TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation



Abstract
:1. Introduction
2. Results
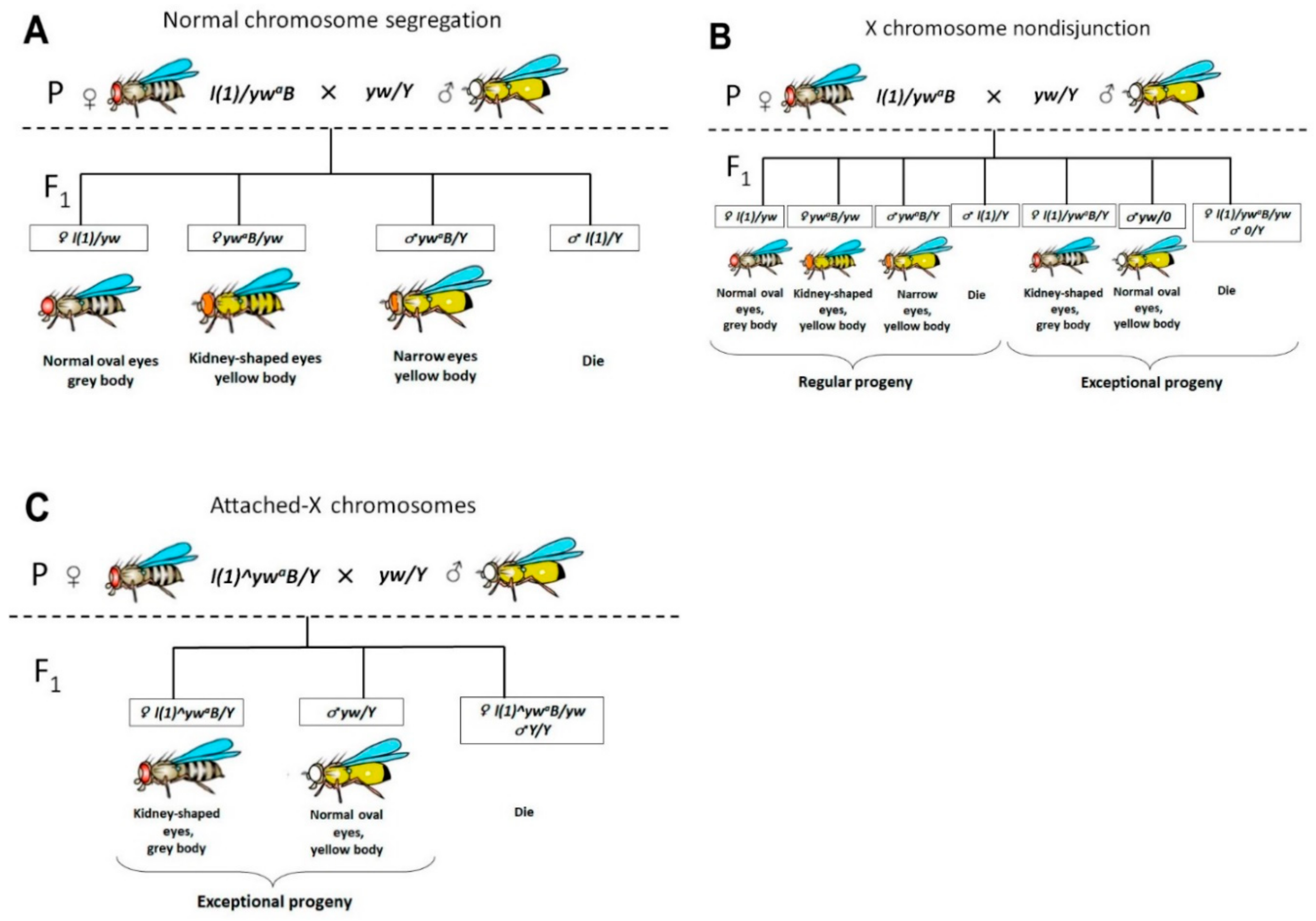
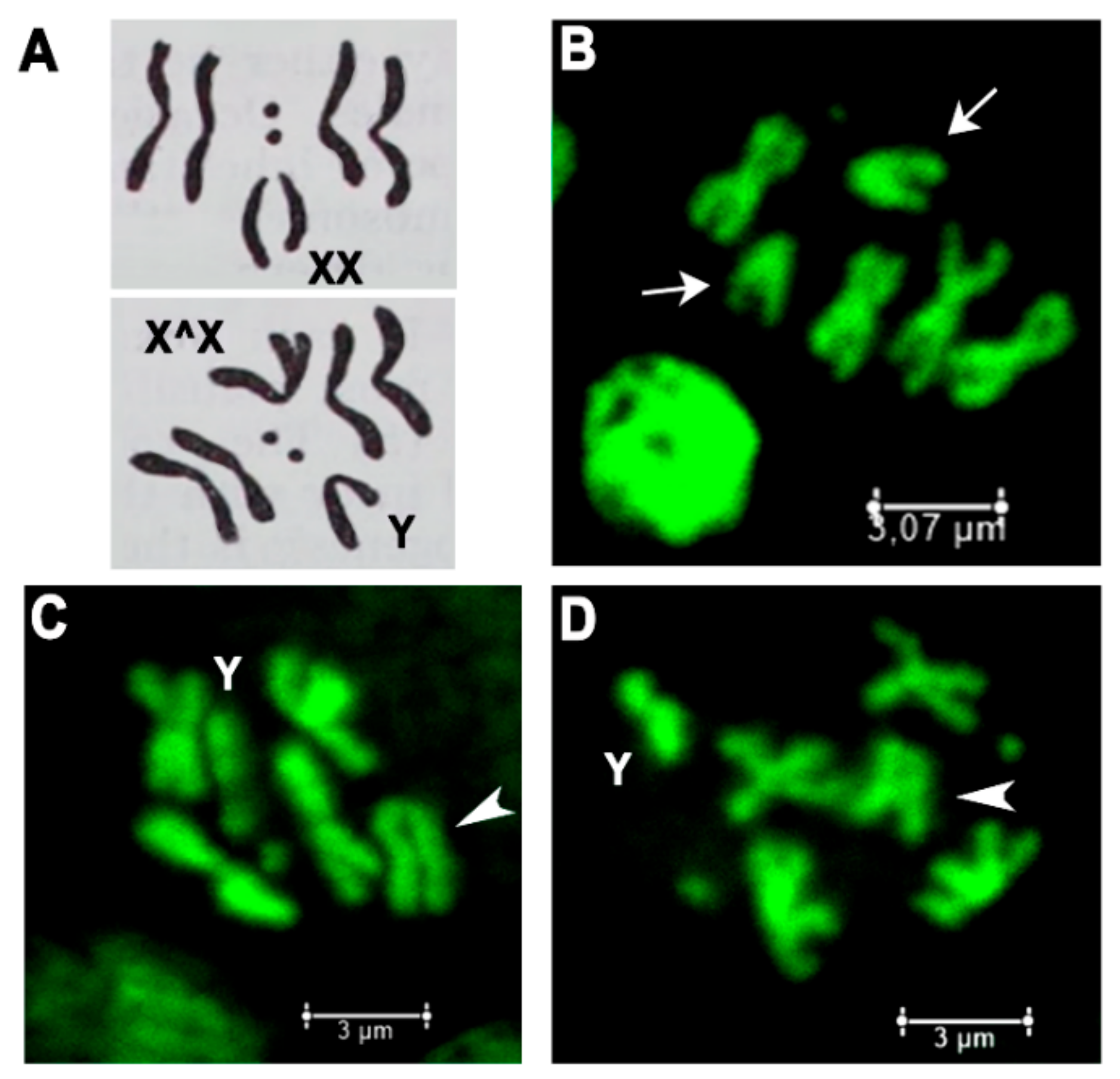
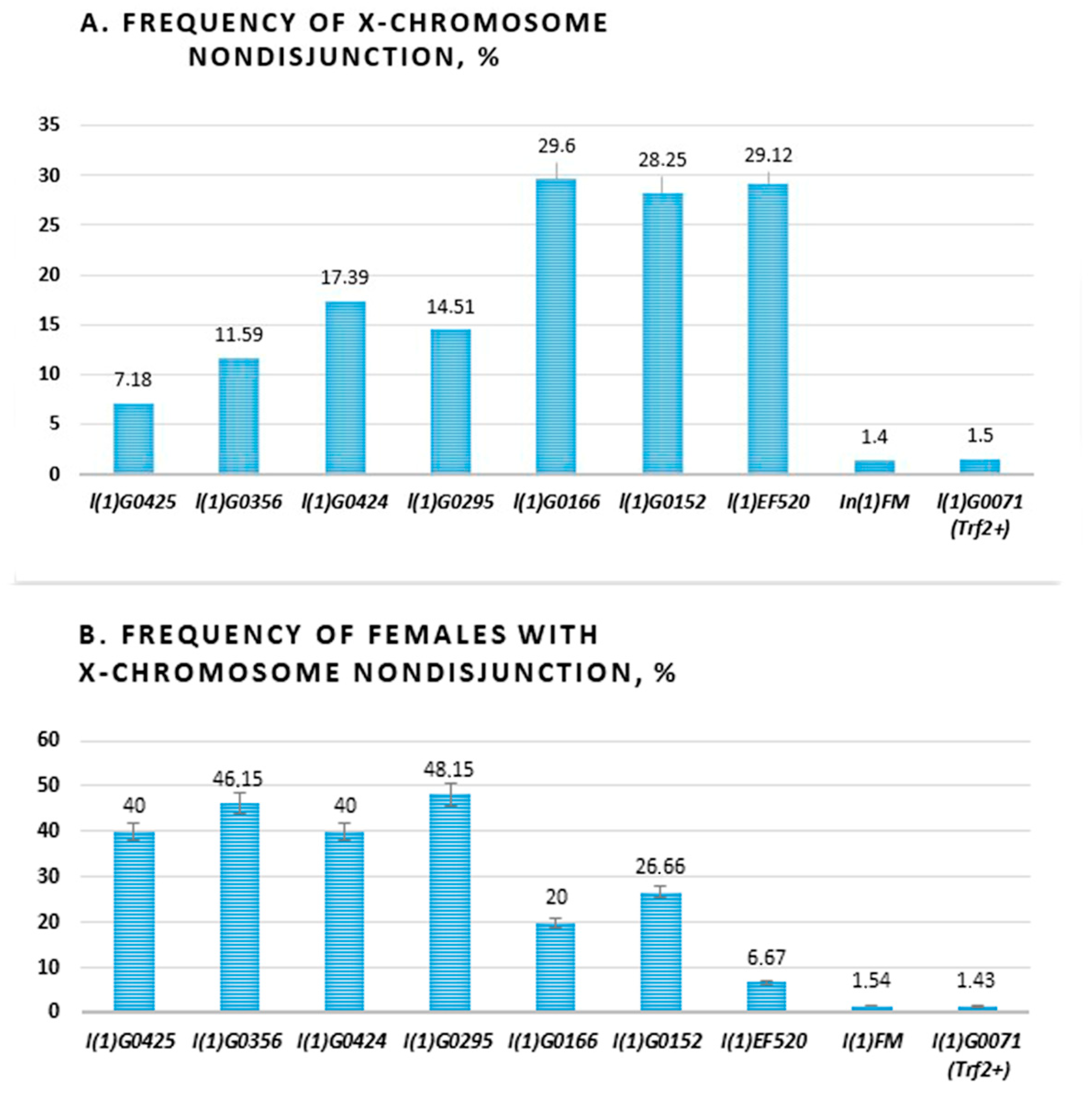
2.1. Frequency of X Chromosome Nondisjunction and Existence of Attached-X Chromosomes in Females with Lethal Trf2 Mutations
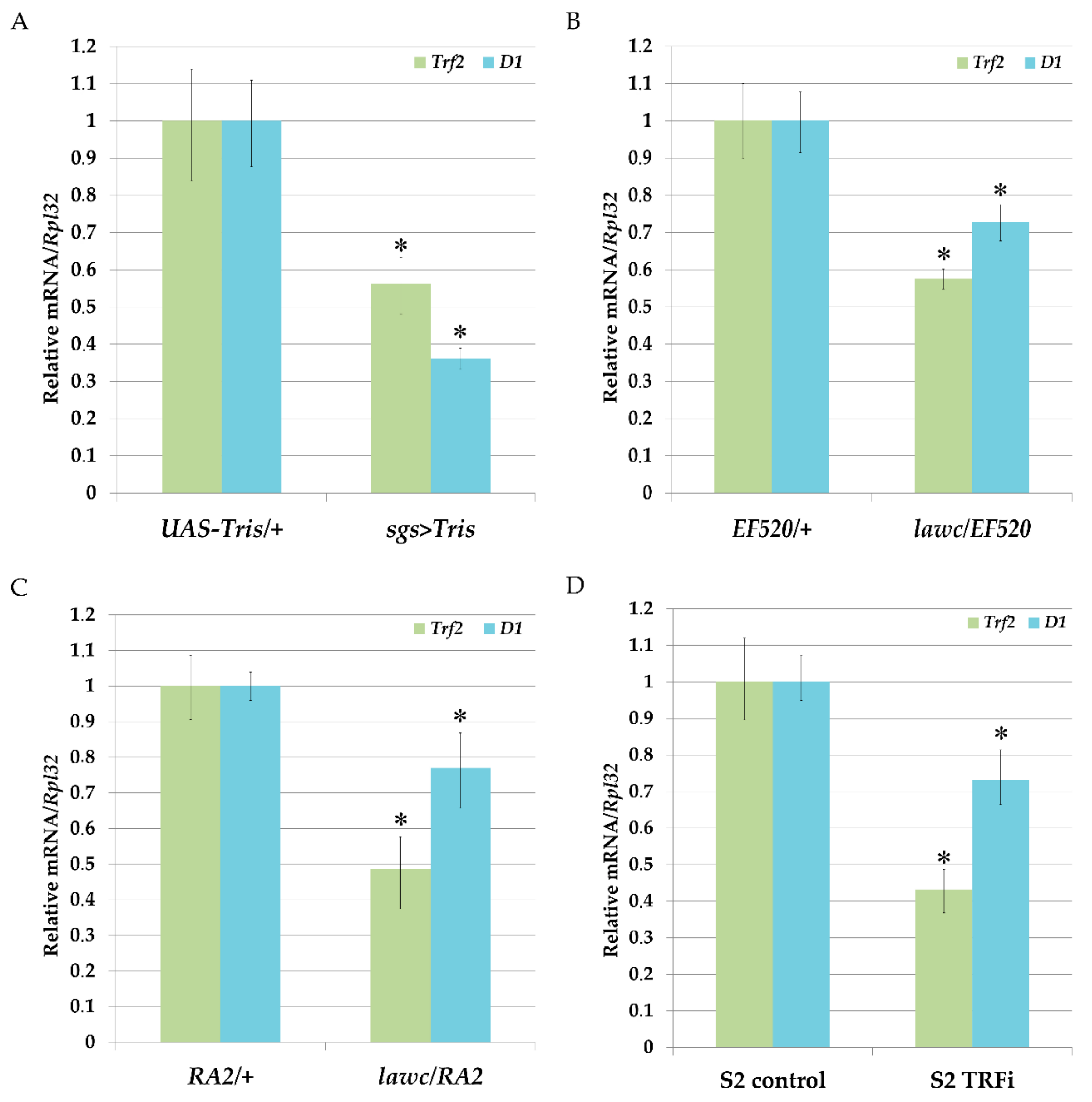
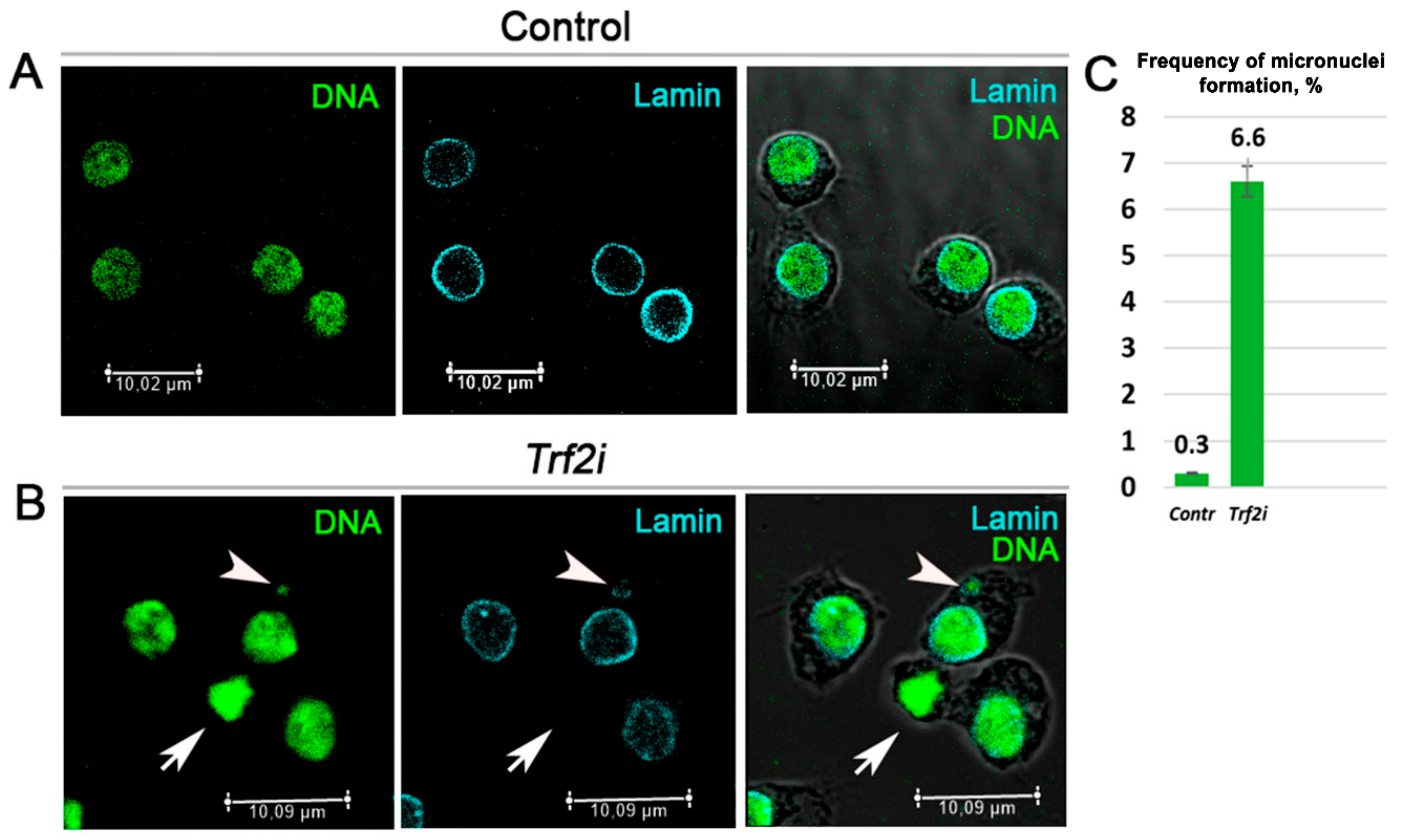
2.2. Decrease of Trf2 Gene Expression Downregulates D1 Chromosomal Protein
3. Discussion
4. Materials and Methods
4.1. Fly Stocks, Rearing Conditions and Genetic Crosses
4.2. Drosophila Melanogaster Schneider 2 (S2) Cell Culture
4.3. RNA Interference (RNAi) in Cultured Drosophila S2 Cells
4.4. Production of dsRNA for RNA Interference in Cultured Drosophila S2 Cells
4.5. RNA extraction and cDNA Preparation
4.6. Real-Time Quantitative Reverse Transcription PCR (qRT-PCR)
4.7. DNA-Directed RNA-Interference
4.8. Computational Analysis of Promoters Regions
4.9. Chromosome Cytology, Immunofluorescence Staining and Microscopy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mack, H.; Swisshelm, K. Robertsonian translocations. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 301–305. ISBN 978-0-08-096156-9. [Google Scholar]
- Stankiewicz, P.; Lupski, J.R. Genome architecture, rearrangements and genomic disorders. Trends Genet. 2002, 18, 74–82. [Google Scholar] [CrossRef]
- Garagna, S.; Page, J.; Fernandez-Donoso, R.; Zuccotti, M.; Searle, J.B. The Robertsonian phenomenon in the house mouse: Mutation, meiosis and speciation. Chromosoma 2014, 123, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Rieger, R. A new mechanism for altering chromosome number during karyotype evolution. Theor. Appl. Genet. 1985, 70, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Rieger, R.; Fuchs, J. Alteration of basic chromosome number by fusion-fission cycles. Genome 1995, 38, 1289–1292. [Google Scholar] [CrossRef]
