Comparative Analysis of ROS Network Genes in Extremophile Eukaryotes


Abstract
:1. Introduction
2. Results
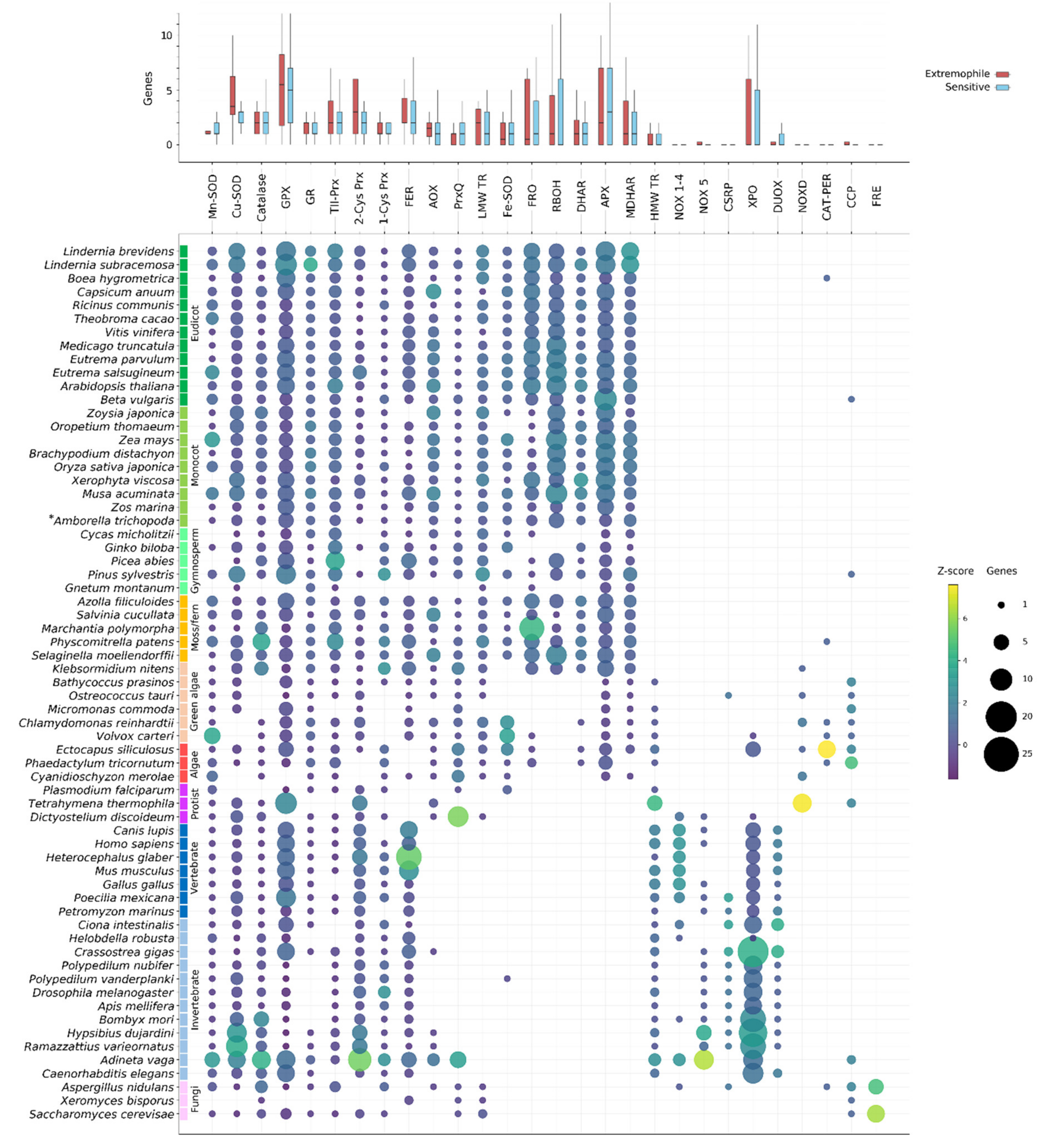
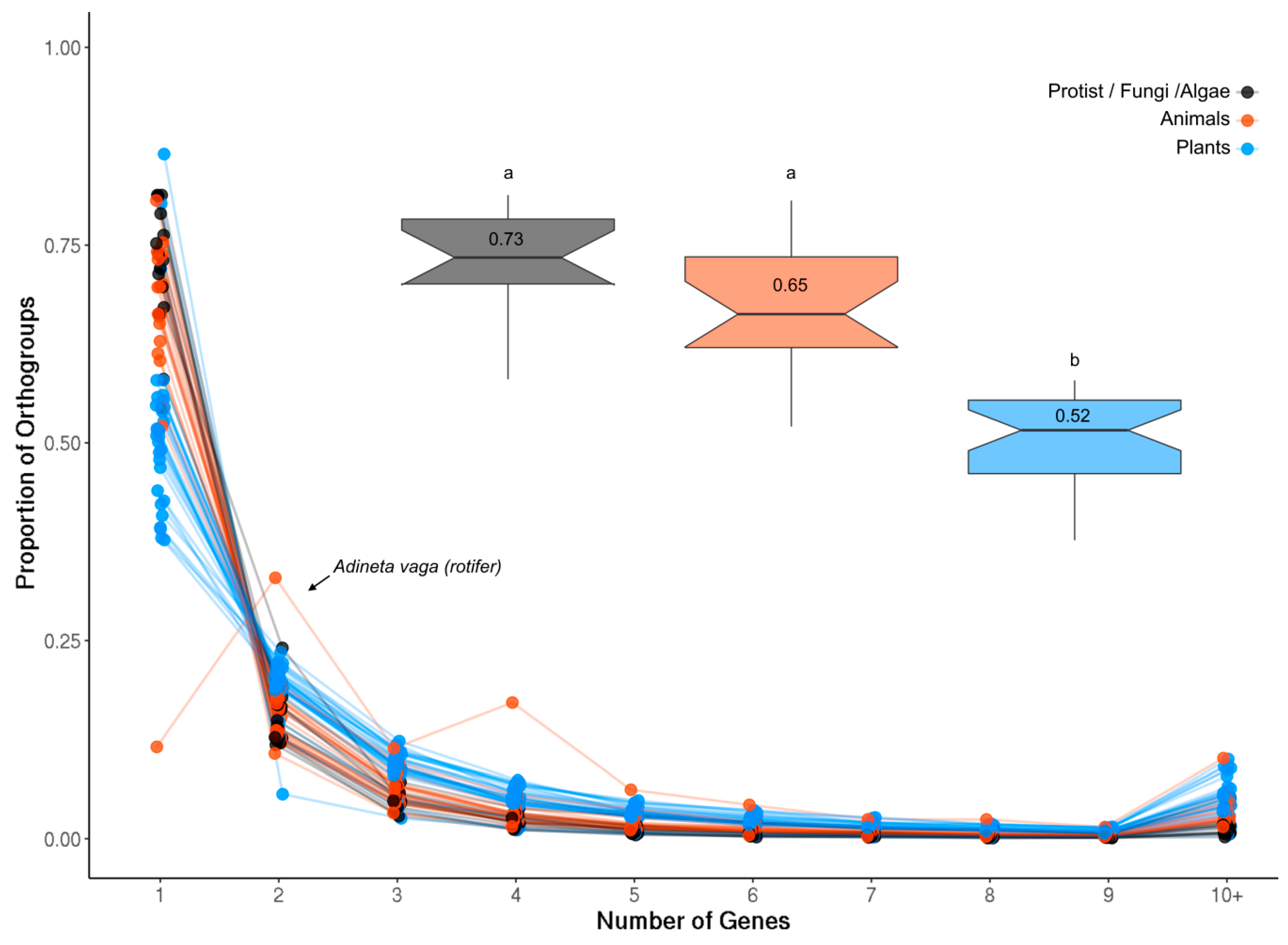
2.1. Evolution of the ROS Network Genes across Eukaryotes
2.2. Superoxide Dismutases
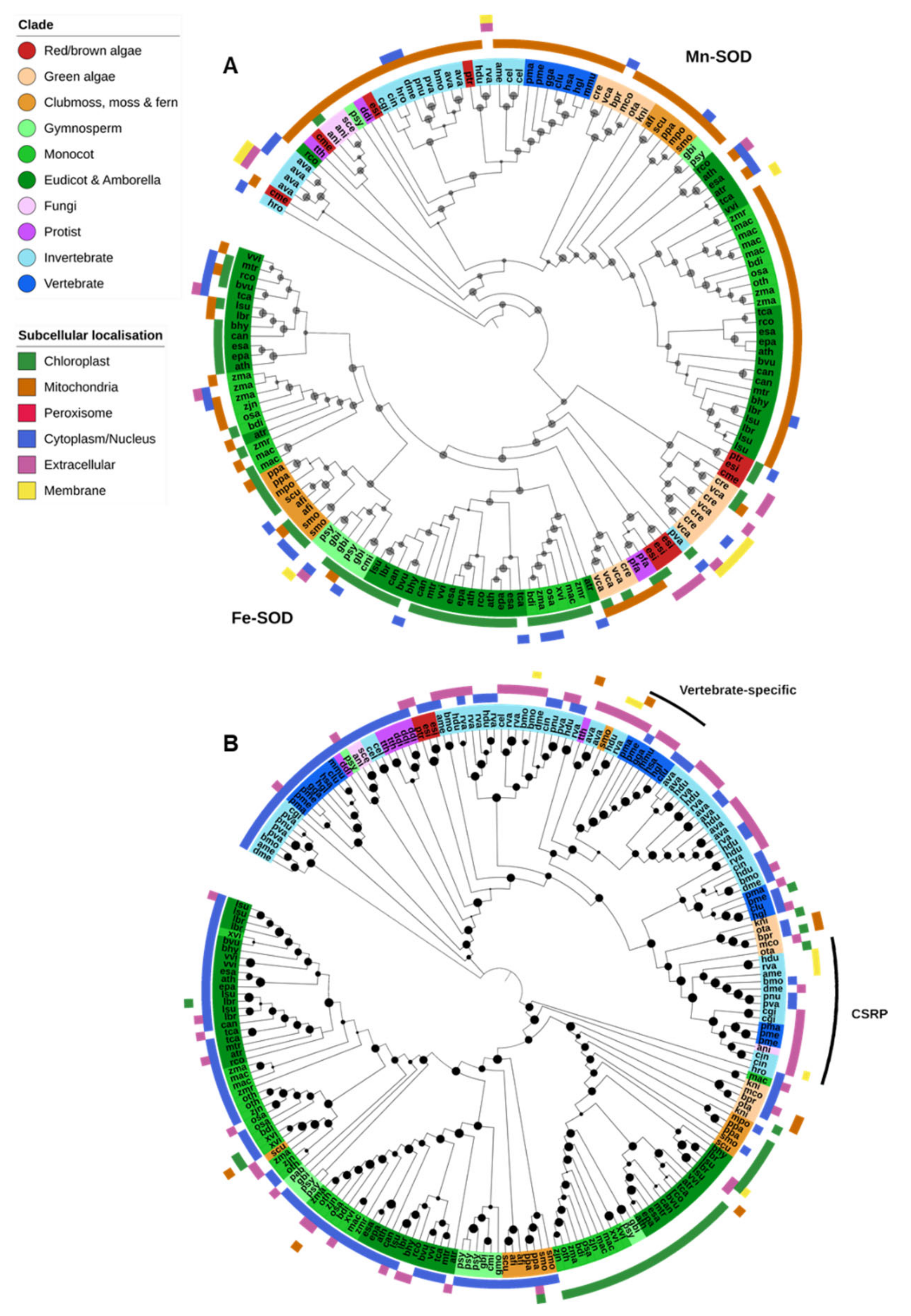
2.2.1. Fe/Mn-SOD
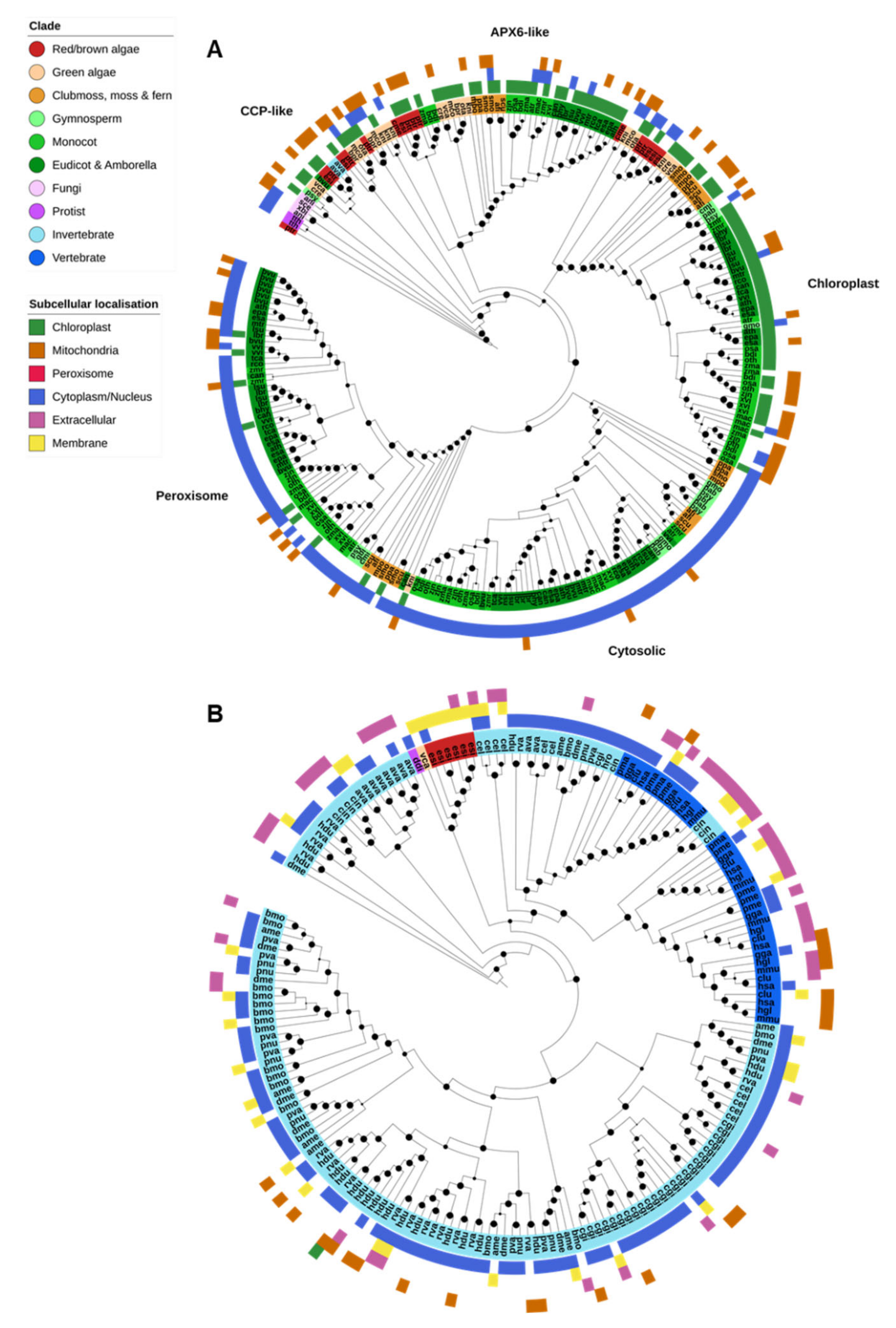
2.2.2. Cu/Zn-SOD & Ni-SOD
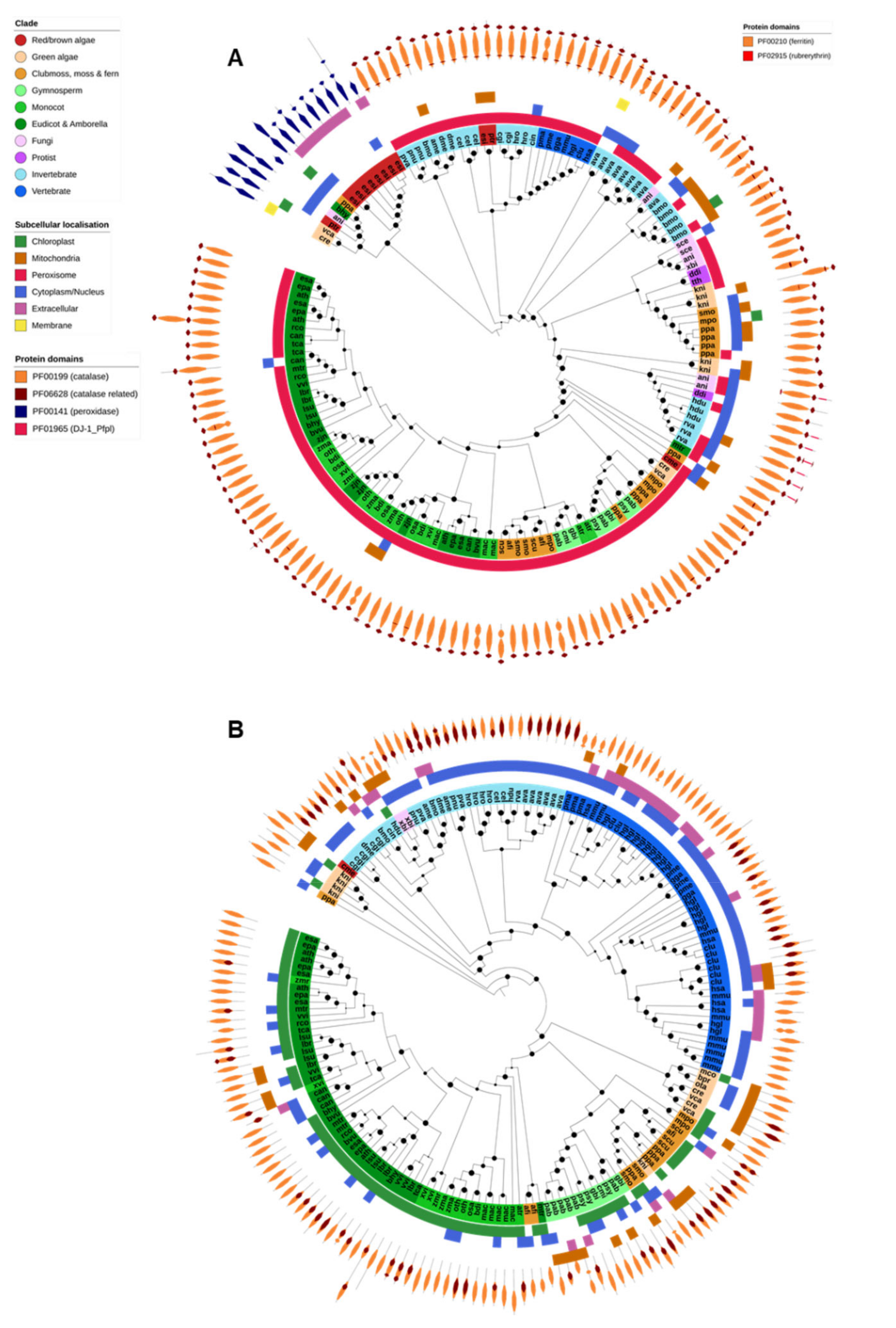
2.3. Catalases
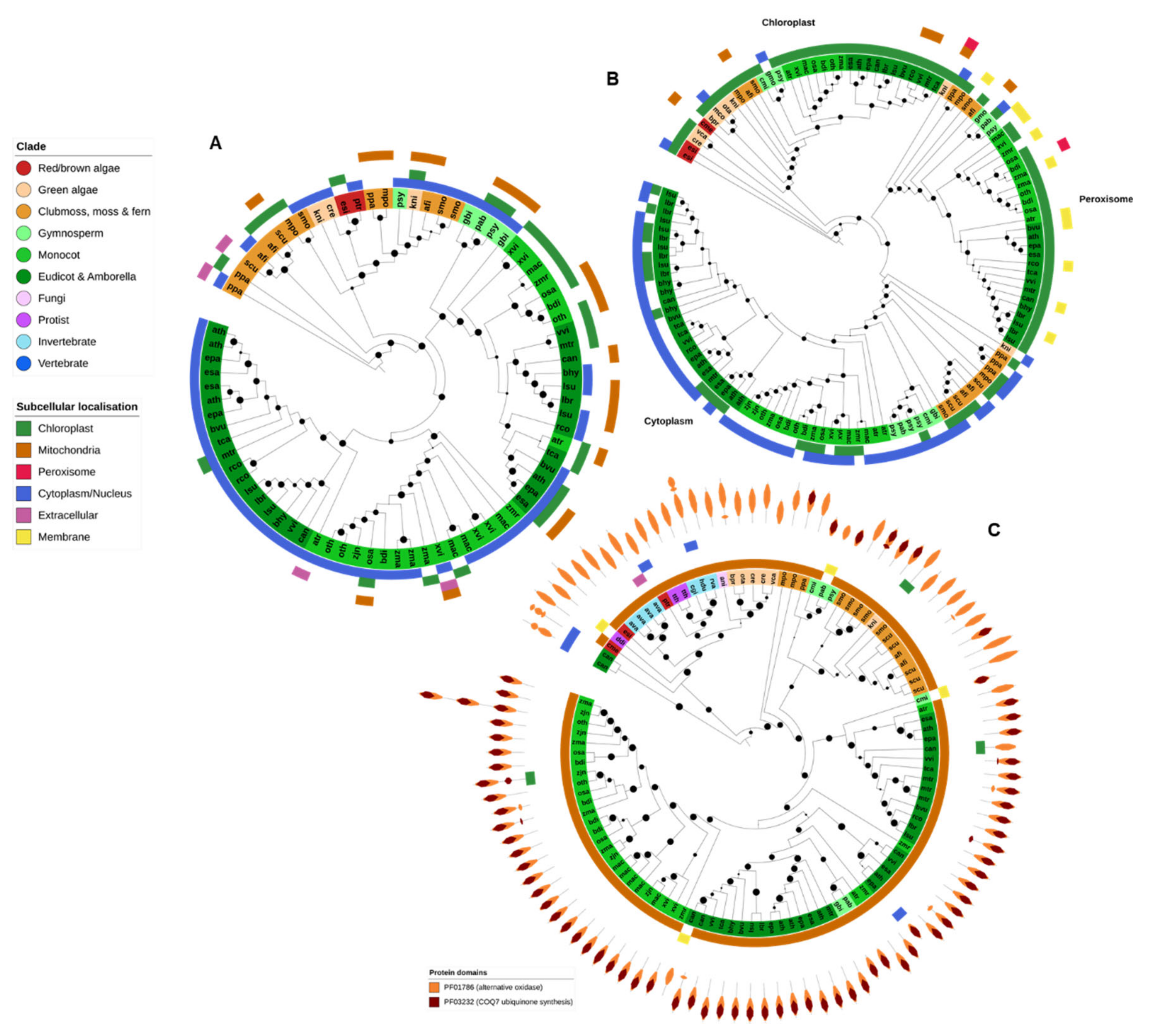
2.4. Ferritin
2.5. Heme Peroxidases
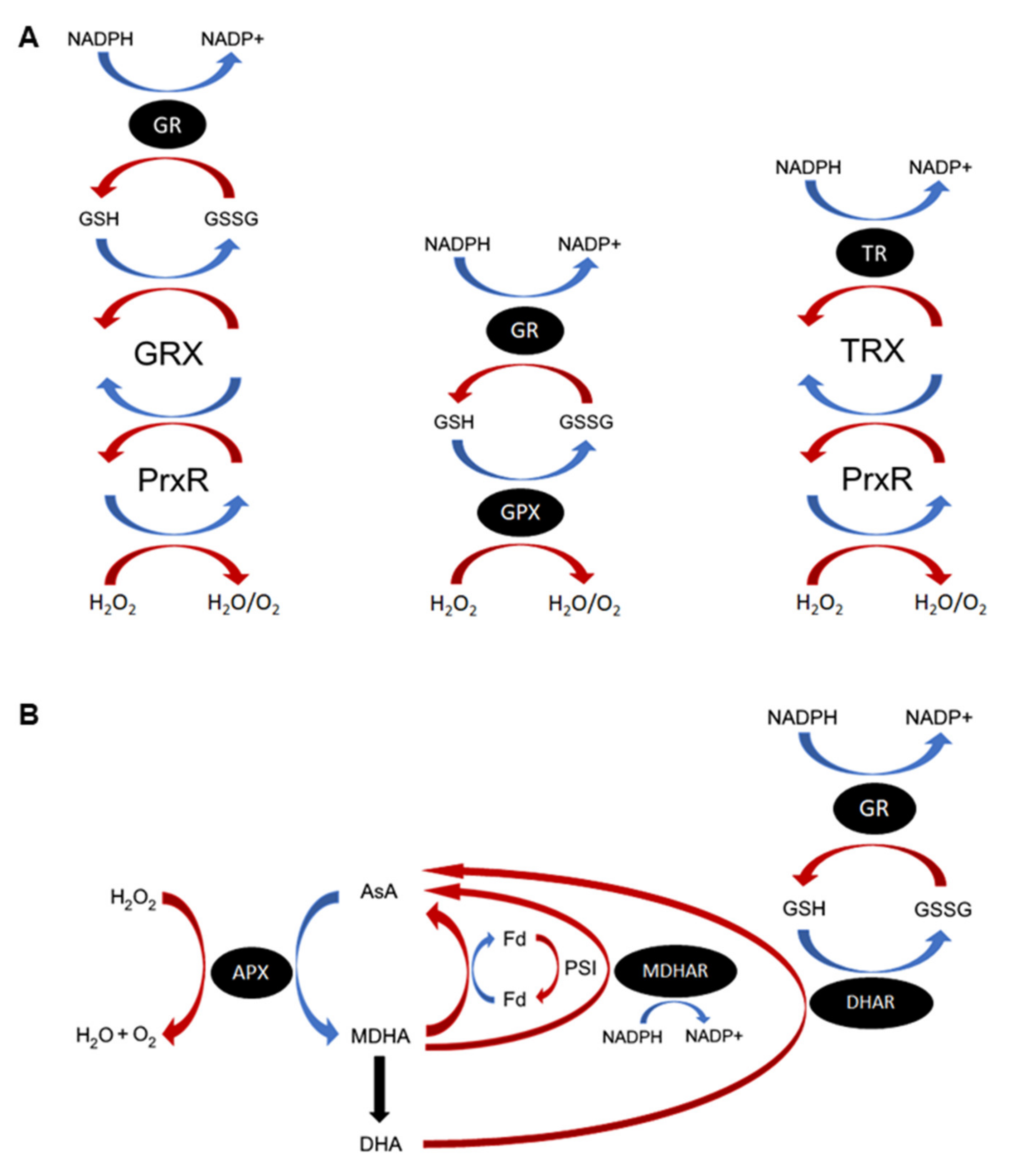
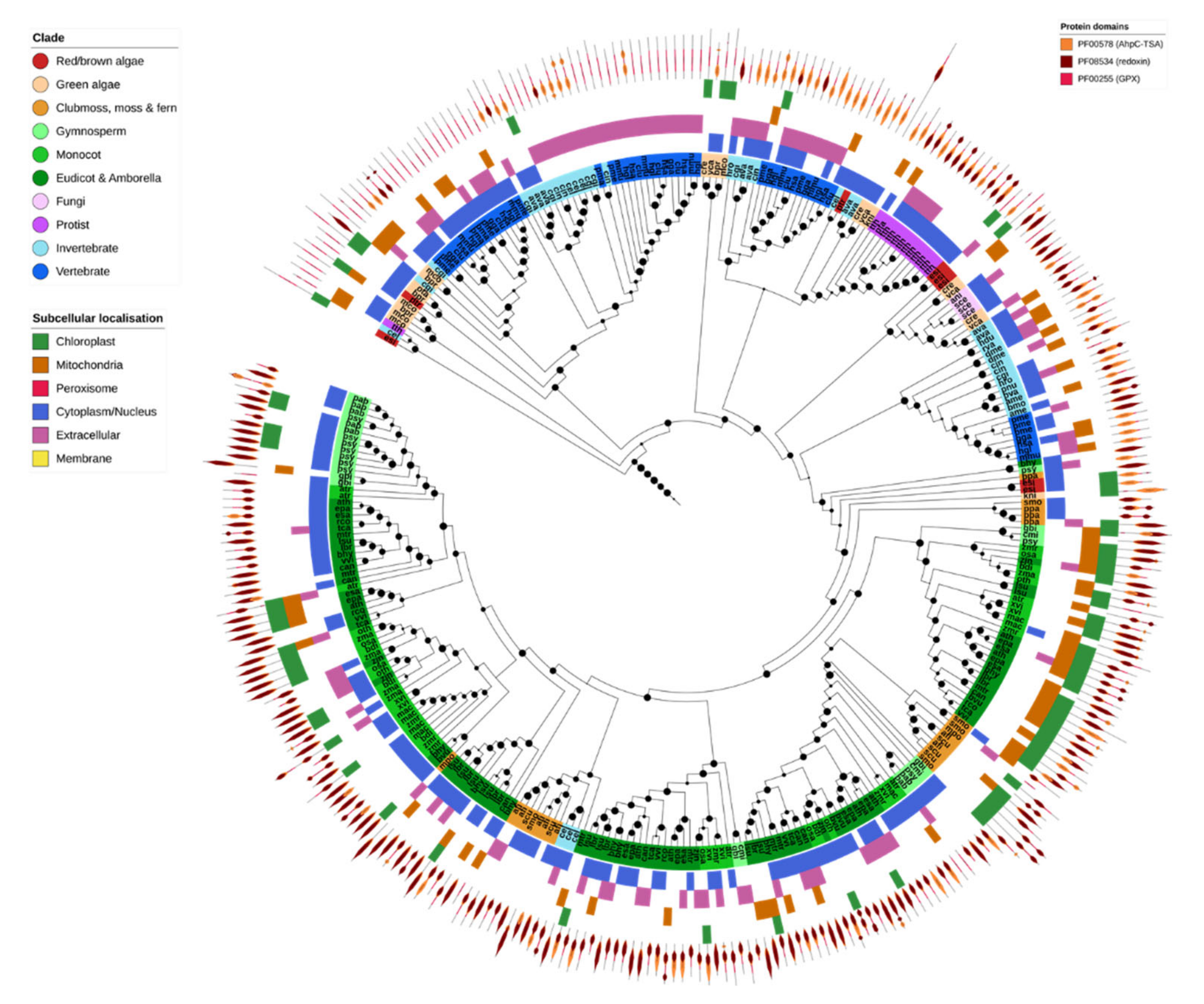
2.6. Non-Heme Peroxidases (Prx & GPX)
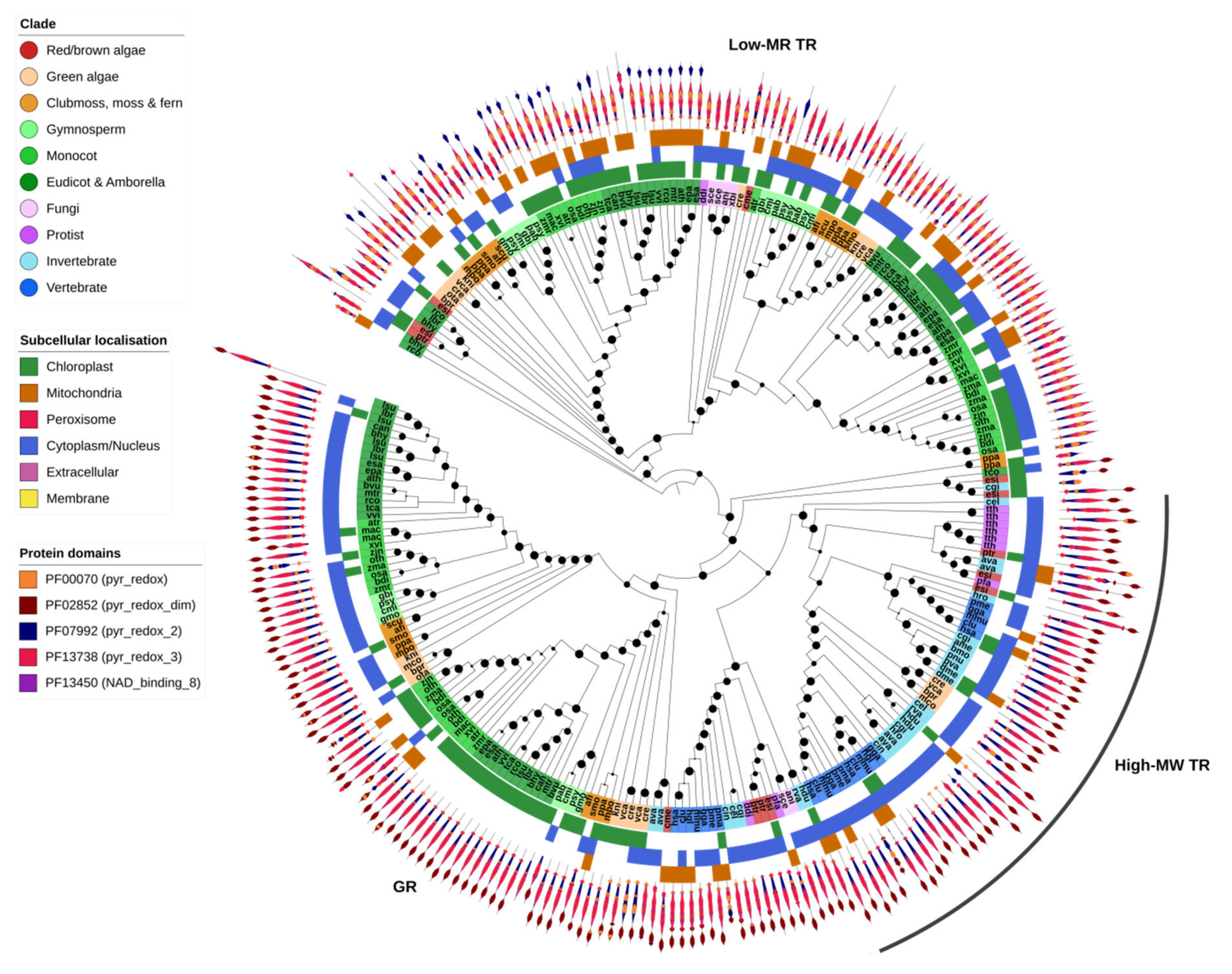
2.7. GSH/TRX Reductases
2.8. DHAR
2.9. MDHAR
2.10. AOX
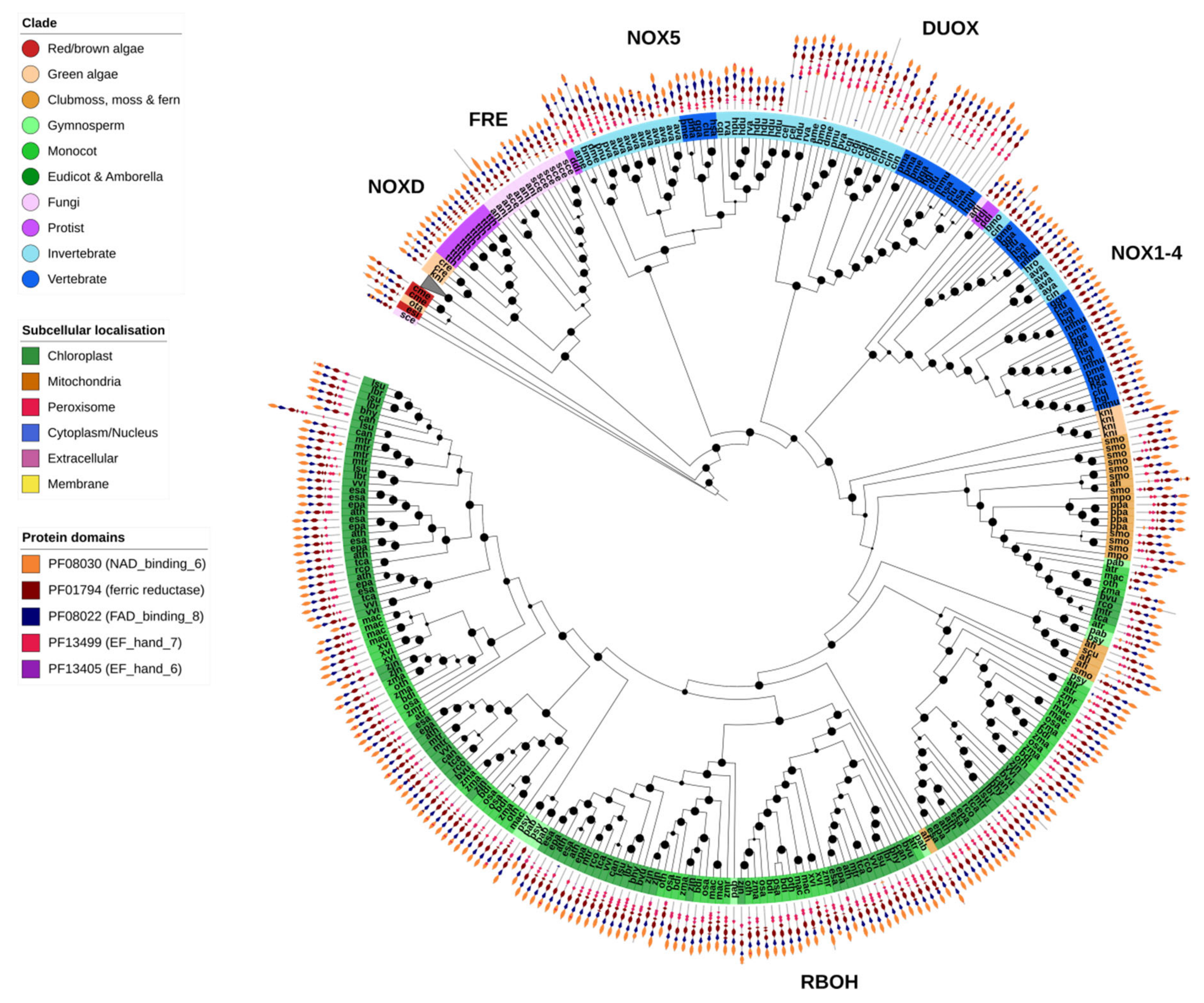
2.11. NOX-like
3. Discussion
4. Materials and Methods
4.1. Genomic Data
4.2. Orthogroup (OG) Analysis
5. Conclusion
Author Contributions
Funding
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Kingdom | Species | Description | Tolerance |
|---|---|---|---|---|
| vca | Algae (Green algae) | Volvox carteri | Green algae representative | |
| cre | Algae (Green algae) | Chlamydomonas reinhardtii | Green algae representative | |
| mco | Algae (Green algae) | Micromonas commoda | Green algae representative | |
| bpr | Algae (Green algae) | Bathycoccus prasinos | Green algae representative | |
| ota | Algae (Green algae) | Ostreococcus tauri | Green algae representative | |
| kni | Algae (Green algae) | Klebsormidium nitens | Green algae representative | |
| ppa | Plant (Moss) | Physcomitrella patens | Model DT moss | Desiccation |
| mpo | Plant (Liverwort) | Marchantia polymorpha | Basal to vascular plants | Desiccation |
| smo | Plant (Clubmoss) | Selaginella moellendorffii | Basal vascular plant | |
| scu | Plant (Fern) | Salvinia cucullata | Fern representative | |
| afi | Plant (Fern) | Azolla filiculoides | Fern representative | |
| gmo | Plant (Gymnosperm) | Gnetum montanum | Gymnosperm representative | |
| psy | Plant (Gymnosperm) | Pinus sylvestris | Gymnosperm representative | |
| pab | Plant (Gymnosperm) | Picea abies | Gymnosperm representative | |
| gbi | Plant (Gymnosperm) | Ginko biloba | Gymnosperm representative | |
| cmi | Plant (Gymnosperm) | Cycas micholitzii | Gymnosperm representative | |
| atr | Plant (Eudicot) | Amborella trichopoda | Basal angiosperm | |
| bvu | Plant (Eudicot) | Beta vulgaris | Crop species | |
| ath | Plant (Eudicot) | Arabidopsis thaliana | Model eudicot species | |
| epa | Plant (Eudicot) | Eutrema parvulum | Sister species to Eutrema salsugineum | |
| esa | Plant (Eudicot) | Eutrema salsugineum | Salt-tolerant dicot related to Arabidopsis thaliana | Salt |
| mtr | Plant (Eudicot) | Medicago truncatula | Model legume species | |
| vvi | Plant (Eudicot) | Vitis vinifera | Crop species | |
| rco | Plant (Eudicot) | Ricinus communis | Crop species | |
| tca | Plant (Eudicot) | Theobroma cacao | Crop species | |
| cca | Plant (Eudicot) | Capsicum anuum | Crop species | |
| bhy | Plant (Eudicot) | Boea hygrometrica | Resurrection plant | Desiccation |
| lsu | Plant (Eudicot) | Lindernia subracemosa | Sensitive sister species to Lindernia brevidens | |
| lbr | Plant (Eudicot) | Lindernia brevidens | Resurrection plant | Desiccation |
| zmr | Plant (Monocot) | Zos marina | Marine monocot | Salt |
| xvi | Plant (Monocot) | Xerophyta viscosa | Resurrection plant | Desiccation |
