Role of Bioactive Peptide Sequences in the Potential Impact of Dairy Protein Intake on Metabolic Health

Abstract
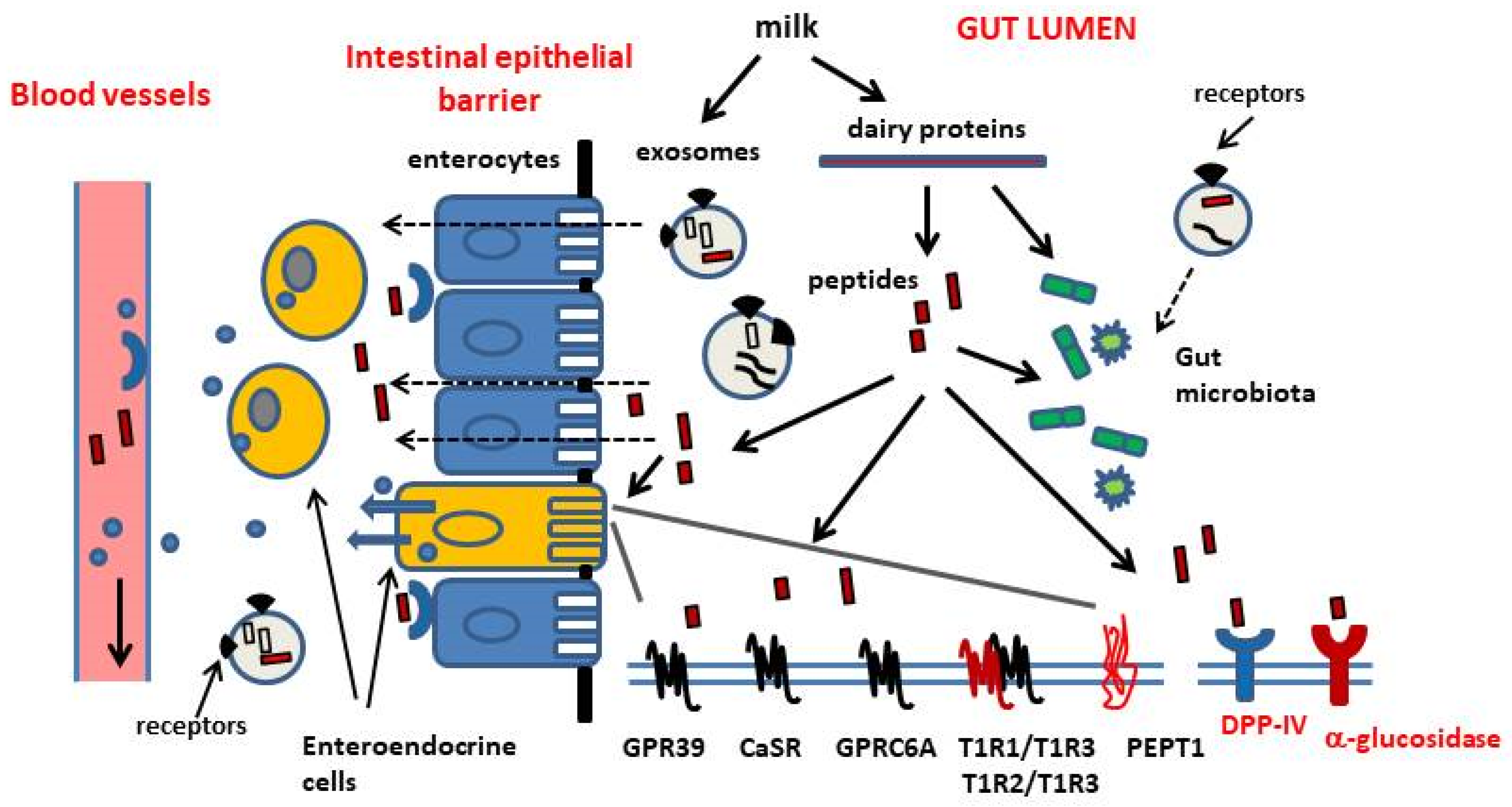
:1. Introduction
1.1. Nutrient-Sensing Receptors
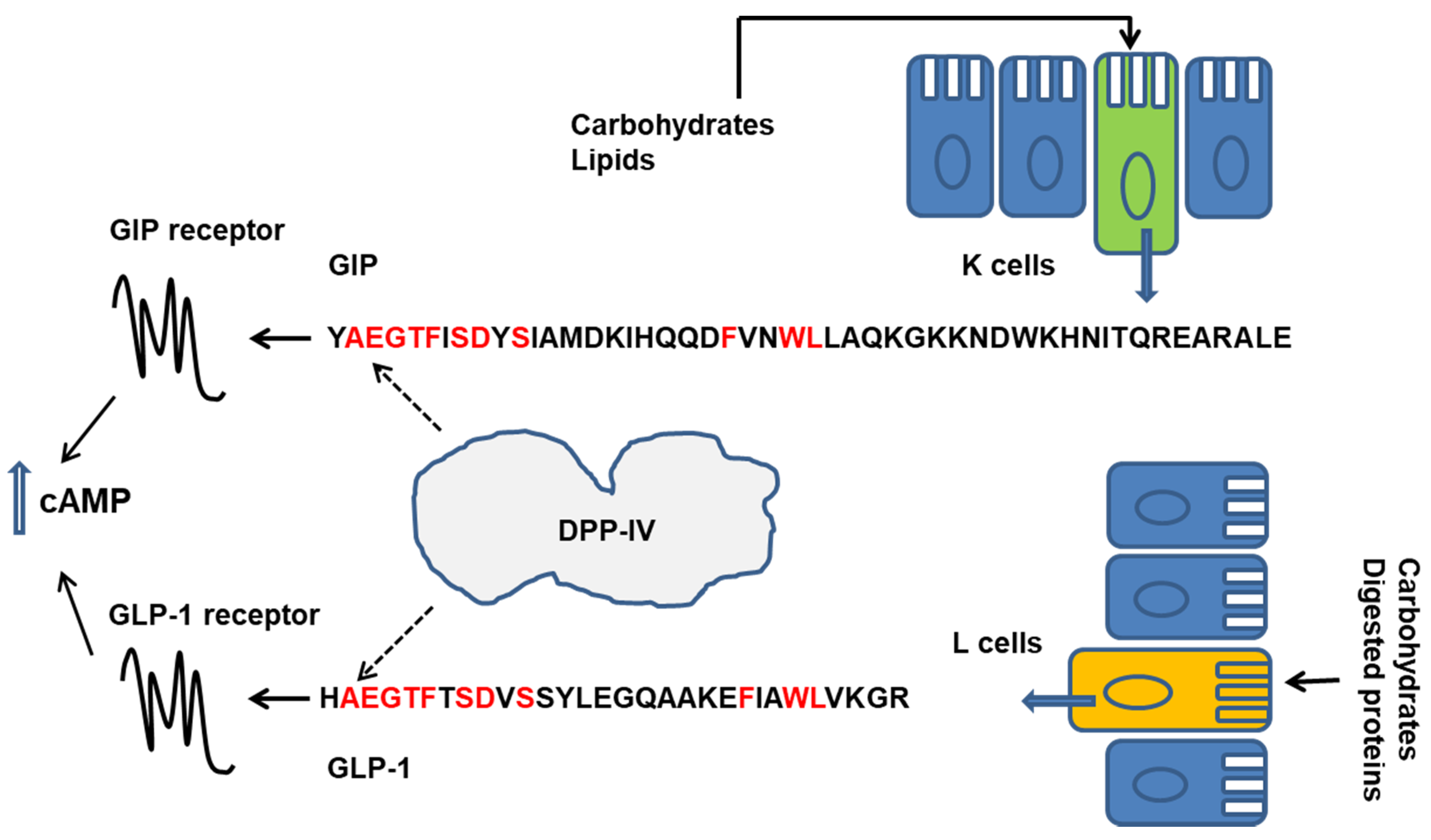
1.2. Gastroenteric Hormones
2. How Dairy Proteins Can Modulate Food Intake and Glucose Homeostasis
2.1. Role of Free Amino Acid as Metabolic Signals
2.2. Evidence Supporting a Role of Bioactive Peptides
3. Final Considerations and Perspectives
Funding
Conflicts of Interest
Abbreviations
| α-LA | α-lactalbumin |
| β-CM7 | β-casomorphin-7 |
| β-LG | β-lactoglobulin |
| CCK | cholecystokinin |
| CMP | caseinomacropeptide |
| DPP-IV | dipeptidyl peptidase-IV |
| GCPR | G-coupled protein receptors |
| GI | gastrointestinal |
| GIP | glucose-dependent insulinotropic peptide |
| GMP | glycomacropeptide |
| GLP-1 | glucagon-like peptide-1 |
| ip | intraperitoneal |
| os | oral |
| WPC | whey proteinconcentrate |
| WPI | whey protein isolate |
References
- Ryann, K.K.; Seeley, R.M. Food as a hormone. Science 2013, 339, 918–919. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, A.; Vazquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–969. [Google Scholar] [CrossRef]
- Hernandez-Ledesma, B.; Garcia-Nebot, M.J.; Fernandez-Tomè, S.; Amigo, L.; Recio, I. Dairy protein hydrolysates: Peptides for health benefits. Int. Dairy J. 2014, 38, 82–100. [Google Scholar] [CrossRef] [Green Version]
- Smithers, G.W. Whey and whey proteins—from gutter-to-gold. Int. Dairy J. 2008, 18, 695–704. [Google Scholar] [CrossRef]
- Bendtsen, L.Q.; Lorenzen, J.K.; Bendsen, N.T.; Rasmussen, C.; Asrup, A. Effect of dairy protein on appetite, energy expenditure, body weight and composition: A review of the evidence from controlled clinical trials. Adv. Nutr. 2013, 4, 418–438. [Google Scholar] [CrossRef] [Green Version]
- Kondrashina, A.; Brodkorp, A.; Giblin, L. Dairy-derived peptides for satiety. J. Funct. Foods 2020, 66, 103801. [Google Scholar] [CrossRef]
- Pesta, D.H.; Samuel, V.T. A high-protein diet for reducing body fat: Mechanisms and possible caveats. Nutr. Metab. 2014, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Rigamonti, A.E.; Leoncini, R.; De Col, A.; Tamini, S.; Cicolini, S.; Abbruzzese, L.; Cella, S.G.; Sartorio, A. The Appetite-suppressant and GLP-1-Stimulating effects of whey proteins in obese subjects are associated with increasing circulating levels of specific amino acids. Nutrients 2020, 12, 775. [Google Scholar] [CrossRef] [Green Version]
- Goodman, B.E. Insights into digestion and absorption of major nutrients in humans. Adv. Physiol. Educ. 2010, 34, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Santos-Hernandez, M.S.; Miralles, B.; Amigo, L.; Recio, I. Intestinal signals of proteins and digestion-derived products relevant to satiety. J. Agric. Food Chem. 2018, 66, 10123–10131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luhovyy, B.L.; Akhavan, T.; Anderson, G.H. Whey proteins in the regulation of food intake and satiety. J. Am. Coll. Nutr. 2007, 26, 704S–712S. [Google Scholar] [CrossRef] [PubMed]
- Westerterp-Plantenga, M.S.; Lemmens, S.G.; Westerterp, K.R. Dietary protein—Its role in satiety, energetics, weight loss and health. Br. J. Nutr. 2012, 108 (Suppl. S2), S105–S112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilaweera, K.N.; Cabrera-Rubio, R.; Speakman, J.R.; O’Connor, P.M.; McAulie, A.; Guinane, C.M.; Lawton, E.M.; Crispie, F.; Aguilera, M.; Stanley, M.; et al. Whey protein effects on energy balance link the intestinal mechanisms of energy absorption with adiposity and hypothalamic neuropeptide gene expression. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E1–E11. [Google Scholar] [CrossRef]
- Giezenaar, C.; Trahair, L.G.; Luscombe-Marsh, N.D.; Hausken, T.; Standfield, S.; Jones, J.K.; Lange, K.; Horowitz, M.; Chapman, I.; Soenen, S. Effects of randomized whey-protein loads on energy intake, appetite, gastric emptying, and plasma gut hormone concentrations in older men and women. Am. J. Clin. Nutr. 2017, 106, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Giezenaar, C.; Luscombe-Marsh, N.D.; Hutchison, A.T.; Standfield, S.; Feinle-Bisset, C.; Horowitz, M.; Chapman, I.; Soenen, S. Dose-Dependent effects of randomized intraduodenal whey-protein Loads on glucose, gut hormone, and amino acid concentrations in healthy older and younger men. Nutrients 2018, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Kassis, A.; Godin, J.P.; Moille, S.E.; Nielsen-Moennoz, C.; Groulx, K.; Oguey-Araymon, S.; Praplan, F.; Beaumont, M.; Sauser, J.; Monnard, I.; et al. Effects of protein quantity and type on diet induced thermogenesis in overweight adults: A randomized controlled trial. Clin. Nutr. 2019, 38, 1570–1580. [Google Scholar] [CrossRef]
- Anuradha, K.V. Amino acid support in the prevention of diabetes and diabetic complications. Curr. Protein Pept. Sci. 2009, 10, 8–17. [Google Scholar] [CrossRef]
- Chungchunlam, S.M.; Henare, S.J.; Ganesh, S.; Moughan, P.J. Dietary whey protein influences plasma satiety-related hormones and plasma amino acids in norma-weight adult women. Eur. J. Clin. Nutr. 2015, 69, 179–186. [Google Scholar] [CrossRef]
