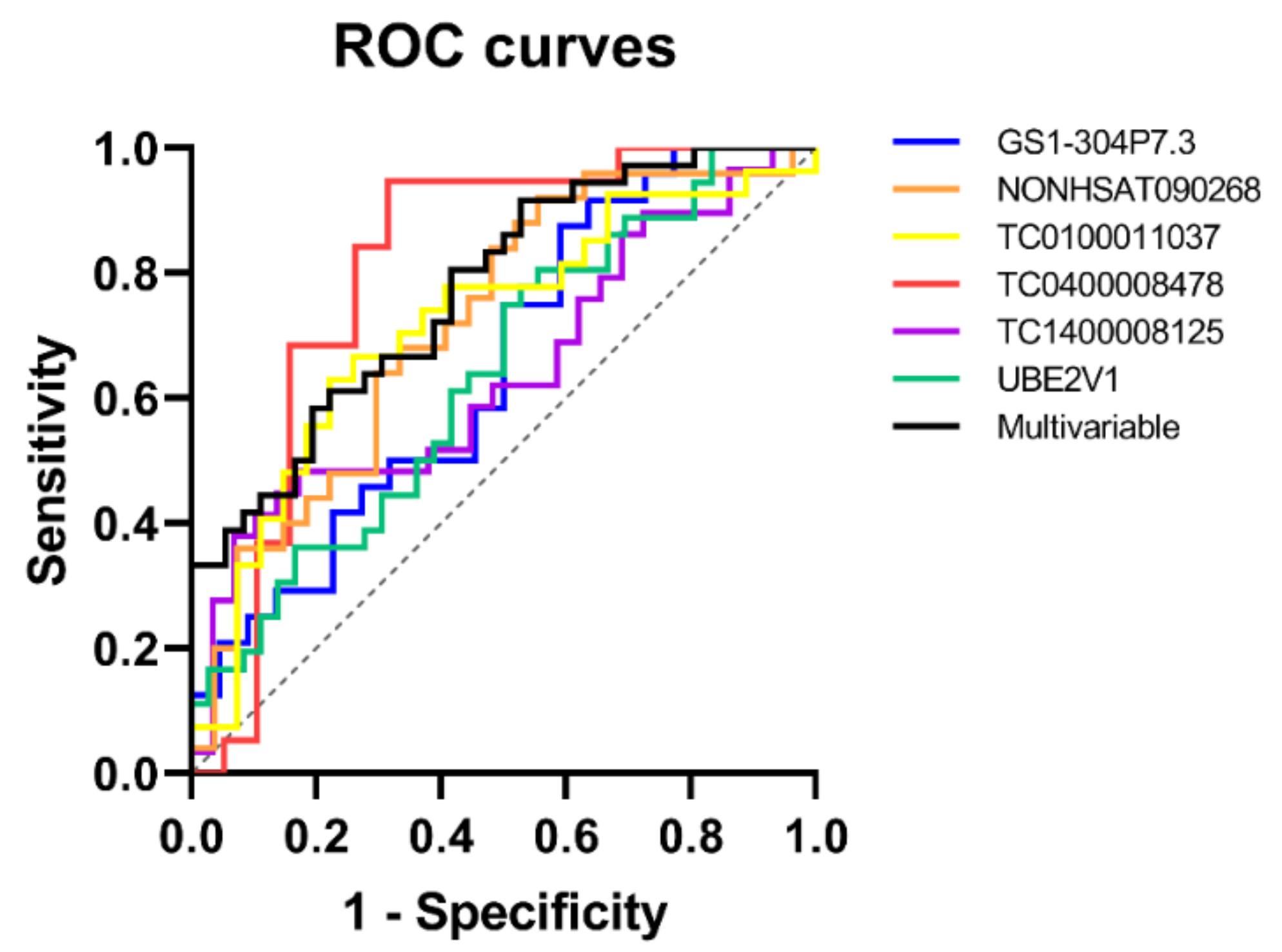
2.3. Evaluation of Diagnostic Accuracy through ROC Curves
2.4. Peripheral Blood-Isolated Cell Expression Analysis
2.5. Correlation with Clinical and Cognitive Phenotypes
3. Discussion
4. Materials and Methods
4.1. Patient Recruitment and Plasma Sample Processing
4.2. RNA Isolation from Plasma Samples
4.3. Microarray Analysis
4.4. Validation of Microarray Results Using Real-Time PCR
4.5. ROC Curve Analysis
4.6. Expression Analysis in Peripheral Blood Mononuclear Cells
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| ADAMTS9 | ADAM metallopeptidase with thrombospondin type 1 motif 9 |
| ADAMTS9-AS2 | ADAMTS9 antisense RNA 2 |
| ARSJ | arylsulfatase family member J |
| AUC | area under the curve |
| Aβ | amyloid beta |
| circRNAs | circular RNAs |
| CIs | confidence intervals |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| CTRL | Controls |
| DE | differentially expressed |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| lncRNAs | long non-coding RNAs |
| miRNAs | microRNAs |
| MMSE | Mini Mental State Examination |
| ncRNAs | non-coding RNAs |
| NPV | negative predictive value |
| PBMCs | peripheral blood mononuclear cells |
| PDC | Phosducin |
| PDC-AS1 | PDC antisense RNA 1 |
| PPV | positive predictive value |
| PSD95 | postsynaptic density-95 |
| PTGS2 | prostaglandin-endoperoxide synthase 2 |
| RNU6 | RNA, U6 small nuclear 1 |
| ROC | receiver operating characteristic |
| RPL23A | ribosomal protein L23a |
| UBE2V1 | ubiquitin conjugating enzyme E2 V1) |
| UGT8 | UDP glycosyltransferase 8 |
References
- Khoury, R.; Grossberg, G.T. Deciphering Alzheimer’s disease: Predicting new therapeutic strategies via improved understanding of biology and pathogenesis. Expert Opin. Ther. Targets 2020, 24, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Castellano, S.; Salomone, S.; Drago, F.; Bosco, P.; Nuovo, S. Searching for Disease-Modifying Drugs in AD: Can We Combine Neuropsychological Tools with Biological Markers? CNS Neurol. Disord. Drug Targets 2014, 13, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Copani, A.; Nicoletti, F.; Drago, F. Depression and Alzheimer’s disease: Neurobiological links and common pharmacological targets. Eur. J. Pharmacol. 2010, 626, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Sharma, N. Exploring Biomarkers for Alzheimer’s Disease. J. Clin. Diagn. Res. 2016, 10, KE01–KE06. [Google Scholar] [CrossRef]
- Hampel, H.; Vergallo, A.; Caraci, F.; Cuello, A.C.; Lemercier, P.; Vellas, B.; Giudici, K.V.; Baldacci, F.; Hänisch, B.; Haberkamp, M.; et al. Future avenues for Alzheimer’s disease detection and therapy: Liquid biopsy, intracellular signaling modulation, systems pharmacology drug discovery. Neuropharmacology 2020, 108081. [Google Scholar] [CrossRef]
- Zetterberg, H.; Burnham, S.C. Blood-based molecular biomarkers for Alzheimer’s disease. Mol. Brain 2019, 12, 26. [Google Scholar] [CrossRef]