| mac | Plant (Monocot) | Musa acuminata | Crop species | |
| osa | Plant (Monocot) | Oryza sativa japonica | Crop species | |
| bdi | Plant (Monocot) | Brachypodium distachyon | Model grass related to cereal species | |
| oth | Plant (Monocot) | Oropetium thomaeum | Resurrection plant | Desiccation |
| zja | Plant (Monocot) | Zoysia japonica | Grass species | |
| zma | Plant (Monocot) | Zea mays | Crop species | |
| cme | Algae (Red algae) | Cyanidioschyzon merolae | Red algae representative | |
| esi | Algae (Brown algae) | Ectocapus siliculosus | Brown algae representative | |
| ptr | Algae (Diatom) | Phaedactylum tricornutum | Diatom representative | |
| pfa | Protist | Plasmodium falciparum | Medically important pathogen | |
| tth | Protist | Tetrahymena thermophila | Heat-tolerant cilliate | Heat |
| ddi | Protist (Amoeba) | Dictyostelium discoideum | Transitions from uni- to multicellular form | |
| sce | Fungi | Saccharomyces cerevisae | Model fungal species | Desiccation |
| ani | Fungi | Aspergillus nidulans | Model fungal species | |
| xbi | Fungi | Xeromyces bisporus | Highly xerophilic fungal species | Desiccation |
| cel | Animal (Invertebrate) | Caenorhabditis elegans | Model animal species | |
| hdu | Animal (Invertebrate) | Hypsibius dujardini | Tardigrade that is DT only after pretreatment | Desiccation |
| rva | Animal (Invertebrate) | Ramazzattius varieornatus | Extremely desiccation-tolerant tardigrade | Desiccation |
| ava | Animal (Invertebrate) | Adineta vaga | Desiccation tolerant rotifer | Desiccation |
| bmo | Animal (Invertebrate) | Bombyx mori | Lepidoptera representative | |
| ame | Animal (Invertebrate) | Apis mellifera | Hymenoptera representative | |
| dme | Animal (Invertebrate) | Drosophila melanogaster | Model insect species | |
| pnu | Animal (Invertebrate) | Polypedilum nubifer | Sensitive sister species to P. vanderplanki | |
| pva | Animal (Invertebrate) | Polypedilum vanderplanki | Anhydriobiotic animal (larval stage) | Desiccation |
| cgi | Animal (Invertebrate) | Crassostrea gigas | Mollusca representative | |
| hro | Animal (Invertebrate) | Helobdella robusta | Annelid representative | |
| cin | Animal (Invertebrate) | Ciona intestinalis | Basal chordate species | |
| pma | Animal (Vertebrate) | Petromyzon marinus | Basal vertebrate species | |
| pme | Animal (Vertebrate) | Poecilia mexicana | Has sulphur-tolerant sub-species | |
| gga | Animal (Vertebrate) | Gallus gallus | Bird model species | |
| mmu | Animal (Vertebrate) | Mus musculus | Model mammal species | |
| hgl | Animal (Vertebrate) | Heterocephalus glaber | Mammal with unique stress-related traits | See reference |
| clu | Animal (Vertebrate) | Canis lupis | Mammal representative | |
| hsa | Animal (Vertebrate) | Homo sapiens | Mammal representative |
References
- Fischer, W.W.; Hemp, J.; Valentine, J.S. How did life survive Earth’s great oxygenation? Curr. Opin. Chem. Biol. 2016, 31, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Clarendon Press: Oxford, UK, 1991. [Google Scholar]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. {ROS} Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.A.D.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Inupakutika, M.A.; Sengupta, S.; Devireddy, A.R.; Azad, R.K.; Mittler, R. The evolution of reactive oxygen species metabolism. J. Exp. Bot. 2016, 67, erw382. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K. V Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Meyer, Y.; Buchanan, B.B.; Vignols, F.; Reichheld, J.-P. Thioredoxins and glutaredoxins: Unifying elements in redox biology. Annu. Rev. Genet. 2009, 43, 335–367. [Google Scholar] [CrossRef]