- Tulipano, G.; Faggi, L.; Caroli, A.M.; Cocchi, D. Whey proteins as functional food affecting glycemic control: Molecular determinants. In Whey: Types, Composition and Health Implications; Benitez, R.M., Ortero, G.M., Eds.; Nova Science Publishers: New York, NY, USA; Huntington: New York, NY, USA, 2012; Chapter 3; pp. 93–114. [Google Scholar]
- Reimann, F. Molecular mechanisms underlying nutrient detection by incretin-secreting cells. Int. Dairy J. 2010, 20, 236–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troke, R.C.; Tan, T.M.; Bloom, S.R. The future role of gut hormones in the treatment of obesity. Ther. Adv. Chronic Dis. 2014, 5, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tulipano, G. Whey proteins and enteric hormones: Interplay between food components and the enteroendocrine system. Agro Food Ind. Hi-Tech. 2015, 26 (Suppl. S2), 15–19. [Google Scholar]
- Wauson, E.M.; Lorente-Rodriguez, A.; Cobb, M.H. Minireview: Nutrient sensing by G protein-coupled receptors. Mol. Endocrinol. 2013, 27, 1188–1197. [Google Scholar] [CrossRef] [Green Version]
- Liou, A.P.; Chavez, D.L.; Espero, E.; Hao, S.; Wank, S.A.; Raybould, H.E. Protein hydrolysate-induced cholecystokinin secretion from enteroendocrine cells is indirectly mediated by the intestinal oligopeptide transporter PepT1. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G895–G902. [Google Scholar] [CrossRef] [Green Version]
- Ezcurra, M.; Reimann, F.; Gribble, F.M.; Emery, E. Molecular mechanisms of incretin hormone secretion. Curr. Opin. Pharmacol. 2013, 13, 922–927. [Google Scholar] [CrossRef] [Green Version]
- Daly, K.; Al-Rammahi, M.; Moran, A.; Marcello, M.; Ninomiya, Y.; Shirazi-Beechey, S.P. Sensing of amino acids by the gut expressed taste receptor T1R1-T1R3 stimulates CCK secretion. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G271–G282. [Google Scholar] [CrossRef] [Green Version]
- Geraedts, M.C.P.; Troost, F.; Fisher, M.A.J.G.; Edens, L.; Saris, W.H.M. Direct induction of CCK and GLP-1 release from murine endocrine cells by intact dietary proteins. Mol. Nutr. Food Res. 2011, 55, 476–484. [Google Scholar] [CrossRef]
- Tulipano, G.; Faggi, L.; Cacciamali, A.; Caroli, A.M. Food protein-derived peptide sensing by enteroendocrine cells compared to osteoblast-like cells: Role of peptide length and peptide composition, focusing on products of beta-lactoglobulin hydrolysis. Int. Dairy J. 2017, 72, 55–62. [Google Scholar] [CrossRef]
- Wang, Y.; Chandra, R.; Samsa, L.A.; Gooch, B.; Fee, B.E.; Cook, J.M.; Vigna, S.R.; Grant, A.O.; Liddle, R.A. Amino acids stimulate cholecystokinin release through the Ca2+ sensing receptor. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G528–G537. [Google Scholar] [CrossRef] [Green Version]
- Hira, T.; Nakajima, S.; Eto, Y.; Hara, H. Calcium-sensing receptor mediates phenylalanine-induced cholecystokinin secretion in enteroendocrine STC-1 cells. FEBS J. 2008, 275, 4620–4626. [Google Scholar] [CrossRef] [PubMed]
- Wellendorph, P.; Brauner-Osborne, H. Molecular cloning, expression and sequence analysis of GPRC6A, a novel family C G-protein-coupled receptor. Gene 2004, 335, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, B.; Hansen, K.B.; Wellendorph, P.; Breauner-Osborne, H. Pharmacological characterization of mouse GPRC6A: A promiscuous l-alpha-amino acid receptor with prevalence for basic amino acids. Br. J. Pharmacol. 2005, 67, 589–597. [Google Scholar]
- Mizokami, A.; Yasutake, Y.; Gao, J.; Matsuda, M.; Takahashi, I.; Takeuchi, H.; Hirata, M. Osteocalcin induces release of glucagon-like peptide-1 and thereby stimulates insulin secretion in mice. PLoS ONE 2013, 8, e57375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Lee, M.; Shiu, A.L.; Yo, S.J.; Hallden, G.; Aponte, G.W. GPR93 activation by protein hydrolysate induces CCK transcription and secretion in STC-1 cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1366–G1375. [Google Scholar] [CrossRef]
- Goltzman, D.; Hendy, G.N. The calcium-sensing receptor in bone--mechanistic and therapeutic insights. Nature Reviews. Endocrinology 2015, 11, 298–307. [Google Scholar]
- Pi, M.; Faber, P.; Ekema, G.; Jackson, P.D.; Ting, A.; Wang, N.; Fontilla-Poole, M.; Mays, R.W.; Brunden, K.R.; Harrington, J.J.; et al. Identification of a novel extracellular cation-sensing G-protein-coupled receptor. J. Biol. Chem. 2005, 280, 40201–40209. [Google Scholar] [CrossRef] [Green Version]
- Pi, M.; Zhang, L.; Lei, S.F.; Huang, M.Z.; Zhu, W.; Zhang, J.; Shen, H.; Deng, H.W.; Quarles, L.D. Impaired osteoblast function in GPRC6A null mice. J. Bone Miner. Res. 2010, 25, 1092–1102. [Google Scholar]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Mishra, A.K.; Dubey, V.; Ghosh, A.R. Obesity: An overview of possible role(s) of gut hormones, lipid sensing and gut microbiota. Metabolism 2016, 65, 48–65. [Google Scholar] [CrossRef]