- Barbagallo, C.; Mostile, G.; Baglieri, G.; Giunta, F.; Luca, A.; Raciti, L.; Zappia, M.; Purrello, M.; Ragusa, M.; Nicoletti, A. Specific Signatures of Serum miRNAs as Potential Biomarkers to Discriminate Clinically Similar Neurodegenerative and Vascular-Related Diseases. Cell. Mol. Neurobiol. 2019, 40, 531–546. [Google Scholar] [CrossRef]
- Sheinerman, K.S.; Toledo, J.B.; Tsivinsky, V.G.; Irwin, D.; Grossman, M.; Weintraub, D.; Hurtig, H.I.; Chen-Plotkin, A.; Wolk, D.A.; McCluskey, L.F.; et al. Circulating brain-enriched microRNAs as novel biomarkers for detection and differentiation of neurodegenerative diseases. Alzheimer’s Res. Ther. 2017, 9, 89. [Google Scholar] [CrossRef]
- Cirnigliaro, M.; Barbagallo, C.; Gulisano, M.; Domini, C.N.; Barone, R.; Barbagallo, D.; Ragusa, M.; Di Pietro, C.; Rizzo, R.; Purrello, M. Expression and Regulatory Network Analysis of miR-140-3p, a New Potential Serum Biomarker for Autism Spectrum Disorder. Front. Mol. Neurosci. 2017, 10, 250. [Google Scholar] [CrossRef] [Green Version]
- Di Pietro, V.; Porto, E.; Ragusa, M.; Barbagallo, C.; Davies, D.; Forcione, M.; Logan, A.; Di Pietro, C.; Purrello, M.; Grey, M.; et al. Salivary MicroRNAs: Diagnostic Markers of Mild Traumatic Brain Injury in Contact-Sport. Front. Mol. Neurosci. 2018, 11, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipów, S.; Łaczmański, Ł. Blood Circulating miRNAs as Cancer Biomarkers for Diagnosis and Surgical Treatment Response. Front. Genet. 2019, 10, 169. [Google Scholar] [CrossRef] [PubMed]
- Drokow, E.K.; Sun, K.; Ahmed, H.A.W.; Akpabla, G.S.; Song, J.; Shi, M. Circulating microRNA as diagnostic biomarkers for haematological cancers: A systematic review and meta-analysis. Cancer Manag. Res. 2019, 11, 4313–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Mauro, S.; Ragusa, M.; Urbano, F.; Filippello, A.; Di Pino, A.; Scamporrino, A.; Pulvirenti, A.; Ferro, A.; Rabuazzo, A.M.; Purrello, M.; et al. Intracellular and extracellular miRNome deregulation in cellular models of NAFLD or NASH: Clinical implications. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 1129–1139. [Google Scholar] [CrossRef]
- Párrizas, M.; Novials, A. Circulating microRNAs as biomarkers for metabolic disease. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 591–601. [Google Scholar] [CrossRef]
- Battaglia, R.; Palini, S.; Vento, M.E.; La Ferlita, A.; Lo Faro, M.J.; Caroppo, E.; Borzì, P.; Falzone, L.; Barbagallo, D.; Ragusa, M.; et al. Identification of extracellular vesicles and characterization of miRNA expression profiles in human blastocoel fluid. Sci. Rep. 2019, 9, 84. [Google Scholar] [CrossRef]
- Alsaweed, M.; Hartmann, P.; Geddes, D.; Kakulas, F. MicroRNAs in Breastmilk and the Lactating Breast: Potential Immunoprotectors and Developmental Regulators for the Infant and the Mother. Int. J. Environ. Res. Public Health 2015, 12, 13981–14020. [Google Scholar] [CrossRef] [Green Version]