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef]
- Maruta, T.; Sawa, Y.; Shigeoka, S.; Ishikawa, T. Diversity and evolution of ascorbate peroxidase functions in chloroplasts: More than just a classical antioxidant enzyme? Plant Cell Physiol. 2016, 57, 1377–1386. [Google Scholar] [CrossRef] [Green Version]
- Kehrer, J.P. The Haber—Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Bresgen, N.; Eckl, P. Oxidative stress and the homeodynamics of iron metabolism. Biomolecules 2015, 5, 808. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.-F. Superoxide dismutases: Ancient enzymes and new insights. FEBS Lett. 2012, 586, 585–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Singh, J.; Achary, V.; Reddy, M.K. Redox homeostasis via gene families of ascorbate-glutathione pathway. Front. Environ. Sci. 2015, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Sumimoto, H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J. 2008, 275, 3249–3277. [Google Scholar] [CrossRef]
- Lemire, J.; Alhasawi, A.; Appanna, V.P.; Tharmalingam, S.; Appanna, V.D. Metabolic defence against oxidative stress: The road less travelled so far. J. Appl. Microbiol. 2017, 123, 798–809. [Google Scholar] [CrossRef] [Green Version]
- Masip, L.; Veeravalli, K.; Georgiou, G. The many faces of glutathione in bacteria. Antioxid. Redox Signal. 2006, 8, 753–762. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. Are respiratory enzymes the primary sources of intracellular hydrogen peroxide? J. Biol. Chem. 2004, 279, 48742–48750. [Google Scholar] [CrossRef] [Green Version]
- Mendoza-Chamizo, B.; Løbner-Olesen, A.; Charbon, G. Coping with Reactive Oxygen Species to Ensure Genome Stability in Escherichia coli. Genes (Basel). 2018, 9, 565. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, G.; Ishikawa, T.; Pornsaksit, V.; Smirnoff, N. Evolution of alternative biosynthetic pathways for vitamin C following plastid acquisition in photosynthetic eukaryotes. Elife 2015, 4, e06369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loewus, F.A. Biosynthesis and metabolism of ascorbic acid in plants and of analogs of ascorbic acid in fungi. Phytochemistry 1999, 52, 193–210. [Google Scholar] [CrossRef]
- Koncarevic, S.; Rohrbach, P.; Deponte, M.; Krohne, G.; Prieto, J.H.; Yates, J.; Rahlfs, S.; Becker, K. The malarial parasite Plasmodium falciparum imports the human protein peroxiredoxin 2 for peroxide detoxification. Proc. Natl. Acad. Sci. USA 2009, 106, 13323–13328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penketh, P.G.; Kennedy, W.P.K.; Patton, C.L.; Sartorelli, A.C. Trypanosomatid hydrogen peroxidase metabolism. FEBS Lett. 1987, 221, 427–431. [Google Scholar] [CrossRef] [Green Version]
- O’neill, E.C.; Trick, M.; Hill, L.; Rejzek, M.; Dusi, R.G.; Hamilton, C.J.; Zimba, P.V; Henrissat, B.; Field, R.A. The transcriptome of Euglena gracilis reveals unexpected metabolic capabilities for carbohydrate and natural product biochemistry. Mol. Biosyst. 2015, 11, 2808–2820. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Tamaki, S.; Maruta, T.; Shigeoka, S. Biochemistry and Physiology of Reactive Oxygen Species in Euglena. In Euglena: Biochemistry, Cell and Molecular Biology; Springer: Berlin, Germany, 2017; pp. 47–64. [Google Scholar]