- Jones, K. Robertsonian fusion and centric fission in karyotype evolution of higher plants. Bot. Rev 1998, 64, 273–289. [Google Scholar] [CrossRef]
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. A conserved function for pericentromeric satellite DNA. ELife 2018, 7, e34122. [Google Scholar] [CrossRef]
- Romanenko, S.A.; Lyapunova, E.A.; Saidov, A.S.; O’Brien, P.C.M.; Serdyukova, N.A.; Ferguson-Smith, M.A.; Graphodatsky, A.S.; Bakloushinskaya, I. Chromosome translocations as a driver of diversification in mole voles ellobius (rodentia, mammalia). Int. J. Mol. Sci. 2019, 20, 4466. [Google Scholar] [CrossRef] [Green Version]
- Sember, A.; de Oliveira, E.A.; Ráb, P.; Bertollo, L.A.C.; de Freitas, N.L.; Viana, P.F.; Yano, C.F.; Hatanaka, T.; Marinho, M.M.F.; de Moraes, R.L.R.; et al. Centric fusions behind the karyotype evolution of neotropical nannostomus pencilfishes (characiforme, lebiasinidae): First insights from a molecular cytogenetic perspective. Genes 2020, 11, 91. [Google Scholar] [CrossRef] [Green Version]
- Matveevsky, S.; Kolomiets, O.; Bogdanov, A.; Alpeeva, E.; Bakloushinskaya, I. Meiotic chromosome contacts as a plausible prelude for robertsonian translocations. Genes 2020, 11, 386. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; O’Brien, P.C.; Milne, B.S.; Graphodatsky, A.S.; Solanky, N.; Trifonov, V.; Rens, W.; Sargan, D.; Ferguson-Smith, M.A. A complete comparative chromosome map for the dog, red fox, and human and its integration with canine genetic maps. Genomics 1999, 62, 189–202. [Google Scholar] [CrossRef]