- Layne, T.R.; Green, R.A.; Lewis, C.A.; Nogales, F.; Cruz, T.C.D.; Zehner, Z.E.; Seashols-Williams, S.J. MicroRNA Detection in Blood, Urine, Semen, and Saliva Stains After Compromising Treatments. J. Forensic Sci. 2019, 64, 1831–1837. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Song, J. The Role of Long Noncoding RNAs in Diabetic Alzheimer’s Disease. J. Clin. Med. 2018, 7, 461. [Google Scholar] [CrossRef] [Green Version]
- Hampel, H.; Caraci, F.; Cuello, A.C.; Caruso, G.; Nisticò, R.; Corbo, M.; Baldacci, F.; Toschi, N.; Garaci, F.; Chiesa, P.A.; et al. A Path Toward Precision Medicine for Neuroinflammatory Mechanisms in Alzheimer’s Disease. Front. Immunol. 2020, 11, 456. [Google Scholar] [CrossRef]
- Itzhaki, R.F.; Lathe, R.; Balin, B.J.; Ball, M.J.; Bearer, E.L.; Braak, H.; Bullido, M.J.; Carter, C.; Clerici, M.; Cosby, S.L.; et al. Microbes and Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 51, 979–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialopiotrowicz, E.; Kuzniewska, B.; Kachamakova-Trojanowska, N.; Barcikowska, M.; Kuznicki, J.; Wojda, U. Cell cycle regulation distinguishes lymphocytes from sporadic and familial Alzheimer’s disease patients. Neurobiol. Aging 2011, 32, 2319.e13–2319.e26. [Google Scholar] [CrossRef]
- Rodrigues, R.; Debom, G.; Soares, F.; Machado, C.; Pureza, J.; Peres, W.; Garcias, G.d.L.; Duarte, M.F.; Schetinger, M.R.C.; Stefanello, F.; et al. Alterations of ectonucleotidases and acetylcholinesterase activities in lymphocytes of Down syndrome subjects: Relation with inflammatory parameters. Clin. Chim. Acta 2014, 433, 105–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuello, A.C. Early and Late CNS Inflammation in Alzheimer’s Disease: Two Extremes of a Continuum? Trends Pharmacol. Sci. 2017, 38, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Gulick, J.; Osinska, H.; Yu, Y.; McLendon, P.M.; Shay-Winkler, K.; Robbins, J.; Yutzey, K.E. Ube2v1 Positively Regulates Protein Aggregation by Modulating Ubiquitin Proteasome System Performance Partially Through K63 Ubiquitination. Circ. Res. 2020, 126, 907–922. [Google Scholar] [CrossRef]
- Sanfeliu, A.; Hokamp, K.; Gill, M.; Tropea, D. Transcriptomic Analysis of Mecp2 Mutant Mice Reveals Differentially Expressed Genes and Altered Mechanisms in Both Blood and Brain. Front. Psychiatry 2019, 10, 278. [Google Scholar] [CrossRef]
- Ma, Q.; Ruan, H.; Peng, L.; Zhang, M.; Gack, M.U.; Yao, W.-D. Proteasome-independent polyubiquitin linkage regulates synapse scaffolding, efficacy, and plasticity. Proc. Natl. Acad. Sci. USA 2017, 114, E8760–E8769. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Zhang, R.; Sun, X. Enhancer LncRNAs Influence Chromatin Interactions in Different Ways. Front. Genet. 2019, 10, 936. [Google Scholar] [CrossRef]