- Rampelotto, P.H. Extremophiles and extreme environments. Life (Basel) 2013, 3, 482. [Google Scholar] [CrossRef]
- Seckbach, J.; Rampelotto, P.H. Model Ecosystems in Extreme Environments; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Gostinčar, C.; Grube, M.; De Hoog, S.; Zalar, P.; Gunde-Cimerman, N. Extremotolerance in fungi: Evolution on the edge. FEMS Microbiol. Ecol. 2009, 71, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Horikawa, D.D.; Saito, Y.; Kuwahara, H.; Kozuka-Hata, H.; Shin, T.; Minakuchi, Y.; Ohishi, K.; Motoyama, A.; Aizu, T.; et al. Extremotolerant tardigrade genome and improved radiotolerance of human cultured cells by tardigrade-unique protein. Nat. Commun. 2016, 7, 12808. [Google Scholar] [CrossRef]
- Voituron, Y.; Barré, H.; Ramløv, H.; Douady, C.J. Freeze tolerance evolution among anurans: Frequency and timing of appearance. Cryobiology 2009, 58, 241–247. [Google Scholar] [CrossRef]
- Saldmann, F.; Viltard, M.; Leroy, C.; Friedlander, G. The Naked Mole Rat: A Unique Example of Positive Oxidative Stress. Oxid. Med. Cell. Longev. 2019, 2019, 1–7. [Google Scholar] [CrossRef]
- Oliver, M.; Tuba, Z.; Mishler, B. The evolution of vegetative desiccation tolerance in land plants. Plant Ecol. 2000, 151, 85–100. [Google Scholar] [CrossRef]
- Gaff, D.F.; Oliver, M. The evolution of desiccation tolerance in angiosperm plants: A rare yet common phenomenon. Funct. Plant Biol. 2013, 40, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Alpert, P. The limits and frontiers of desiccation-tolerant life. Integr. Comp. Biol. 2005, 45, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinton, H.E. A Fly Larva that tolerates Dehydration and Temperatures of −270 °C. Nature 1960, 188, 336. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Lyall, R.; Gechev, T. Multi-Omics Insights into the Evolution of Angiosperm Resurrection Plants. Annu. Plant Rev. Online 2020, 3, 77–110. [Google Scholar]
- Kranner, I.; Beckett, R.P.; Worknik, S.; Zorn, M.; Pfeifhofer, W. Antioxidants help the resurrection plant Myrothamnus flabellifolia survive desiccation. Plant J. 2002, 30, 1–13. [Google Scholar]
- Yoshida, Y.; Koutsovoulos, G.; Laetsch, D.R.; Stevens, L.; Kumar, S.; Horikawa, D.D.; Ishino, K.; Komine, S.; Kunieda, T.; Tomita, M.; et al. Comparative genomics of the tardigrades Hypsibius dujardini and Ramazzottius varieornatus. PLoS Biol. 2017, 15, e2002266. [Google Scholar] [CrossRef] [Green Version]
- Krisko, A.; Leroy, M.; Radman, M.; Meselson, M. Extreme anti-oxidant protection against ionizing radiation in bdelloid rotifers. Proc. Natl. Acad. Sci. USA 2012, 109, 2354–2357. [Google Scholar] [CrossRef] [Green Version]
- Boschetti, C.; Carr, A.; Crisp, A.; Eyres, I.; Wang-Koh, Y.; Lubzens, E.; Barraclough, T.G.; Micklem, G.; Tunnacliffe, A. Biochemical diversification through foreign gene expression in bdelloid rotifers. PLoS Genet. 2012, 8, e1003035. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.