miR-155 Contributes to Normal Keratinocyte Differentiation and Is Upregulated in the Epidermis of Psoriatic Skin Lesions

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
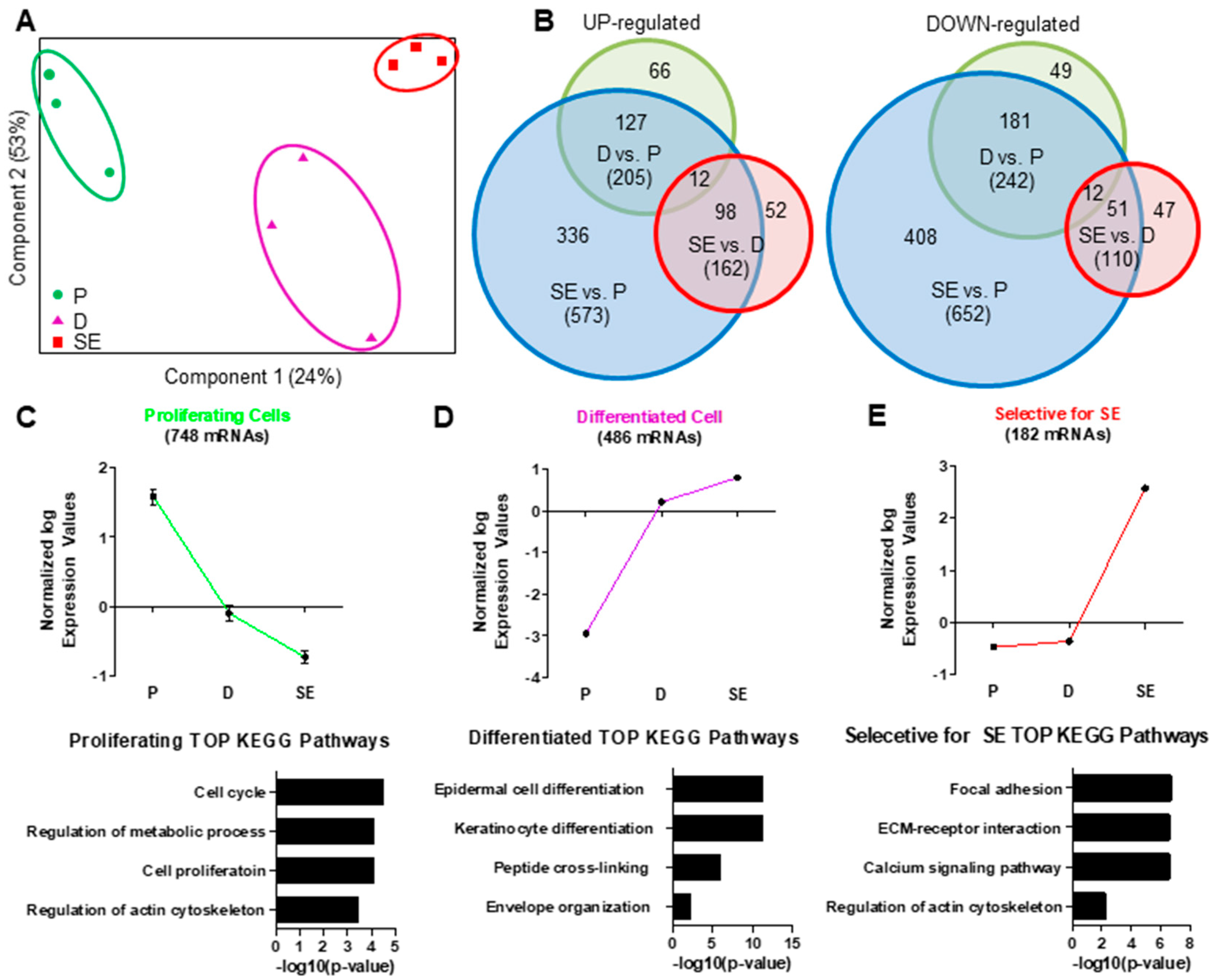
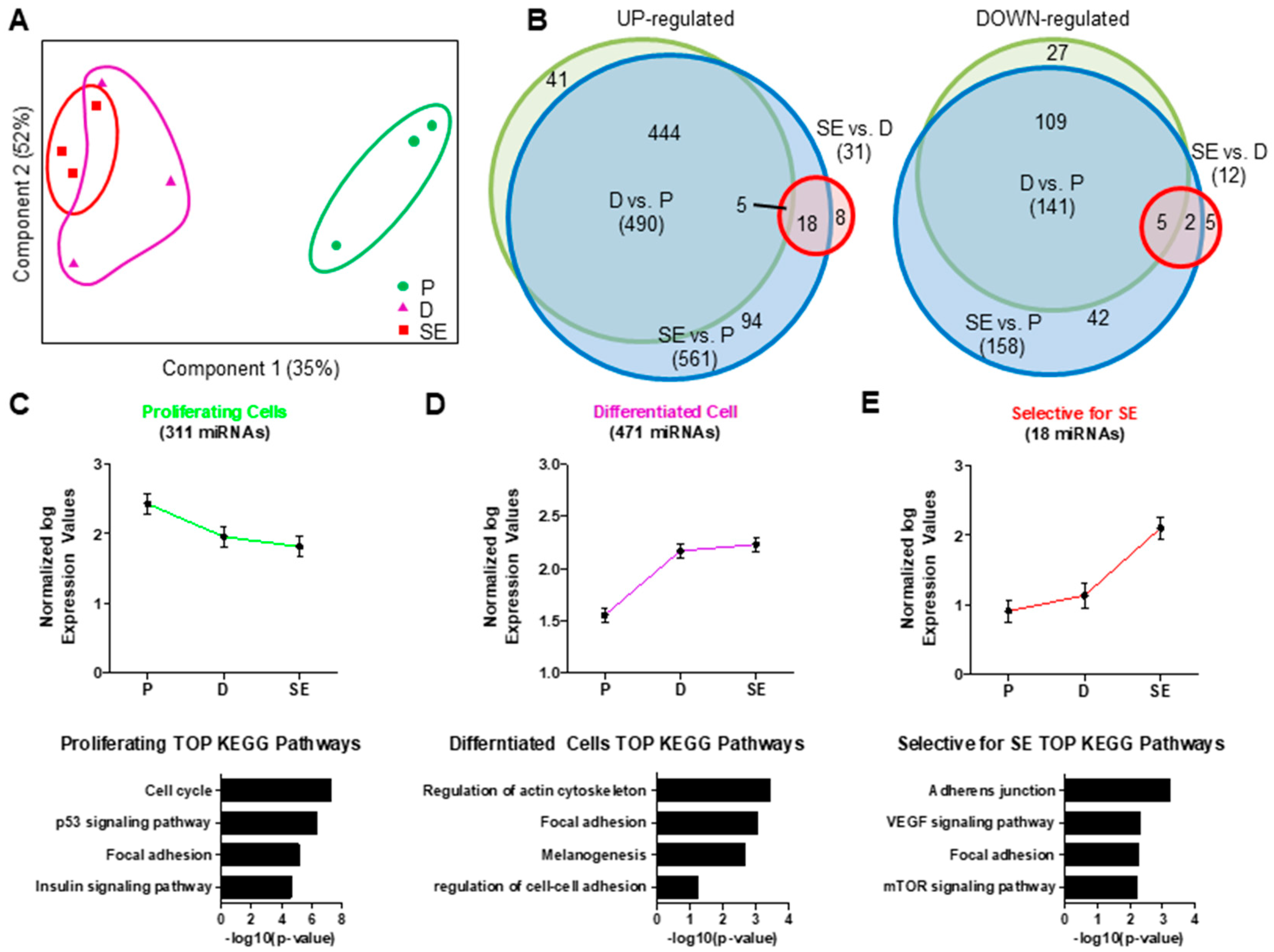