- Romero-Barrios, N.; Legascue, M.F.; Benhamed, M.; Ariel, F.; Crespi, M. Splicing regulation by long noncoding RNAs. Nucleic Acids Res. 2018, 46, 2169–2184. [Google Scholar] [CrossRef] [Green Version]
- Müller, R.; Weirick, T.; John, D.; Militello, G.; Chen, W.; Dimmeler, S.; Uchida, S. ANGIOGENES: Knowledge database for protein-coding and noncoding RNA genes in endothelial cells. Sci. Rep. 2016, 6, 32475. [Google Scholar] [CrossRef] [Green Version]
- Vagnucci, A.H.; Li, W.W. Alzheimer’s disease and angiogenesis. Lancet 2003, 361, 605–608. [Google Scholar] [CrossRef]
- Castro, M.G.; Biron, K.E.; Dickstein, D.L.; Gopaul, R.; Jefferies, W.A. Amyloid Triggers Extensive Cerebral Angiogenesis Causing Blood Brain Barrier Permeability and Hypervascularity in Alzheimer’s Disease. PLoS ONE 2011, 6, e23789. [Google Scholar] [CrossRef] [Green Version]
- Ragusa, M.; Barbagallo, C.; Brex, D.; Caponnetto, A.; Cirnigliaro, M.; Battaglia, R.; Barbagallo, D.; Di Pietro, C.; Purrello, M. Molecular Crosstalking among Noncoding RNAs: A New Network Layer of Genome Regulation in Cancer. Int. J. Genom. 2017, 2017, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Broeckel, U.; Stoll, M.; Hein, L. The identification of phosducin as a novel candidate gene for hypertension and its role in sympathetic activation. Curr. Opin. Nephrol. Hypertens. 2011, 20, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Beetz, N.; Harrison, M.D.; Brede, M.; Zong, X.; Urbanski, M.J.; Sietmann, A.; Kaufling, J.; Barrot, M.; Seeliger, M.W.; Vieira-Coelho, M.A.; et al. Phosducin influences sympathetic activity and prevents stress-induced hypertension in humans and mice. J. Clin. Investig. 2009, 119, 3597–3612. [Google Scholar] [CrossRef]
- Skoog, I.; Gustafson, D. Update on hypertension and Alzheimer’s disease. Neurol. Res. 2013, 28, 605–611. [Google Scholar] [CrossRef]
- García-Alberca, J.M.; Mendoza, S.; Gris, E.; Royo, J.L.; Cruz-Gamero, J.M.; García-Casares, N. White matter lesions and temporal atrophy are associated with cognitive and neuropsychiatric symptoms in patients with hypertension and Alzheimer’s disease. Int. J. Geriatr. Psychiatry 2020. [Google Scholar] [CrossRef]
- Chen, Q.; Liang, B.; Wang, Z.; Cheng, X.; Huang, Y.; Liu, Y.; Huang, Z. Influence of four polymorphisms in ABCA1 and PTGS2 genes on risk of Alzheimer’s disease: A meta-analysis. Neurol. Sci. 2016, 37, 1209–1220. [Google Scholar] [CrossRef]
- Ma, S.L.; Tang, N.L.S.; Zhang, Y.P.; Ji, L.-d.; Tam, C.W.C.; Lui, V.W.C.; Chiu, H.F.K.; Lam, L.C.W. Association of prostaglandin-endoperoxide synthase 2 (PTGS2) polymorphisms and Alzheimer’s disease in Chinese. Neurobiol. Aging 2008, 29, 856–860. [Google Scholar] [CrossRef]
- Wang, P.; Guan, P.-P.; Wang, T.; Yu, X.; Guo, J.-J.; Wang, Z.-Y. Aggravation of Alzheimer’s disease due to the COX-2-mediated reciprocal regulation of IL-1β and Aβ between glial and neuron cells. Aging Cell 2014, 13, 605–615. [Google Scholar] [CrossRef]