- Miller, L.J.; Desai, A.J. Metabolic actions of the Type 1 Cholecystokinin Receptor: Its potential as a therapeutic target. Trends Endocrinol. Metab. 2016, 27, 609–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, J.E.; Drucker, D.J. Pharmacology, physiology and mechanisms of incretin hormone action. Cell Metab. 2013, 17, 819–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raschke, S.; Eckardt, K.; Bjørklund Holven, K.; Jensen, J.; Eckel, J. Identification and validation of novel contraction regulated myokines released from primary human skeletal muscle cells. PLoS ONE 2013, 8, e62008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darmoul, D.; Voisin, T.; Couvineau, A.; Rouyer-Fessard, C.; Salomon, R.; Wang, Y.; Swallow, D.M.; Laburthe, M. Regional expression of epithelial dipeptidyl peptidase IV in the human intestines. BioChem. Biophys. Res. Commun. 1994, 203, 1224–1229. [Google Scholar] [CrossRef]
- Holst, J.J.; Deacon, C.F. Inhibition of the activity of dipeptidyl-peptidase IV as a treatment for type 2 diabetes. Diabetes 1998, 47, 1663–1670. [Google Scholar] [CrossRef]
- Gunnarsson, P.T.; Winzell, M.S.; Deacon, C.F.; Larsen, M.O.; Jelic, K.; Carr, R.D.; Ahren, B. Glucose-induced incretin hormone release and inactivation are differently modulated bu oral fat and protein in mice. Endocrinology 2006, 147, 3173–3180. [Google Scholar] [CrossRef] [Green Version]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Tulipano, G.; Sibilia, V.; Caroli, A.M.; Cocchi, D. Whey proteins as source of dipeptidyl dipeptidase IV (dipeptidyl peptidase-4) inhibitors. Peptides 2011, 32, 835–838. [Google Scholar] [CrossRef]
- Tulipano, G.; Faggi, L.; Nardone, A.; Cocchi, D.; Caroli, A.M. Characterisation of the potential of beta-lactoglobulin and alpha-lactalbumin as sources of bioactive peptides affecting incretin function: In silico and in vitro comparative studies. Int. Dairy J. 2015, 48, 66–72. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Isolation and characterization of peptides with dipeptidyl peptidase-IV inhibitory activity from pepsin-treated bovine whey proteins. Peptides 2014, 54, 39–48. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Structure activity relationship modelling of milk protein-derived peptides with dipeptidyl peptidase IV (DPP-IV) inhibitory activity. Peptides 2016, 79, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnett, M.P.; McNabb, W.C.; Roy, N.C.; Woodford, K.B.; Clarke, A.J. Dietary A1 β-casein affects gastrointestinal transit time, dipeptidyl peptidase-4 activity, and inflammatory status relative to A2 β-casein in Wistar rats. Int. J. Food Sci. Nutr. 2014, 65, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Howick, K.; Griffin, B.; Cryan, J.; Schellekens, H. From Belly to Brain: Targeting the ghrelin receptor in appetite and food intake regulation. Int. J. Mol. Sci. 2017, 18, 273. [Google Scholar] [CrossRef] [Green Version]
- Grosse, J.; Heffron, H.; Burling, K.; Hossain, M.A.; Abob, A.M.; Rogers, G.J.; Richards, P.; Larder, L.; Rimmington, D.; Adriaenssens, A.A.; et al. Insulin-like peptide-5 is an orexigenic gastrointestinal hormone. Proc. Natl. Acad. Sci. USA 2014, 111, 11133–11138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, E.L.; Hollingsworth, K.G.; Aribisale, B.S.; Chen, M.J.; Mothers, J.C.; Taylor, R. Reversal of type 2 diabetes: Normalization of beta cell function in association with decreased pancreas and liver triacylglycerol. Diabetologia 2011, 54, 2506–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ericson, U.C.; Sonestedt, E.; Gullberg, B.; Hellstrand, S.; Hindy, G.; Wirfalt, E.; Orho-Melander, M. High intake of protein and processed meat is associated with increased incidence of type 2 diabetes. Diabetologia 2011, 54 (Suppl. S1), S12. [Google Scholar]
- Ley, S.H.; Hamdy, O.; Mohan, V.; Hu, F.B. Prevention and management of type 2 diabetes: Dietary components and nutritional strategies. Lancet 2014, 383, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Antebi, A.; Bartke, A.; Barzilai, N.; Brown-Borg, H.M.; Caruso, C.; Curiel, T.J.; de Cabo, R.; Franceschi, C.; Gems, D.; et al. Interventions to Slow Aging in Humans: Are We Ready? Aging Cell 2015, 14, 497–510. [Google Scholar] [CrossRef]
- Buse, J.B.; Polonsky, K.S.; Burant, C.F. Type 2 diabetes mellitus. In Williams Textbook of Endocrinology, 11th ed.; Kronenberg, H.M., Melmed, S., Polonsky, K.S., Reed Larsen, P., Eds.; Saunders-Elsevier: Philadelphia, PA, USA, 2008; pp. 1329–1389. [Google Scholar]
- O’Connor, L.M.; Lntjes, M.A.H.; Luben, R.N. Dietary dairy product intake and incident type 2 diabetes: A prospective study using dietary data from a 7-day food diary. Diabetologia 2014, 57, 907–917. [Google Scholar] [CrossRef] [Green Version]
- Rigamonti, A.E.; Leoncini, R.; Casnici, C.; Marelli, O.; Col, A.; Tamini, S.; Lucchetti, E.; Cicolini, S.; Abbruzzese, L.; Cella, S.G.; et al. Whey proteins reduce appetite, stimulate anorexigenic gastrointestinal peptides and improve glucometabolic homeostasis in young obese women. Nutrients 2019, 11, 247. [Google Scholar] [CrossRef] [Green Version]