- Ferguson-Smith, M.A.; Trifonov, V. Mammalian karyotype evolution. Nat. Rev. Genet. 2007, 8, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Kirsch-Volders, M.; Cundari, E.; Verdoodt, B. Towards a unifying model for the metaphase/anaphase transition. Mutagenesis 1998, 13, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.E.; Miller, D.E.; Miller, A.L.; Hawley, R.S. Female meiosis: Synapsis, recombination, and segregation in drosophila melanogaster. Genetics 2018, 208, 875–908. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.S.; Holland, A.J. The impact of mitotic errors on cell proliferation and tumorigenesis. Genes Dev. 2018, 32, 620–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, L.; Voisin, M.; Tatout, C.; Probst, A.V. Structure and function of centromeric and pericentromeric heterochromatin in arabidopsis thaliana. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Zowalaty, A.E.; Baumann, C.; Li, R.; Chen, W.; De La Fuente, R.; Ye, X. Seipin deficiency increases chromocenter fragmentation and disrupts acrosome formation leading to male infertility. Cell Death Dis. 2015, 6, e1817. [Google Scholar] [CrossRef] [Green Version]
- Belli, M.; Vigone, G.; Merico, V.; Redi, C.A.; Garagna, S.; Zuccotti, M. Time-lapse dynamics of the mouse oocyte chromatin organisation during meiotic resumption. BioMed Res. Int. 2014, 2014, 207357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalenskaya, I.A.; Zalensky, A.O. Non-random positioning of chromosomes in human sperm nuclei. Chromosome Res. 2004, 12, 163–173. [Google Scholar] [CrossRef]
- Funabiki, H.; Hagan, I.; Uzawa, S.; Yanagida, M. Cell cycle-dependent specific positioning and clustering of centromeres and telomeres in fission yeast. J. Cell Biol. 1993, 121, 961–976. [Google Scholar] [CrossRef] [Green Version]
- Probst, A.V.; Okamoto, I.; Casanova, M.; El Marjou, F.; Le Baccon, P.; Almouzni, G. A strand-specific burst in transcription of pericentric satellites is required for chromocenter formation and early mouse development. Dev. Cell 2010, 19, 625–638. [Google Scholar] [CrossRef] [Green Version]