- Woodling, N.S.; Colas, D.; Wang, Q.; Minhas, P.; Panchal, M.; Liang, X.; Mhatre, S.D.; Brown, H.; Ko, N.; Zagol-Ikapitte, I.; et al. Cyclooxygenase inhibition targets neurons to prevent early behavioural decline in Alzheimer’s disease model mice. Brain 2016, 139, 2063–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Esparcia, P.; Sideris-Lampretsas, G.; Hernandez-Ortega, K.; Grau-Rivera, O.; Sklaviadis, T.; Gelpi, E.; Ferrer, I. Altered mechanisms of protein synthesis in frontal cortex in Alzheimer disease and a mouse model. Am. J. Neurodegener. Dis. 2017, 6, 15–25. [Google Scholar] [PubMed]
- Evans, H.T.; Benetatos, J.; van Roijen, M.; Bodea, L.G.; Götz, J. Decreased synthesis of ribosomal proteins in tauopathy revealed by non-canonical amino acid labelling. EMBO J. 2019, 38, e101174. [Google Scholar] [CrossRef] [PubMed]
- Cooper-Knock, J.; Shaw, P.J.; Moll, T. Disrupted glycosylation of lipids and proteins is a cause of neurodegeneration. Brain 2020, 143, 1332–1340. [Google Scholar] [CrossRef]
- McKenzie, A.T.; Moyon, S.; Wang, M.; Katsyv, I.; Song, W.-M.; Zhou, X.; Dammer, E.B.; Duong, D.M.; Aaker, J.; Zhao, Y.; et al. Multiscale network modeling of oligodendrocytes reveals molecular components of myelin dysregulation in Alzheimer’s disease. Mol. Neurodegener. 2017, 12, 82. [Google Scholar] [CrossRef]
- Katsel, P.; Li, C.; Haroutunian, V. Gene Expression Alterations in the Sphingolipid Metabolism Pathways during Progression of Dementia and Alzheimer’s Disease: A Shift Toward Ceramide Accumulation at the Earliest Recognizable Stages of Alzheimer’s Disease? Neurochem. Res. 2007, 32, 845–856. [Google Scholar] [CrossRef]
- Yao, J.; Zhou, B.; Zhang, J.; Geng, P.; Liu, K.; Zhu, Y.; Zhu, W. A new tumor suppressor LncRNA ADAMTS9-AS2 is regulated by DNMT1 and inhibits migration of glioma cells. Tumor Biol. 2014, 35, 7935–7944. [Google Scholar] [CrossRef]
- Kamboh, M.I.; Demirci, F.Y.; Wang, X.; Minster, R.L.; Carrasquillo, M.M.; Pankratz, V.S.; Younkin, S.G.; Saykin, A.J.; Jun, G.; Baldwin, C.; et al. Genome-wide association study of Alzheimer’s disease. Transl. Psychiatry 2012, 2, e117. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.-G.; Lin, E.; Tsai, S.-J.; Kuo, P.-H.; Liu, Y.-L.; Yang, A.C.; Kao, C.-F.; Yang, C.-H. The ADAMTS9 gene is associated with cognitive aging in the elderly in a Taiwanese population. PLoS ONE 2017, 12, e0172440. [Google Scholar] [CrossRef]
- Li, N.; Li, J.; Mi, Q.; Xie, Y.; Li, P.; Wang, L.; Binang, H.; Wang, Q.; Wang, Y.; Chen, Y.; et al. Long non-coding RNA ADAMTS9-AS1 suppresses colorectal cancer by inhibiting the Wnt/β-catenin signalling pathway and is a potential diagnostic biomarker. J. Cell. Mol. Med. 2020. [Google Scholar] [CrossRef]