- Watson, L.E.; Phillips, L.K.; Wu, T.; Bound, M.J.; Checklin, H.; Grivell, J.; Jones, K.L.; Horowitz, M.; Rayner, C.K. Differentiating the effects of whey protein and guar gum preloads on postprandial glycemia in type 2 diabetes. Clin. Nutr. 2019, 38, 2827–2832. [Google Scholar] [CrossRef]
- Watson, L.E.; Phillips, L.K.; Wu, T.; Bound, M.J.; Checklin, H.L.; Grivell, J.; Jones, K.L.; Clifton, P.M.; Horowitz, M.; Rayner, C.K. A whey/guar “preload” improves postprandial glycaemia and glycated haemoglobin levels in type 2 diabetes: A 12-week, single-blind, randomized, placebo-controlled trial. Diabetes Obes. Metab. 2019, 21, 930–938. [Google Scholar] [CrossRef]
- Boscaini, S.; Cabrera-Rubio, R.; Nychyk, O.; Roger Speakman, J.; Francis Cryan, J.; David Cotter, P.; Nilaweera, K.N. Age- and duration-dependent effects of whey protein on high-fat diet-induced changes in body weight, lipid metabolism, and gut microbiota in mice. Physiol. Rep. 2020, 8, e14523. [Google Scholar] [CrossRef]
- Nonogaki, K.; Kaji, T. Whey protein isolate inhibits hepatic FGF21 production, which precedes weight gain, hyperinsulinemia and hyperglycemia in mice fed a high-fat diet. Sci. Rep. 2020, 10, 15784. [Google Scholar] [CrossRef]
- Fuglsang-Nielsen, R.; Rakvaag, E.; Langdahl, B.; Knudsen, K.E.B.; Hartmann, B.; Holst, J.J.; Hermansen, K.; Gregersen, S. Effects of whey protein and dietary fiber intake on insulin sensitivity, body composition, energy expenditure, blood pressure, and appetite in subjects with abdominal obesity. Eur. J. Clin. Nutr. 2020. Epub ahead of print. [Google Scholar] [CrossRef]
- Nair Sreekumaran, K.; Short, K.R. Hormonal and signaling role of branched-chain amino acids. J. Nutr. 2005, 135, 1547S–1551S. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 6, 311–326. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef]
- Xiao, F.; Huang, Z.; Li, H.; Yu, J.; Wang, C.; Chen, S.; Meng, Q.; Cheng, Y.; Gao, X.; Li, J.; et al. Leucine deprivation increases hepatic insulin sensitivity via GCN2/mTOR/S6K1 and AMPK pathways. Diabetes 2011, 60, 746–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremblay, F.; Marette, A. Amino acid and insulin signaling via the mTOR/p70 S6 kinase pathway. J. Biol. Chem. 2001, 276, 38052–38060. [Google Scholar]
- Tremblay, F.; Lavigne, C.; Jacques, H.; Marette, A. Role of dietary proteins and amino acids in the pathogenesis of insulin resistance. Ann. Rev. Nutr. 2007, 27, 293–310. [Google Scholar] [CrossRef]
- Qin, L.Q.; Xun, P.; Bujnowski, D.; Daviglus, M.L.; Van Horn, L.; Stamler, J.; He, K.; for the INTERMAP Cooperative Research Group. Higher branched-chain amino acid intake is associated with a lower prevalence of being overweight or obese in middle-aged East Asian and Western adults. J. Nutr. 2011, 141, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Solerte, S.B.; Fioravanti, M.; Locatelli, E.; Bonacasa, R.; Zamboni, M.; Basso, C.; Mazzoleni, A.; Mansi, V.; Geroutis, N.; Gazzaruso, C. Improvement of blood glucose control and insulin sensitivity during a long term (60 weeks) randomized study with amino acid dietary supplements in elderly subjects with type 2 diabetes mellitus. Am. J. Cardiol. 2008, 101, 82E–88E. [Google Scholar] [CrossRef]
- Macotela, Y.; Emanuelli, B.; Bang, A.M.; Espinoza, D.O.; Boucher, J.; Beebe, K.; Gall, W.; Kahn, C.R. Dietary Leucine—An environmental modifier of insulin resistance acting on multiple levels of metabolism. PLoS ONE 2011, 6, e2187. [Google Scholar] [CrossRef] [Green Version]
- Doi, M.; Yamaoka, I.; Fukunaga, T.; Nakayama, M. Isoleucine, a potent plasma glucose-lowering amino acid, stimulates glucose uptake in C2C12 myotubes. BioChem. Biophys. Res. Commun. 2003, 312, 1111–1117. [Google Scholar] [CrossRef]
- Kleinert, M.; Liao, Y.H.; Nelson, J.L.; Bernard, J.R.; Wang, W.; Ivy, J.L. An amino acid mixture enhances insulin-stimulated glucose uptake in isolated rat epitrochlearis muscle. J. Appl. Physiol. 2011, 111, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Valerio, A.; D’Antona, G.; Nisoli, E. Branched-chain amino acids, mitochondrial biogenesis and healthspan: An evolutionary perspective. Aging 2011, 3, 464–478. [Google Scholar] [CrossRef] [Green Version]
- Corrochano, A.R.; Ferraretto, A.; Arranz, E.; Stuknytė, M.; Bottani, M.; O’Connor, P.M.; Kelly, P.M.; De Noni, I.; Buckin, V.; Giblin, L. Bovine whey peptides transit the intestinal barrier to reduce oxidative stress in muscle cells. Food Chem. 2019, 288, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Jakubowicz, D.; Froy, O. Biochemical and metabolic mechanisms by which dietary whey protein may combat obesity and type 2 diabetes. J. Nutr. BioChem. 2013, 24, 1–5. [Google Scholar] [CrossRef]
- Jakubowicz, D.; Froy, O.; Ahren, B.; Boaz, M.; Landau, Z.; Bar-Dayan, Y.; Ganz, T.; Barnea, M.; Wainstein, J. Incretin, insulinotropic and glucose-lowering effects of whey protein pre-load in type 2 diabetes: A randomised clinical trial. Diabetologia 2014, 57, 1807–1811. [Google Scholar] [CrossRef]
- King, D.G.; Walker, M.; Campbell, M.D.; Breen, L.; Stevenson, E.J.; West, D.J. A small dose of whey protein co-ingested with mixed macronutrient breakfast and lunch meals improves postprandial glycemia and suppresses appetite in mem with type 2 diabetes: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 107, 550–557. [Google Scholar] [CrossRef]