- Chubykin, V.L. The structure of the chromocenter in the oocytes, the initiation of homologous pairing and the regulation of the formation of crossing over in drosophila. Tsitologiia 1995, 37, 481–490. [Google Scholar] [PubMed]
- Zuccotti, M.; Garagna, S.; Merico, V.; Monti, M.; Alberto Redi, C. Chromatin organisation and nuclear architecture in growing mouse oocytes. Mol. Cell. Endocrinol. 2005, 234, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Chubykin, V.L. Role of the chromocenter in nonrandom meiotic segregation of nonhomologous chromosomes in drosophila melanogaster females. Russ. J. Genet. 2001, 37, 205–212. [Google Scholar] [CrossRef]
- Chubykin, V.L. Genetic control of the formation and reorganization of chromocenter in drosophila. Russ. J. Genet. 2001, 37, 888–893. [Google Scholar] [CrossRef]
- Fedorova, S.; Nokkala, S.; Chubykin, V.; Omelyanchuk, L. The isolation of a mutation causing abnormal cytokinesis in male and split chromocenter in female meiosis in drosophila melanogaster. Hereditas 2001, 134, 125–134. [Google Scholar] [CrossRef]
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. The modular mechanism of chromocenter formation in drosophila. Elife 2019, 8. [Google Scholar] [CrossRef]
- Rabenstein, M.D.; Zhou, S.; Lis, J.T.; Tjian, R. TATA box-binding protein (TBP)-related factor 2 (TRF2), a third member of the TBP family. Proc. Natl. Acad. Sci. USA 1999, 96, 4791–4796. [Google Scholar] [CrossRef] [Green Version]
- Simonova, O.B.; Kuzin, B.A.; Georgiev, P.G.; Gerasimova, T.I. Novel regulatory mutation in drosophila melanogaster. Russ. J. Genet. 1992, 28, 164–167. [Google Scholar]
- Vorontsova, J.E.; Modestova, E.A.; Burdina, N.V.; Korochkin, L.I.; Simonova, O.B. Restoring viability of lethal mutants for the leg-arista-wing-complex gene in rescue experiments with transgenic constructs that express the trf2 gene domains in drosophila melanogaster. Dokl. Biol. Sci. 2007, 417, 429–431. [Google Scholar] [CrossRef]