- Caricasole, A. Induction of Dickkopf-1, a Negative Modulator of the Wnt Pathway, Is Associated with Neuronal Degeneration in Alzheimer’s Brain. J. Neurosci. 2004, 24, 6021–6027. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.; Tapia-Rojas, C. Loss of canonical Wnt signaling is involved in the pathogenesis of Alzheimer’s disease. Neural Regen. Res. 2018, 13, 1705. [Google Scholar] [CrossRef] [PubMed]
- Khoury, R.; Ghossoub, E. Diagnostic biomarkers of Alzheimer’s disease: A state-of-the-art review. Biomark. Neuropsychiatry 2019, 1, 100005. [Google Scholar] [CrossRef]
- Risacher, S.L.; Fandos, N.; Romero, J.; Sherriff, I.; Pesini, P.; Saykin, A.J.; Apostolova, L.G.; Zetterberg, H. Plasma amyloid beta levels are associated with cerebral amyloid and tau deposition. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2019, 11, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Kaneko, N.; Villemagne, V.L.; Kato, T.; Doecke, J.; Doré, V.; Fowler, C.; Li, Q.-X.; Martins, R.; Rowe, C.; et al. High performance plasma amyloid-β biomarkers for Alzheimer’s disease. Nature 2018, 554, 249–254. [Google Scholar] [CrossRef]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group* under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef] [Green Version]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. Mini-mental state. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Doody, R.S.; Massman, P.; Dunn, J.K. A Method for Estimating Progression Rates in Alzheimer Disease. Arch. Neurol. 2001, 58, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Di Mauro, S.; Scamporrino, A.; Petta, S.; Urbano, F.; Filippello, A.; Ragusa, M.; Di Martino, M.T.; Scionti, F.; Grimaudo, S.; Pipitone, R.M.; et al. Serum coding and non-coding RNAs as biomarkers of NAFLD and fibrosis severity. Liver Int. 2019, 39, 1742–1754. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.H. Biostatistics 202: Logistic regression analysis. Singap. Med. J. 2004, 45, 149–153. [Google Scholar]
- Jager, K.J.; Zoccali, C.; MacLeod, A.; Dekker, F.W. Confounding: What it is and how to deal with it. Kidney Int. 2008, 73, 256–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| TAC ID | Gene Symbol | GAPDH | RNU6 | ||
|---|---|---|---|---|---|
| Paired | Unpaired | Paired | Unpaired | ||
| TC0100010930 | GS1-304P7.3 | 1.93 (0.027) | 2.79 (0.008) | 1.91 (0.043) | 2.18 (0.04) |
| TC0100011037 | −2.16 (0.045) | −2.58 (0.046) | −3.17 (0.006) | −3.21 (0.01) | |
| TC0100013007 | −1.12 (0.63) | −1.21 (0.44) | 1.09 (0.77) | 1.04 (0.5) | |
| TC0100015528 | 1.18 (0.86) | 1.24 (0.87) | 2.48 (0.84) | 2 (0.83) | |
| TC0100016418 | −1.31 (0.52) | −1.31 (0.51) | −1.52 (0.64) | −2.07 (0.64) | |
| TC0300007694 | NONHSAT090268 | −2.74 (0.007) | −3.04 (0.006) | −1.71 (0.007) | −3.74 (0.008) |
| TC0300013071 | zyjeebu | −1.78 (0.71) | −2.36 (0.65) | −1.77 (0.83) | −1.44 (0.82) |
| TC0400008478 | −15.75 (0.016) | −11.22 (0.003) | −63.17 (0.016) | −7.04 (0.002) | |
| TC0500012139 | peybleeby | −1.22 (0.8) | 1.17 (0.74) | −1.33 (0.68) | 1.12 (0.9) |
| TC0600007285 | HIST1H2AE | 1.06 (0.98) | 1.57 (0.98) | 1.36 (0.48) | −1.78 (0.47) |