- Patil, P.; Mandal, S.; Kumar Tomar, S.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 64, 863–880. [Google Scholar] [CrossRef]
- Sartorius, T.; Weidner, A.; Dharsono, T.; Boulier, A.; Wilhelm, M.; Schön, C. Postprandial Effects of a Proprietary Milk Protein Hydrolysate Containing Bioactive Peptides in Prediabetic Subjects. Nutrients 2019, 11, 1700, Erratum in: Nutrients 2020, 12, 1266. [Google Scholar] [CrossRef] [Green Version]
- Elovaris, R.A.; Hutchison, A.T.; Lange, K.; Horowitz, M.; Feinle-Bisset, C.; Luscombe-Marsh, N.D. Plasma Free Amino Acid Responses to Whey Protein and Their Relationships with Gastric Emptying, Blood Glucose- and Appetite-Regulatory Hormones and Energy Intake in Lean Healthy Men. Nutrients 2019, 11, 2465. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, M.; Holst, J.J.; Björck, I.M. Metabolic effects of amino acid mixtures and whey protein in healthy subjects: Studies using glucose-equivalent drinks. Am. J. Clin. Nutr. 2007, 85, 996–1004. [Google Scholar] [CrossRef] [Green Version]
- Butterworth, M.; Lees, M.; Harlow, P.; Hind, K.; Duckworth, L.; Ispoglou, T. Acute effects of essential amino acid gel-based and whey protein supplements on appetite and energy intake in older women. Appl. Physiol. Nutr. Metab. 2019, 44, 1141–1149. [Google Scholar] [CrossRef]
- Meisel, H. Biochemical properties of regulatory peptides derived from milk proteins. Biopolymers 1997, 43, 119–128. [Google Scholar] [CrossRef]
- Hernandez-Ledesma, B.; Recio, I.; Amigo, L. Beta-lactoglobulin as a source of bioactive peptides. Amino Acids 2008, 35, 257–265. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef] [Green Version]
- Lorenzini, E.C.; Chessa, S.; Chiatti, F.; Caroli, A.; Pagnacco, G. Peptidi bioattivi di latte e derivati. Scienza e Tecnica Lattiero-Casearia 2007, 58, 113–156. [Google Scholar]
- Rafferty, E.P.; Wylie, A.R.; Elliott, C.T.; Chevallier, O.P.; Grieve, D.J.; Green, B.D. In vitro and in vivo effects of natural putative secretagogues of glucagon-like peptide-1 (GLP-1). Sci. Pharm. 2011, 79, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Power-Grant, O.; Bruen, C.; Brennan, L.; Giblin, L.; Jakeman, P.; FitzGerald, R.J. In vitro bioactive properties of intact and enzymatically hydrolysed whey protein: Targeting the enteroinsular axis. Food Funct. 2015, 6, 972–980. [Google Scholar] [CrossRef]
- Gillespie, A.L.; Calderwood, D.; Hobson, L.; Green, B.D. Whey proteins have beneficial effects on intestinal enteroendocrine cells stimulating cell growth and increasing the production and secretion of incretin hormones. Food Chem. 2015, 189, 120–128. [Google Scholar] [CrossRef]
- Corrochano, A.R.; Arranz, E.; De Noni, I.; Stuknytė, M.; Ferraretto, A.; Kelly, P.M.; Buckin, V.; Giblin, L. Intestinal health benefits of bovine whey proteins after simulated gastrointestinal digestion. J. Funct. Foods 2018, 49, 526–535. [Google Scholar] [CrossRef]
- Gillespie, A.L.; Green, B.D. The bioactive effects of casein proteins on enteroendocrine cell health, proliferation and incretin hormone secretion. Food Chem. 2016, 211, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Moya, T.; Planes-Muñoz, D.; Frontela-Saseta, C.; Ros-Berruezo, G.; López-Nicolás, R. Milk whey from different animal species stimulates the in vitro release of CCK and GLP-1 through a whole simulated intestinal digestion. Food Funct. 2020, 11, 7208–7216. [Google Scholar] [CrossRef]
- Ripken, D.; van der Wielen, N.; Wortelboer, H.M.; Meijerink, J.; Witkamp, R.F.; Hendriks, H.F. Nutrient-induced glucagon like peptide-1 release is modulated by serotonin. J. Nutr. BioChem. 2016, 32, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Kondrashina, A.; Bruen, C.; McGrath, B.; Murray, B.; McCarthy, T.; Schellekens, H.; Buzoianu, S.; Cryan, J.F.; Kelly, A.L.; McSweeney, P.L.H.; et al. Satiating effect of a sodium caseinate hydrolysate and its fate in the upper gastrointestinal tract. J. Funct. Foods 2018, 49, 306–313. [Google Scholar] [CrossRef]
- CordierBussat, M.; Bernard, C.; Haouche, S.; Roche, C.; Abello, J.; Chayvialle, J.A.; Cuber, J.C. Peptones stimulate cholecystokinin secretion and gene transcription in the intestinal cell line STC-1. Endocrinology 1997, 138, 1137–1144. [Google Scholar] [CrossRef]
- Hira, T.; Maekawa, T.; Asano, K.; Hara, H. Cholecystokinin secretion induced by β-conglycinin peptone depends on Gαq-mediated pathways in enteroendocrine cells. Eur. J. Nutr. 2009, 48, 124–127. [Google Scholar] [CrossRef]
- Perazza, L.R.; Daniel, N.; Dubois, M.J.; Pilon, G.; Varin, T.V.; Blais, M.; Martinez Gonzales, J.L.; Bouchard, M.; Asselin, C.; Lessard, M.; et al. Distinct Effects of Milk-Derived and Fermented Dairy Protein on Gut Microbiota and Cardiometabolic Markers in Diet-Induced Obese Mice. J. Nutr. 2020. Epub ahead of print. [Google Scholar] [CrossRef]
- Schellekens, H.; Nongonierma, A.B.; Clarke, G.; van Oeffelen, W.E.; FitzGerald, R.J.; Dinan, T.G.; Cryan, J.F. Milk protein-derived peptides induce 5-HT2C mediated satiety in vivo. Int. Dairy J. 2014, 38, 55–64. [Google Scholar] [CrossRef] [Green Version]