- Simonova, O.B.; Petruk, S.F.; Dzhagaeva, I.V.; Korochkin, L.I. Role of the lawc(p1) mutation in the regulation of the white locus expression in drosophila. Genetika 1998, 34, 349–354. [Google Scholar]
- Wu, C.T. Transvection, nuclear structure, and chromatin proteins. J. Cell Biol. 1993, 120, 587–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopytova, D.V.; Krasnov, A.N.; Kopantceva, M.R.; Nabirochkina, E.N.; Nikolenko, J.V.; Maksimenko, O.; Kurshakova, M.M.; Lebedeva, L.A.; Yerokhin, M.M.; Simonova, O.B.; et al. Two isoforms of drosophila TRF2 are involved in embryonic development, premeiotic chromatin condensation, and proper differentiation of germ cells of both sexes. Mol. Cell. Biol. 2006, 26, 7492–7505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martianov, I.; Brancorsini, S.; Gansmuller, A.; Parvinen, M.; Davidson, I.; Sassone-Corsi, P. Distinct functions of TBP and TLF/TRF2 during spermatogenesis: Requirement of TLF for heterochromatic chromocenter formation in haploid round spermatids. Development 2002, 129, 945–955. [Google Scholar] [PubMed]
- Vorontsova, Y.E.; Cherezov, R.O.; Simonova, O.B. Effect of mutations in lawc/Trf2 gene on development of chromocenter and chromosome disjunction in drosophila melanogaster. Genetika 2013, 49, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Kedmi, A.; Zehavi, Y.; Glick, Y.; Orenstein, Y.; Ideses, D.; Wachtel, C.; Doniger, T.; Waldman Ben-Asher, H.; Muster, N.; Thompson, J.; et al. Drosophila TRF2 is a preferential core promoter regulator. Genes Dev. 2014, 28, 2163–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, P.J.; Greenman, C.D.; Fu, B.; Yang, F.; Bignell, G.R.; Mudie, L.J.; Pleasance, E.D.; Lau, K.W.; Beare, D.; Stebbings, L.A.; et al. Massive genomic rearrangement acquired in a single catastrophic event during cancer development. Cell 2011, 144, 27–40. [Google Scholar] [CrossRef]