| TC0600007784 | −1.23 (0.99) | −1.23 (0.32) | −1.31 (0.79) | 1.48 (0.81) | |
| TC0800009993 | blawker | −1.15 (0.91) | 2.43 (0.88) | 1.47 (0.72) | 1.76 (0.66) |
| TC1000010059 | NONHSAT011783 | 1.19 (0.37) | 1.85 (0.19) | 1.3 (0.88) | −1.35 (0.91) |
| TC1200011311 | NAP1L1 | 1.07 (0.99) | 4.21 (0.85) | 1.17 (0.9) | 3.9 (0.85) |
| TC1400008125 | −1.55 (0.037) | −2.25 (0.032) | −1.62 (0.021) | −2.36 (0.042) | |
| TC1600007870 | 1.08 (0.99) | −2.04 (0.99) | −1.35 (0.95) | 1.02 (0.94) | |
| TC1600010293 | swoyry | −1.27 (0.3) | −1.76 (0.5) | −1.78 (0.06) | −3.32 (0.09) |
| TC1900010363 | −2.57 (0.67) | −2.46 (0.71) | −1.59 (0.63) | 1.02 (0.56) | |
| TC2000010025 | UBE2V1 | −1.37 (0.037) | −1.63 (0.048) | −1.98 (0.028) | −2.1 (0.045) |
| DE Transcript | AUC | Std Error | P-Value | 95% CIs | Cut-Off | Sensitivity | Specificity | Accuracy | PPV | NPV |
|---|---|---|---|---|---|---|---|---|---|---|
| GS1-304P7.3 | 0.722 | 0.074 | 0.008 | 0.578–0.867 | 1.85 | 0.75 | 0.71 | 0.73 | 0.72 | 0.74 |
| NONHSAT090268 | 0.753 | 0.069 | 0.002 | 0.618–0.887 | 3.12 | 0.68 | 0.81 | 0.74 | 0.77 | 0.71 |
| TC0100011037 | 0.716 | 0.072 | 0.006 | 0.576–0.865 | 2.66 | 0.67 | 0.74 | 0.7 | 0.72 | 0.69 |
| TC0400008478 | 0.803 | 0.078 | 0.001 | 0.65–0.957 | 1.97 | 0.95 | 0.68 | 0.82 | 0.75 | 0.93 |
| TC1400008125 | 0.644 | 0.076 | 0.064 | 0.494–0.794 | 2.07 | 0.5 | 0.89 | 0.7 | 0.82 | 0.64 |
| UBE2V1 | 0.637 | 0.065 | 0.045 | 0.51–0.765 | 1.95 | 0.78 | 0.47 | 0.63 | 0.6 | 0.68 |
| Transcript Signature | P-Value | AUC | Std Error | 95% CIs | Sensitivity | Specificity | Accuracy | PPV | NPV |
|---|---|---|---|---|---|---|---|---|---|
| GS1-304P7.3, NONHSAT090268, TC0100011037, TC0400008478, TC1400008125, UBE2V1 | 0.00007 | 0.772 | 0.054 | 0.667–0.878 | 0.64 | 0.72 | 0.68 | 0.7 | 0.67 |
| Transcript | ACTB | RNU6 |
|---|---|---|
| GS1-304P7.3 | −2.55 (0.37) | −1.42 (0.51) |
| NONHSAT090268 | −2.51 (0.36) | −1.41 (0.36) |
| TC0100011037 | −2.54 (0.37) | −1.42 (0.51) |
| TC0400008478 | −2.49 (0.39) | −1.39 (0.54) |
| TC1400008125 | −3.01 (0.39) | −1.68 (0.34) |
| UBE2V1 | −1.52 (0.53) | 1.17 (0.47) |
| DE Transcript | MMSE T0 | Delta MMSE/Month |
|---|---|---|
| GS1-304P7.3 | −0.36 (0.024) | 0.29 (0.17) |
| NONHSAT090268 | 0.38 (0.012) | 0.14 (0.51) |
| TC0100011037 | 0.32 (0.03) | 0.16 (0.42) |
| TC0400008478 | 0.48 (0.02) | 0.33 (0.13) |
| TC1400008125 | 0.02 (0.85) | −0.1 (0.85) |
| UBE2V1 | −0.05 (0.67) | −0.14 (0.65) |
| Sex (M/F) | Age | MMSE T0 | MMSE T1 | Delta MMSE/Month | |
|---|---|---|---|---|---|
| AD | 17/25 | 74.51 ± 6.95 | 18.58 ± 5.4 | 14.54 ± 6.01 | −0.32 ± 0.21 |
| CTRL | 17/25 | 73.72 ± 7.34 | 29.64 ± 0.48 | N/A | N/A |
| Transcript | Forward Primer | Reverse Primer |
|---|---|---|
| ACTB | GAGCACAGAGCCTCGCCTTT | GAGCGCGGCGATATCATCA |
| blawker | AACCTGGGGCTGGTAAAGGTA | TGTGCTGCTGTTTTGGTAGTCA |
| GAPDH | TGCACCACCAACTGCTTAGC | GGCATGGACTGTGGTCATGAG |