- O’Halloran, F.; Bruen, C.; McGrath, B.; Schellekens, H.; Murray, B.; Cryan, J.F.; Kelly, A.L.; McSweeney, P.L.H.; Giblin, L. A casein hydrolysate increases GLP-1 secretion and reduces food intake. Food Chem. 2018, 252, 303–310. [Google Scholar] [CrossRef]
- Osborne, S.; Chen, W.; Addepalli, R.; Colgrave, M.; Singh, T.; Tran, C.; Day, L. In vitro transport and satiety of a beta-lactoglobulin dipeptide and beta-casomorphin-7 and its metabolites. Food Funct. 2014, 5, 2706–2718. [Google Scholar] [CrossRef] [Green Version]
- Backus, R.C.; Howard, K.A.; Rogers, Q.R. The potency of dietary amino acids in elevating plasma cholecystokinin immunoreactivity in cats is related to amino acid hydrophobicity. Regul. Pept. 1997, 72, 31–40. [Google Scholar] [CrossRef]
- Pupovac, J.; Anderson, G.H. Dietary peptides induce satiety via cholecystokinin-A and peripheral opioid receptors in rats. J. Nutr. 2002, 132, 2775–2780. [Google Scholar] [CrossRef] [Green Version]
- Zong, Y.-F.; Chen, W.-H.; Zhang, Y.-S.; Zou, S.-X. Effects of intra-gastric beta-casomorphin-7 on somatostatin and gastrin gene expression in rat gastric mucosa. World J. Gastroenterol. 2007, 13, 2094. [Google Scholar] [CrossRef]
- Günther, T.; Tulipano, G.; Dournaud, P.; Bousquet, C.; Csaba, Z.; Kreienkamp, H.J.; Lupp, A.; Korbonits, M.; Castaño, J.P.; Wester, H.J.; et al. International Union of Basic and Clinical Pharmacology. CV. Somatostatin Receptors: Structure, Function, Ligands, and New Nomenclature. Pharmacol. Rev. 2018, 70, 763–835. [Google Scholar] [CrossRef] [Green Version]
- Boutrou, R.; Henry, G.; Sanchez-Rivera, L. On the trail of milk bioactive peptides in human and animal intestinal tracts during digestion: A review. Dairy Sci. Technol. 2015, 95, 815–829. [Google Scholar] [CrossRef] [Green Version]
- Aslam, H.; Ruusunen, A.; Berk, M.; Loughman, A.; Rivera, L.; Pasco, J.A.; Jacka, F.N. Unravelled facets of milk derived opioid peptides: A focus on gut physiology, fractures and obesity. Int. J. Food Sci. Nutr. 2020, 71, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Foisy Sauvé, M.; Spahis, S.; Delvin, E.; Levy, E. Glycomacropeptide: A Bioactive Milk Derivative to Alleviate Metabolic Syndrome Outcomes. Antioxid. Redox Signal. 2020. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Beucher, S.; Levenez, F.; Yvon, M.; Corring, T. Effects of gastric digestive products from casein on CCK release by intestinal cells in rat. J. Nutr. BioChem. 1994, 5, 578–584. [Google Scholar] [CrossRef]
- Chen, Q.; Cao, J.; Jia, Y.; Liu, X.; Yan, Y.; Pang, G. Modulation of mice fecal microbiota by administration of casein glycomacropeptide. MicroBiol. Res. 2012, 3, e3. [Google Scholar] [CrossRef]
- Royle, P.J.; McIntosh, G.H.; Clifton, P.M. Whey protein isolate and glycomacropeptide decrease weight gain and alter body composition in male Wistar rats. Br. J. Nutr. 2008, 100, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Burton-Freeman, B.M. Glycomacropeptide (GMP) is not critical to whey-induced satiety, but may have a unique role in energy intake regulation through cholecystokinin (CCK). Physiol. Behav. 2008, 93, 379–387. [Google Scholar] [CrossRef]
- Veldhorst, M.A.B.; Nieuwenhuizen, A.G.; Hochstenbach-Waelen, A.; Westerterp, K.R.; Engelen, M.P.K.J.; Brummer, R.J.M.; Deutz, N.E.; Westerterp-Plantenga, M.S. Effects of complete whey-protein breakfasts versus whey without GMP-breakfasts on energy intake and satiety. Appetite 2009, 52, 388–395. [Google Scholar] [CrossRef]
- Clifton, P.M.; Keogh, J.; Woonton, B.; Taylor, C.; Janakievski, F.; De Silva, K. Effect of glycomacropeptides (GMP) on satiety hormones and food intake. Aust. J. Dairy Technol. 2009, 64, 29. [Google Scholar]
- Keogh, J.B.; Woonton, B.W.; Taylor, C.M.; Janakievski, F.; Desilva, K.; Clifton, P.M. Effect of glycomacropeptide fractions on cholecystokinin and food intake. Br. J. Nutr. 2010, 104, 286–290. [Google Scholar] [CrossRef] [Green Version]
- Wernlund, P.G.; Hvas, C.L.; Dahlerup, J.F.; Bahl, M.I.; Licht, T.R.; Knudsen, K.E.; Agnholt, J.S. Casein glycomacropeptide is well tolerated in healthy adults and changes neither high-sensitive C-reactive protein, gut microbiota nor faecal butyrate: A restricted randomised trial. Br. J. Nutr. 2020, 24, 1–35. [Google Scholar] [CrossRef]
- Aslam, H.; Marx, W.; Rocks, T.; Loughman, A.; Chandrasekaran, V.; Ruusunen, A.; Dawson, S.L.; West, M.; Mullarkey, E.; Pasco, J.A.; et al. The effects of dairy and dairy derivatives on the gut microbiota: A systematic literature review. Gut Microbes 2020, 12, 1799533. [Google Scholar] [CrossRef] [PubMed]