-C.D.; Ligterink, W.; Derks, M.F.L.; Farrant, J.M.; Artur, M.A.S.; Oliver, M.J.; Hesselink, T.; Jonkheer, E.; Maia, J.; Williams, B.; et al. A footprint of desiccation tolerance in the genome of Xerophyta viscosa. Nat. Plants 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Buren, R.; Wai, C.M.; Pardo, J.; Giarola, V.; Ambrosini, S.; Song, X.; Bartels, D. Desiccation tolerance evolved through gene duplication and network rewiring in Lindernia. Plant Cell 2018, 30, 2943–2958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Yang, G.; Zhang, L.; Yang, X.; Zhao, S.; Ji, Z.; Zhou, Q.; Hu, M.; Wang, Y.; Chen, M.; et al. The resurrection genome of Boea hygrometrica: A blueprint for survival of dehydration. Proc. Natl. Acad. Sci. USA 2015, 112, 5833–5837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanBuren, R.; Bryant, D.; Edger, P.P.; Tang, H.; Burgess, D.; Challabathula, D.; Spittle, K.; Hall, R.; Gu, J.; Lyons, E.; et al. Single-molecule sequencing of the desiccation-tolerant grass Oropetium thomaeum. Nature 2015, 527, 508–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowell, R.W.; Almeida, P.; Wilson, C.G.; Smith, T.P.; Fontaneto, D.; Crisp, A.; Micklem, G.; Tunnacliffe, A.; Boschetti, C.; Barraclough, T.G. Comparative genomics of bdelloid rotifers: Insights from desiccating and nondesiccating species. PLoS Biol. 2018, 16, e2004830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.-F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella Genome Reveals Evolutionary Insights into the Conquest of Land by Plants. Science 2007, 319, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Li, F.-W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; De Vries, J.; Delaux, P.-M.; Eily, A.; Koppers, N.; Kuo, L.-Y.; Li, Z.; et al. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460. [Google Scholar] [CrossRef] [Green Version]
- Wang, M. Characterizing the Cu-only SOD Repeat Protein of the Zebrafish Danio Rerio. Master’s Thesis, Johns Hopkins University, Baltimore, MD, USA, 2019. [Google Scholar]
- Zamocky, M.; Furtmüller, P.G.; Obinger, C. Evolution of catalases from bacteria to humans. Antioxid. Redox Signal. 2008, 10, 1527–1548. [Google Scholar] [CrossRef] [Green Version]
- Vlasits, J.; Jakopitsch, C.; Bernroitner, M.; Zamocky, M.; Furtmüller, P.G.; Obinger, C. Mechanisms of catalase activity of heme peroxidases. Arch. Biochem. Biophys. 2010, 500, 74–81. [Google Scholar] [CrossRef]
- Dayer, R.; Fischer, B.B.; Eggen, R.I.L.; Lemaire, S.D. The peroxiredoxin and glutathione peroxidase families in Chlamydomonas reinhardtii. Genetics 2008, 179, 41–57. [Google Scholar] [CrossRef] [Green Version]
- Zámocký, M.; Obinger, C. Molecular phylogeny of heme peroxidases. In Biocatalysis Based on Heme Peroxidases; Springer: Berlin, Germany, 2010; pp. 7–35. [Google Scholar]
- Hirt, R.P.; Müller, S.; Embley, T.M.; Coombs, G.H. The diversity and evolution of thioredoxin reductase: New perspectives. Trends Parasitol. 2002, 18, 302–308. [Google Scholar] [CrossRef]
- Novoselov, S.V; Gladyshev, V.N. Non-animal origin of animal thioredoxin reductases: Implications for selenocysteine evolution and evolution of protein function through carboxy-terminal extensions. Protein Sci. 2003, 12, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Lunde, C.; Baumann, U.; Shirley, N.J.; Drew, D.P.; Fincher, G.B. Gene Structure and Expression Pattern Analysis of Three Monodehydroascorbate Reductase (Mdhar) Genes in Physcomitrella patens: Implications for the Evolution of the {MDHAR} Family in Plants. Plant Mol. Biol. 2006, 60, 259–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahara, T.; Quinn, M.T.; Lambeth, J.D. Molecular evolution of the reactive oxygen-generating NADPH oxidase (Nox/Duox) family of enzymes. BMC Evol. Biol. 2007, 7, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Katsuyama, M. NOX/NADPH oxidase, the superoxide-generating enzyme: Its transcriptional regulation and physiological roles. J. Pharmacol. Sci. 2010, 114, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Considine, M.J.; Holtzapffel, R.C.; Day, D.A.; Whelan, J.; Millar, A.H. Molecular distinction between alternative oxidase from monocots and dicots. Plant Physiol. 2002, 129, 949–953. [Google Scholar] [CrossRef] [Green Version]