- Vorontsova, J.; Cherezov, R.; Simonova, O. The effect of TBP-related factor 2 on chromocenter formation and chromosome segregation in drosophila melanogaster. Chromosom. Abnorm. A Hallmark Manif. Genom. Instab. 2017. [Google Scholar] [CrossRef] [Green Version]
- Andersen, P.R.; Tirian, L.; Vunjak, M.; Brennecke, J. A heterochromatin-dependent transcription machinery drives piRNA expression. Nature 2017, 549, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-interacting small RNAs: The vanguard of genome defence. Nat. Rev. Mol. Cell Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef]
- Barra, V.; Fachinetti, D. The dark side of centromeres: Types, causes and consequences of structural abnormalities implicating centromeric DNA. Nat. Commun. 2018, 9, 4340. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Pellman, D. The coordination of nuclear envelope assembly and chromosome segregation in metazoans. Nucleus 2020, 11, 35–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehavi, Y.; Kedmi, A.; Ideses, D.; Juven-Gershon, T. TRF2: TRansForming the view of general transcription factors. Transcription 2015, 6, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duttke, S.H.C.; Doolittle, R.F.; Wang, Y.-L.; Kadonaga, J.T. TRF2 and the evolution of the bilateria. Genes Dev. 2014, 28, 2071–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, A.; Schöttler, P.; Werner, M.; Beinert, N.; Dowe, G.; Burkert, P.; Mourkioti, F.; Dentzer, L.; He, Y.; Deak, P.; et al. Mapping and identification of essential gene functions on the X chromosome of drosophila. EMBO Rep. 2002, 3, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Zitron, A.E.; Hawley, R.S. The genetic analysis of distributive segregation in drosophila melanogaster. I. isolation and characterization of aberrant X segregation (Axs), a mutation defective in chromosome partner choice. Genetics 1989, 122, 801–821. [Google Scholar]