- Kagebayashi, T.; Kontani, N.; Yamada, Y.; Mizushige, T.; Arai, T.; Kino, K.; Ohinata, K. Novel CCK-dependent vasorelaxing dipeptide, Arg-Phe, decreases blood pressure and food intake in rodents. Mol. Nutr. Food Res. 2012, 56, 1456–1463. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Le Maux, S.; Hamayon, J.; FitzGerald, R.J. Strategies for the release of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides in an enzymatic hydrolyzate of α-lactalbumin. Food Funct. 2016, 7, 3437–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Cheng, J.; Wu, H. Discovery of food-derive dipeptidyl peptidase IV inhibitory peptides: A review. Int. J. Mol. Sci. 2019, 20, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, K.; Wu, J. Purification and characterisation of angiotensin converting enzyme (ACE) inhibitory peptides derived from enzymatic hydrolysate of ovotransferrin. Food Chem. 2011, 126, 1614–1619. [Google Scholar] [CrossRef]
- Horner, K.; Drummond, E.; Brennan, L. Bioavailability of milk protein-derived bioactive peptides: A glycaemic management perspective. Nutr. Res. Rev. 2016, 29, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacroix, I.M.E.; Chen, X.M.; Kitts, D.D.; Li-Chan, E.C.Y. Investigation into the bioavailability of milk protein-derived peptides with dipeptidyl-peptidase IV inhibitory activity using Caco-2 cell monolayers. Food Funct. 2017, 8, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.D.; Beverly, R.L.; Underwood, M.A.; Dallas, D.C. Release of functional peptides from mother’s milk and fortifier proteins in the premature infant stomach. PLoS ONE 2018, 13, e0208204. [Google Scholar] [CrossRef]
- Silveira, S.T.; Martínez-Maqueda, D.; Recio, I.; Hernández-Ledesma, B. Dipeptidyl peptidase-IV inhibitory peptides generated by tryptic hydrolysis of a whey protein concentrate rich in β-lactoglobulin. Food Chem. 2013, 141, 1072–1077. [Google Scholar] [CrossRef]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Fitzgerald, R.J. Inhibition of dipeptidyl peptidase IV (DPP-IV) by tryptophan containing dipeptides. Food Funct. 2013, 4, 1843–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, Y.; Hara, H.; Hira, T. Glucagon-like peptide-1 response to whey protein is less diminished by dipeptidyl peptidase-4 in comparison with responses to dextrin, a lipid and casein in rats. Br. J. Nutr. 2020, 1, 10, Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Yin, Y.; Zhao, W.; Yu, Y.; Liu, B.; Liu, J.; Chen, F. Novel peptides derived from egg white protein inhibiting alpha-glucosidase. Food Chem. 2011, 129, 1376–1382. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C.Y. Inhibition of dipeptidyl peptidase (DPP)-IV and α-glucosidase activities by pepsin treated whey proteins. J. Agric. Food Chem. 2013, 61, 7500–7506. [Google Scholar] [CrossRef]
- Konrad, B.; Anna, D.; Marek, S.; Marta, P.; Aleksandra, Z.; Jozefa, C. The evaluation of dipeptidyl peptidase (DPP)-IV, α-Glucosidase and angiotensin converting enzyme (ACE) inhibitory activities of whey proteins hydrolyzed with serine protease isolated from asian pumpkin (Cucurbita ficifolia). Int. J. Pept Res. Ther. 2014, 20, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Aoki, H.; Nakato, J.; Mizushige, T.; Iwakura, H.; Sato, M.; Suzuki, H.; Kanamoto, R.; Ohinata, K. Lacto-ghrestatin, a novel bovine milk-derived peptide, suppresses ghrelin secretion. FEBS Lett. 2017, 591, 2121–2130. [Google Scholar] [CrossRef] [Green Version]
- Hou, I.-C.; Yoshikawa, M.; Ohinata, K. β-Lactotensin derived from bovine β-lactoglobulin suppresses food intake via the CRF system followed by the CGRP system in mice. Peptides 2009, 30, 2228–2232. [Google Scholar] [CrossRef]
- Ogiwara, M.; Ota, W.; Mizushige, T.; Kanamoto, R.; Ohinata, K. Enzymatic digest of whey protein and wheylin-1, a dipeptide released in the digest, increase insulin sensitivity in an Akt phosphorylation-dependent manner. Food Funct. 2018, 9, 4635–4641. [Google Scholar] [CrossRef]
- Prasad, C.; Imrhan, V.; Juma, S.; Maziarz, M.; Prasad, A.; Tiernan, C.; Vijayagopal, P. Bioactive Plant Metabolites in the Management of Non-Communicable Metabolic Diseases: Looking at Opportunities beyond the Horizon. Metabolites 2015, 5, 733–765. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Hayashi, M.; Murakami, Y.; Araki, Y.; Otsuka, Y.; Kashiwagi, T.; Shimamura, T.; Ukeda, H. Development of LC-MS/MS analysis of cyclic dipeptides and its application to tea extract. BioSci. Biotechnol. BioChem. 2016, 80, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Nagai, A.; Mizushige, T.; Matsumura, S.; Inoue, K.; Ohinata, K. Orally administered milk-derived tripeptide improved cognitive decline in mice fed a high-fat diet. FASEB J. 2019, 33, 14095–14102. [Google Scholar] [CrossRef] [Green Version]
- Howick, K.; Wallace-Fitzsimons, S.E.; Kandil, D.; Chruścicka, B.; Calis, M.; Murphy, E.; Murray, B.A.; Fernandez, A.; Barry, K.M.; Kelly, P.M.; et al. A Dairy-Derived Ghrelinergic Hydrolysate Modulates Food Intake In Vivo. Int. J. Mol. Sci. 2018, 19, 2780. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.S.; Adama, E.; Clayton, K.; Holbrey, J.; Palubiski, G.; Smith, H.A.; Gonzalez, J.T.; Betts, J.A. Nocturnal whey protein ingestion impairs post-prandial glucose tolerance at breakfast. Br. J. Nutr. 2020, 1, 9, Epub ahead of print. [Google Scholar] [CrossRef]