- Clifton, R.; Millar, A.H.; Whelan, J. Alternative oxidases in Arabidopsis: A comparative analysis of differential expression in the gene family provides new insights into function of non-phosphorylating bypasses. Biochim. Biophys. Acta (BBA) Bioenerg. 2006, 1757, 730–741. [Google Scholar] [CrossRef] [Green Version]
- Ragland, M.; Briat, J.F.; Gagnon, J.; Laulhere, J.-P.; Massenet, O.; Theil, E.C. Evidence for conservation of ferritin sequences among plants and animals and for a transit peptide in soybean. J. Biol. Chem. 1990, 265, 18339–18344. [Google Scholar]
- Grossman, M.J.; Hinton, S.M.; Minak-Bernero, V.; Slaughter, C.T.; Stiefel, E.I. Unification of the ferritin family of proteins. Proc. Natl. Acad. Sci. USA 1992, 89, 2419–2423. [Google Scholar] [CrossRef] [Green Version]
- Yévenes, A. The Ferritin Superfamily. In Macromolecular Protein Complexes; Springer: Cham, Switzerland, 2017; pp. 75–102. [Google Scholar]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef]
- Ding, Y.; Ortelli, F.; Rossiter, L.C.; Hemingway, J.; Ranson, H. The Anopheles gambiae glutathione transferase supergene family: Annotation, phylogeny and expression profiles. BMC Genomics 2003, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Oakley, A.J. Glutathione transferases: New functions. Curr. Opin. Struct. Biol. 2005, 15, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Yang, Z.-L.; Yang, X.; Liu, Y.-J.; Wang, X.-R.; Zeng, Q.-Y. Extensive functional diversification of the Populus glutathione S-transferase supergene family. Plant Cell 2009, 21, 3749–3766. [Google Scholar] [CrossRef] [Green Version]
- Dixon, D.P.; Edwards, R. Glutathione transferases. Arabidopsis Book 2010, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-J.; Han, X.-M.; Ren, L.-L.; Yang, H.-L.; Zeng, Q.-Y. Functional divergence of the glutathione S-transferase supergene family in Physcomitrella patens reveals complex patterns of large gene family evolution in land plants. Plant Physiol. 2013, 161, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Page, M.D.; Allen, M.D.; Kropat, J.; Urzica, E.I.; Karpowicz, S.J.; Hsieh, S.I.; Loo, J.A.; Merchant, S.S. Fe sparing and Fe recycling contribute to increased superoxide dismutase capacity in iron-starved Chlamydomonas reinhardtii. Plant Cell 2012, 24, 2649–2665. [Google Scholar] [CrossRef] [Green Version]
- Robinett, N.G.; Peterson, R.L.; Culotta, V.C. Eukaryotic copper-only superoxide dismutases ({SODs}): A new class of {SOD} enzymes and {SOD}-like protein domains. J. Biol. Chem. 2017, 293, 4636–4643. [Google Scholar] [CrossRef] [Green Version]
- Peterson, R.L.; Galaleldeen, A.; Villarreal, J.; Taylor, A.B.; Cabelli, D.E.; Hart, P.J.; Culotta, V.C. The Phylogeny and Active Site Design of Eukaryotic Copper-only Superoxide Dismutases. J. Biol. Chem. 2016, 291, 20911–20923. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Gube, M.; Schmidt, A.; Kothe, E. In silico analysis of nickel containing superoxide dismutase evolution and regulation. J. Basic Microbiol. 2009, 49, 109–118. [Google Scholar] [CrossRef]