- Clemens, J.C.; Worby, C.A.; Simonson-Leff, N.; Muda, M.; Maehama, T.; Hemmings, B.A.; Dixon, J.E. Use of double-stranded RNA interference in Drosophila cell lines to dissect signal transduction pathways. Proc. Natl. Acad. Sci. USA 2000, 97, 6499–6503. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Comjean, A.; Roesel, C.; Vinayagam, A.; Flockhart, I.; Zirin, J.; Perkins, L.; Perrimon, N.; Mohr, S.E. FlyRNAi.org-the database of the drosophila RNAi screening center and transgenic RNAi project: 2017 update. Nucleic Acids Res. 2017, 45, D672–D678. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Arikawa, E.; Sun, Y.; Wang, J.; Zhou, Q.; Ning, B.; Dial, S.L.; Guo, L.; Yang, J. Cross-platform comparison of SYBR green real-time PCR with TaqMan PCR, microarrays and other gene expression measurement technologies evaluated in the microarray quality control (MAQC) study. BMC Genom. 2008, 9, 328. [Google Scholar] [CrossRef] [Green Version]
- Tajadini, M.; Panjehpour, M.; Javanmard, S.H. Comparison of SYBR green and TaqMan methods in quantitative real-time polymerase chain reaction analysis of four adenosine receptor subtypes. Adv. BioMed Res. 2014, 3. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.A.; Giniger, E.; Maniatis, T.; Ptashne, M. GAL4 activates transcription in drosophila. Nature 1988, 332, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [PubMed]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherezov, R.O.; Vorontsova, I.E.; Mertsalov, I.B.; Kulikova, D.A.; Simonova, O.B. Influence of hairpin RNA, specific to the gene lawc, on expression of overlapping genes of the lawc/Trf2 complex in D. melanogaster. Izv. Akad. Nauk. Ser. Biol. 2013, 2, 133–137. [Google Scholar]