- Allerton, D.M.; Rumbold, P.L.S.; West, D.J.; Stevenson, E.J. Effects of supplemental whey protein timing on postprandial glycaemia in centrally obese males. Br. J. Nutr. 2018, 121, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Akbar, N.; Azzimato, V.; Choudhury, R.P.; Aouadi, M. Extracellular vesicles in metabolic disease. Diabetologia 2019, 62, 2179–2187. [Google Scholar] [CrossRef] [Green Version]
- Brooks Mobley, C.; Mumford, P.W.; McCarthy, J.J.; Miller, M.E.; Young, K.C.; Martin, J.S.; Beck, D.T.; Lockwood, C.M.; Roberts, M.D. Whey protein-derived exosomes increase protein synthesis and hypertrophy in C2 C12 myotubes. J. Dairy Sci. 2016, 100, 48–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiferman, A.; Shu, J.; Grove, R.; Cui, J.; Adamec, J.; Zempleni, J. A diet defined by its content of bovine milk exosomes and their RNA cargos has moderate effects on gene expression, amino acid profiles and grip strength in skeletal muscle in C57BL/6 mice. J. Nutr. BioChem. 2018, 59, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Cundiff, J.K.; McConnell, E.J.; Lohe, K.J.; Maria, S.D.; McMahon, R.J.; Zhang, Q. Sensing Small Changes in Protein Abundance: Stimulation of Caco-2 Cells by Human Whey Proteins. J. Proteome Res. 2016, 15, 125–143. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Source | Peptides | Actions | References |
|---|---|---|---|
| β-lactoglobulin | ALPMH | -stimulates CCK secretion in vitro | [30] |
| β-casein (A1 genetic variant) | YPFPGPI YPFPGPI FPGPI | -opiod receptor agonist, -delays gut transit, -stimulates somatostatin release in vivo, -other μ-opiod receptor mediated effects in the gut, -up-regulation of DPP-IV espression in the small intestine (non opiod effect), -stimulate CCK secretion in vitro | [109,110] [113] [53] [8,107] |
| k-casein Whey protein fraction | Glycomacropeptide (GMP) | -stimulates CCK secretion from isolated rat intestine, -stimulates the release of satiety enteric hormones in mice, by altering the gut microbiota composition, -contradictory results as to the effects on appetite, food intake and enteric hormone release in humans, | [115] [116] [118,119,120,121,122] |
| Caseins Whey proteins | DPP-IV inhibitory peptides LKPTPEGDL, LPYPY, IPIQY, IPI, WR LPQNIPPL | -inhibit DPP-IV protease activity in vitro -possible delay of incretin hormone (GLP-1 and GIP) degradation and improvement of the control of postprandial glucose levels (systemic effect), -possible local action as inhibitors of DPP-IV activity in the gut (brush border of intestinal mucosa and gut microbiota) No clear evidence of their passage across the intestinal epithelial barrier into circulation -DPP-IV inhibitors in vitro, can cross an epithelial cell monolayer (Caco-2 cells) in vitro -DPP-IV inhibitor in vitro, improves postprandial glucose control in rats | [49,50,51,52,125,126] [134] [126,129] [8,132] |
| β-lactoglobulin Whey protein isolate | α-glucosidase inhibitory peptides AP and other peptides (not isolated and identified) | -inhibit α-glucosidase activity in vitro -inhibit α-glucosidase activity in vitro, -delay glucose absorption after meal in prediabetic subjects | [85,136,137] [86] |
| β-lactoglobulin | LIVTQTMKG | -decreases acylated ghrelin release in vitro, -decreases plasma ghrelin levels and food intake in fasted mice after oral administration | [138] |
| β-lactoglobulin | HIRL | -reduces food intake in fasted mice after oral-, ic- or ip administration | [139] |
| β-lactoglobulin | MH | -anxiolytic-like dipeptide, -increases insulin sensitivity in mice and Akt phosphorylation (insulin receptor signalling) in hepatocytes and skeletal muscle cells in vitro and in vivo | [140] |
| α-S1-casein | YLG | -reverses cognitive decline in mice fed a high-fat diet after oral administration (possibly direct effects on neuronal cells or indirect effects mediated by peripheral actions, including actions on metabolic health) | [143] |
| caseins | Ghrelinergic peptide mixtures | -activate ghrelin receptor (GHSR1a) in vitro -induce a short-term increase of food intake in rats after oral administration (not so after ip administration) | [144] |
| sodium caseinate | RF | -stimulates CCK secretion in vitro, -delays gut transit of food after oral administration and decreases food intake after ip administration in mice | [8,106,124] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tulipano, G. Role of Bioactive Peptide Sequences in the Potential Impact of Dairy Protein Intake on Metabolic Health. Int. J. Mol. Sci. 2020, 21, 8881. https://doi.org/10.3390/ijms21228881
Tulipano G. Role of Bioactive Peptide Sequences in the Potential Impact of Dairy Protein Intake on Metabolic Health. International Journal of Molecular Sciences. 2020; 21(22):8881. https://doi.org/10.3390/ijms21228881
Chicago/Turabian StyleTulipano, Giovanni. 2020. "Role of Bioactive Peptide Sequences in the Potential Impact of Dairy Protein Intake on Metabolic Health" International Journal of Molecular Sciences 21, no. 22: 8881. https://doi.org/10.3390/ijms21228881
APA StyleTulipano, G. (2020). Role of Bioactive Peptide Sequences in the Potential Impact of Dairy Protein Intake on Metabolic Health. International Journal of Molecular Sciences, 21(22), 8881. https://doi.org/10.3390/ijms21228881