| GS1-304P7.3 | CCAGGGACCCAGAACAGATAGT | GGTCCCTAGACACTGACGAAATC |
| HIST1H2AE | AAGAAGACGGAGAGCCACCA | GACTCGGGATCACTGACGGA |
| NAP1L1 | GGCAGACATTGACAACAAAGAAC | AGCTGACGTGCTTTGAG |
| NONHSAT011783 | TTGGTGATAGAAAAGGGCTGAAGT | GTGGCTCTCTCGGACAATGC |
| NONHSAT090268 | TCTGGCCTTACCACCTCCTTT | GAGTGGAAATGACAACTTGATGCTC |
| peybleeby | ATGGTACAGGGTGATGGGCT | GCACCCTCCCCCACCTAATA |
| RNU6 | CTCGCTTCGGCAGCACA | AACGCTTCACGAATTTGCGT |
| swoyry | TTCCTGGATGAGTGTCCTGGG | TATGGTGAGGGCAGTTGTCTCT |
| TC0100011037 | TTGAGTTAGCGAGTGGGGAGA | TGCAAATCTGGGGTTTGACCT |
| TC0100013007 | GGAAAGTCTCTGAGGAAACAGCA | GAGTAACCCATGCCTGCTCC |
| TC0100015528 | CACCTAGCCATCCCCACTGA | TTCTTTTGCTTGTGGCGTGC |
| TC0100016418 | TGACACAGGATAAGCGCAACA | CCCCCTTTACCTTCCTTGAGC |
| TC0400008478 | GCTCTGGAAAACCACAGGGTC | ATAGATCTGTGGCCAGGTGAGG |
| TC0600007784 | CCTGATCCATGCCTAGAGGTTGA | TGGAGAAACTCAATGACACCAGAAG |
| TC1400008125 | AGTTGCAAGAACGAACGGGA | CATAGGCTGGCTTGTGGAGG |
| TC1600007870 | CGCCTCTACCTCCAGTGTGA | GGCCAGAGTGGAGCCATGTA |
| TC1900010363 | AGGAGGAGACACACCCAAAAGA | GAATGCTTTTTAAGGGTGCGAGC |
| UBE2V1 | GTTGTCCTGCAAGAGCTTCG | TGTAACACTGTCCTTCGGGC |
| zyjeebu | TGTTGGCACAGTCCGTTGTC | CTCCCCTAACCTCACAGGCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbagallo, C.; Di Martino, M.T.; Grasso, M.; Salluzzo, M.G.; Scionti, F.; Cosentino, F.I.I.; Caruso, G.; Barbagallo, D.; Di Pietro, C.; Ferri, R.; et al. Uncharacterized RNAs in Plasma of Alzheimer’s Patients Are Associated with Cognitive Impairment and Show a Potential Diagnostic Power. Int. J. Mol. Sci. 2020, 21, 7644. https://doi.org/10.3390/ijms21207644
Barbagallo C, Di Martino MT, Grasso M, Salluzzo MG, Scionti F, Cosentino FII, Caruso G, Barbagallo D, Di Pietro C, Ferri R, et al. Uncharacterized RNAs in Plasma of Alzheimer’s Patients Are Associated with Cognitive Impairment and Show a Potential Diagnostic Power. International Journal of Molecular Sciences. 2020; 21(20):7644. https://doi.org/10.3390/ijms21207644
Chicago/Turabian StyleBarbagallo, Cristina, Maria Teresa Di Martino, Margherita Grasso, Maria Grazia Salluzzo, Francesca Scionti, Filomena Irene Ilaria Cosentino, Giuseppe Caruso, Davide Barbagallo, Cinzia Di Pietro, Raffaele Ferri, and et al. 2020. "Uncharacterized RNAs in Plasma of Alzheimer’s Patients Are Associated with Cognitive Impairment and Show a Potential Diagnostic Power" International Journal of Molecular Sciences 21, no. 20: 7644. https://doi.org/10.3390/ijms21207644
APA StyleBarbagallo, C., Di Martino, M. T., Grasso, M., Salluzzo, M. G., Scionti, F., Cosentino, F. I. I., Caruso, G., Barbagallo, D., Di Pietro, C., Ferri, R., Caraci, F., Purrello, M., & Ragusa, M. (2020). Uncharacterized RNAs in Plasma of Alzheimer’s Patients Are Associated with Cognitive Impairment and Show a Potential Diagnostic Power. International Journal of Molecular Sciences, 21(20), 7644. https://doi.org/10.3390/ijms21207644