- Horvath, M.M.; Grishin, N.V. The C-terminal domain of HPII catalase is a member of the type I glutamine amidotransferase superfamily. Proteins Struct. Funct. Bioinform. 2001, 42, 230–236. [Google Scholar] [CrossRef]
- Kawasaki, L.; Aguirre, J. Multiple catalase genes are differentially regulated in Aspergillus nidulans. J. Bacteriol. 2001, 183, 1434–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.-Q.; Yu, Q.-Y.; Zhang, Z. Annotation and evolution of the antioxidant gene in the silkworm, Bombyx mori. Arch. Insect Biochem. Physiol. 2012, 79, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family - an evolutionary overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef]
- Brawley, S.H.; Blouin, N.A.; Ficko-Blean, E.; Wheeler, G.L.; Lohr, M.; Goodson, H.V; Jenkins, J.W.; Blaby-Haas, C.E.; Helliwell, K.E.; Chan, C.X.; et al. Insights into the red algae and eukaryotic evolution from the genome of Porphyra umbilicalis (Bangiophyceae, Rhodophyta). Proc. Natl. Acad. Sci. USA 2017, 114, E6361–E6370. [Google Scholar] [CrossRef] [Green Version]
- McDonald, A.E.; Vanlerberghe, G.C.; Staples, J.F. Alternative oxidase in animals: Unique characteristics and taxonomic distribution. J. Exp. Biol. 2009, 212, 2627–2634. [Google Scholar] [CrossRef] [Green Version]
- Kanzok, S.M.; Fechner, A.; Bauer, H.; Ulschmid, J.K.; Müller, H.-M.; Botella-Munoz, J.; Schneuwly, S.; Schirmer, R.H.; Becker, K. Substitution of the thioredoxin system for glutathione reductase in Drosophila melanogaster. Science 2001, 291, 643–646. [Google Scholar] [CrossRef]
- Van Bel, M.; Diels, T.; Vancaester, E.; Kreft, L.; Botzki, A.; de Peer, Y.; Coppens, F.; Vandepoele, K. PLAZA 4.0: An integrative resource for functional, evolutionary and comparative plant genomics. Nucleic Acids Res. 2017, 46, D1190–D1196. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Brown, G.R.; Maglott, D.R. NCBI Reference Sequences (RefSeq): Current status, new features and genome annotation policy. Nucleic Acids Res. 2011, 40, D130–D135. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2018, 46, 2699. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2017, 46, D754–D761. [Google Scholar] [CrossRef] [PubMed]
- Acharjee, A.; Finkers, R.; Visser, R.G.; Maliepaard, C. Comparison of Regularized Regression Methods for ~Omics Data. Metabolomics Open Access 2013, 3. [Google Scholar] [CrossRef]
- Gusev, O.; Suetsugu, Y.; Cornette, R.; Kawashima, T.; Logacheva, M.D.; Kondrashov, A.S.; Penin, A.A.; Hatanaka, R.; Kikuta, S.; Shimura, S.; et al. Comparative genome sequencing reveals genomic signature of extreme desiccation tolerance in the anhydrobiotic midge. Nat. Commun. 2014, 5, 4784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.-J.; Zhang, Z.; Wang, J.-Y.; Oh, D.-H.; Dassanayake, M.; Liu, B.; Huang, Q.; Sun, H.-X.; Xia, R.; Wu, Y.; et al. Insights into salt tolerance from the genome of Thellungiella salsuginea. Proc. Natl. Acad. Sci. USA 2012, 109, 12219–12224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Lerat, E.; Daubin, V.; Moran, N.A. From gene trees to organismal phylogeny in prokaryotes: The case of the γ-Proteobacteria. PLoS Biol. 2003, 1, e19. [Google Scholar] [CrossRef]
- Blom, J.; Albaum, S.P.; Doppmeier, D.; Pühler, A.; Vorhölter, F.-J.; Zakrzewski, M.; Goesmann, A. EDGAR: A software framework for the comparative analysis of prokaryotic genomes. BMC Bioinform. 2009, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [Green Version]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.-S.; Chen, Y.-C.; Lu, C.-H.; Hwang, J.-K. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinform. 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar]