- Phelps, C.B.; Brand, A.H. Ectopic gene expression in drosophila using GAL4 system. Methods 1998, 14, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Reese, M.G. Application of a time-delay neural network to promoter annotation in the drosophila melanogaster genome. Comput. Chem. 2001, 26, 51–56. [Google Scholar] [CrossRef]
- Sloutskin, A.; Danino, Y.M.; Orenstein, Y.; Zehavi, Y.; Doniger, T.; Shamir, R.; Juven-Gershon, T. ElemeNT: A computational tool for detecting core promoter elements. Transcription 2015, 6, 41–50. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stocks | Attached X | Qn, % (N) | Qf, % | Total |
|---|---|---|---|---|
| l(1)G0425/In(1)FM | + | 7.17 (779) | 40 | 25 |
| l(1)G0356/In(1)FM | + | 11.59 (845) | 46.15 | 26 |
| l(1)G0424/In(1)FM | − | 17.39 (777) | 40 | 25 |
| l(1)G0295/In(1)FM | − | 14.51 (1201) | 48.15 | 27 |
| l(1)G0166/In(1)FM | − | 29.6 (236) | 20 | 30 |
| l(1)G0152/In(1)FM | − | 28.25 (370) | 26.66 | 30 |
| l(1)EF520/In(1)FM | − | 29.12 (88) | 6.67 | 30 |
| Control stocks | ||||
| In(1)FM | − | 1.4 (2445) | 1.54 | 65 |
| l(1)G0071/In(1)FM | − | 1.5 (650) | 1.43 | 70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cherezov, R.O.; Vorontsova, J.E.; Simonova, O.B. TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation. Int. J. Mol. Sci. 2020, 21, 8871. https://doi.org/10.3390/ijms21228871
Cherezov RO, Vorontsova JE, Simonova OB. TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation. International Journal of Molecular Sciences. 2020; 21(22):8871. https://doi.org/10.3390/ijms21228871
Chicago/Turabian StyleCherezov, Roman O., Julia E. Vorontsova, and Olga B. Simonova. 2020. "TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation" International Journal of Molecular Sciences 21, no. 22: 8871. https://doi.org/10.3390/ijms21228871
APA StyleCherezov, R. O., Vorontsova, J. E., & Simonova, O. B. (2020). TBP-Related Factor 2 as a Trigger for Robertsonian Translocations and Speciation. International Journal of Molecular Sciences, 21(22), 8871. https://doi.org/10.3390/ijms21228871