| Family | Sub-Family | Function and References |
|---|---|---|
| Superoxide dismutase (SOD) | Iron/Manganese SOD (Fe/Mn-SOD) | Catalyzes the detoxification of superoxide radicals into oxygen and hydrogen peroxide (H2O2). [14,51] |
| Copper/Zinc SOD (Cu/Zn-SOD) | ||
| Copper-only SOD (Cu-SOD) | ||
| Copper-only SOD-repeat protein (CSRP) | ||
| Nickel SOD (Ni-SOD) | ||
| Catalase (CAT) | Typical catalase | Catalyzes the dismutation of H2O2 into oxygen and water. [52,53] |
| Catalase-peroxidase | ||
| Manganese-containing catalase | ||
| Heme peroxidases (PRX) | Animal heme-PRX | Functions in the detoxification of H2O2 or peroxide radicals via the oxidation of a wide variety of organic and inorganic substrates, using either a heme or cysteine/selenocysteine cofactor. [54,55] |
| Ascorbate PRX (APX) | ||
| Ligninase (LiP/MnP/VP) | ||
| Classical plant PRX (POX) | ||
| DyP-type | ||
| Non-heme peroxidases (GPX, peroxiredoxins/Prx) | Glutathione PRX (GPX) | |
| Type-II Prx | ||
| PrxQ | ||
| 1-Cys Prx | ||
| 2-Cys Prx | ||
| Redoxins | Thioredoxin (TRX) | Involved in dithioldisulphide exchange, such as reduction of thiol-containing Prxs. [9] |
| Glutaredoxin (GRX) | ||
| Glutathione reductase (GR) | GR | NADPH-dependent maintenance of the cellular pool of reduced glutathione (GSH). [24] |
| Thioredoxin reductase (TR) | High-MW TR | NADPH-dependent maintenance of the cellular pool of reduced redoxins. [56,57] |
| Low-MW TR | ||
| Monodehydroascorbate reductase | MDHAR | Regenerates ascorbate (AsA) from MDHA radicals generated during the AsA-GSH cycle. [58] |
| NADPH oxidase (NOX) | NOX1-4 | Membrane-bound oxidases that specifically produce superoxide. [5,17,59,60] |
| NOX5 | ||
| DUOX | ||
| RBOH | ||
| NOXD | ||
| Alternative oxidase | AOX | Terminal oxidase in an alternative mitochondrial respiration pathway. [61,62] |
| Ferritin | Ferritin | Fe-sequestering proteins that play an essential role in iron homoeostasis. [63,64,65] |
| Glutathione S-transferase (GST) | Alpha | Catalyzes the conjugation of GSH to a multitude of activated xenobiotic substrates, including toxins and secondary metabolites. [66,67,68,69,70,71,72] |
| Delta | ||
| DHAR | ||
| Ef1Bg | ||
| Epsilon | ||
| Hemerythrin | ||
| Iota | ||
| Lambda | ||
| Mu | ||
| Omega | ||
| Phi | ||
| Pi | ||
| Sigma | ||
| Tau | ||
| THCQD | ||
| Theta | ||
| Zeta |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyall, R.; Nikoloski, Z.; Gechev, T. Comparative Analysis of ROS Network Genes in Extremophile Eukaryotes. Int. J. Mol. Sci. 2020, 21, 9131. https://doi.org/10.3390/ijms21239131
Lyall R, Nikoloski Z, Gechev T. Comparative Analysis of ROS Network Genes in Extremophile Eukaryotes. International Journal of Molecular Sciences. 2020; 21(23):9131. https://doi.org/10.3390/ijms21239131
Chicago/Turabian StyleLyall, Rafe, Zoran Nikoloski, and Tsanko Gechev. 2020. "Comparative Analysis of ROS Network Genes in Extremophile Eukaryotes" International Journal of Molecular Sciences 21, no. 23: 9131. https://doi.org/10.3390/ijms21239131
APA StyleLyall, R., Nikoloski, Z., & Gechev, T. (2020). Comparative Analysis of ROS Network Genes in Extremophile Eukaryotes. International Journal of Molecular Sciences, 21(23), 9131. https://doi.org/10.3390/ijms21239131