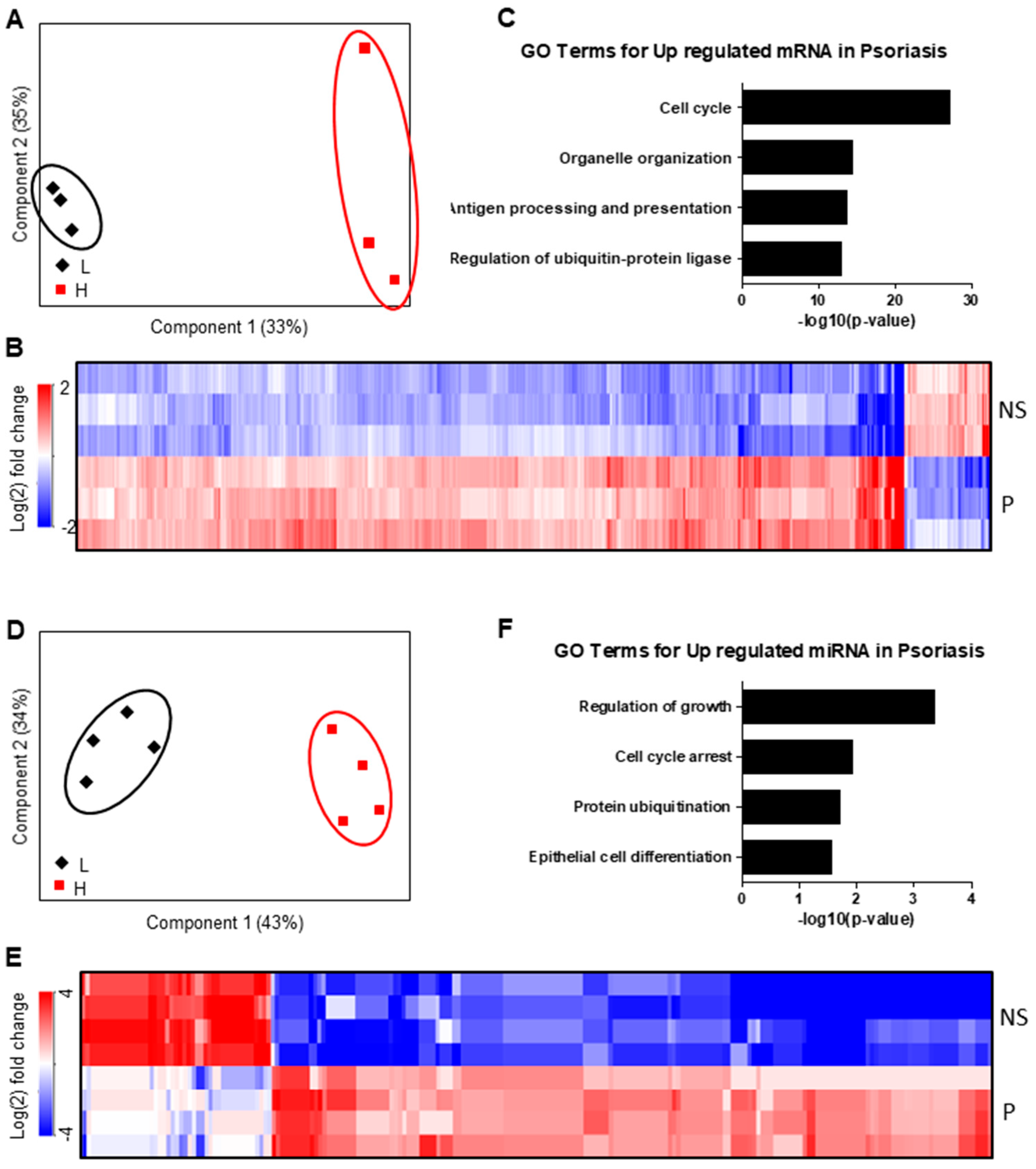
2.1. Regulation of mRNA and miRNA Expression During Keratinocyte Differentiation
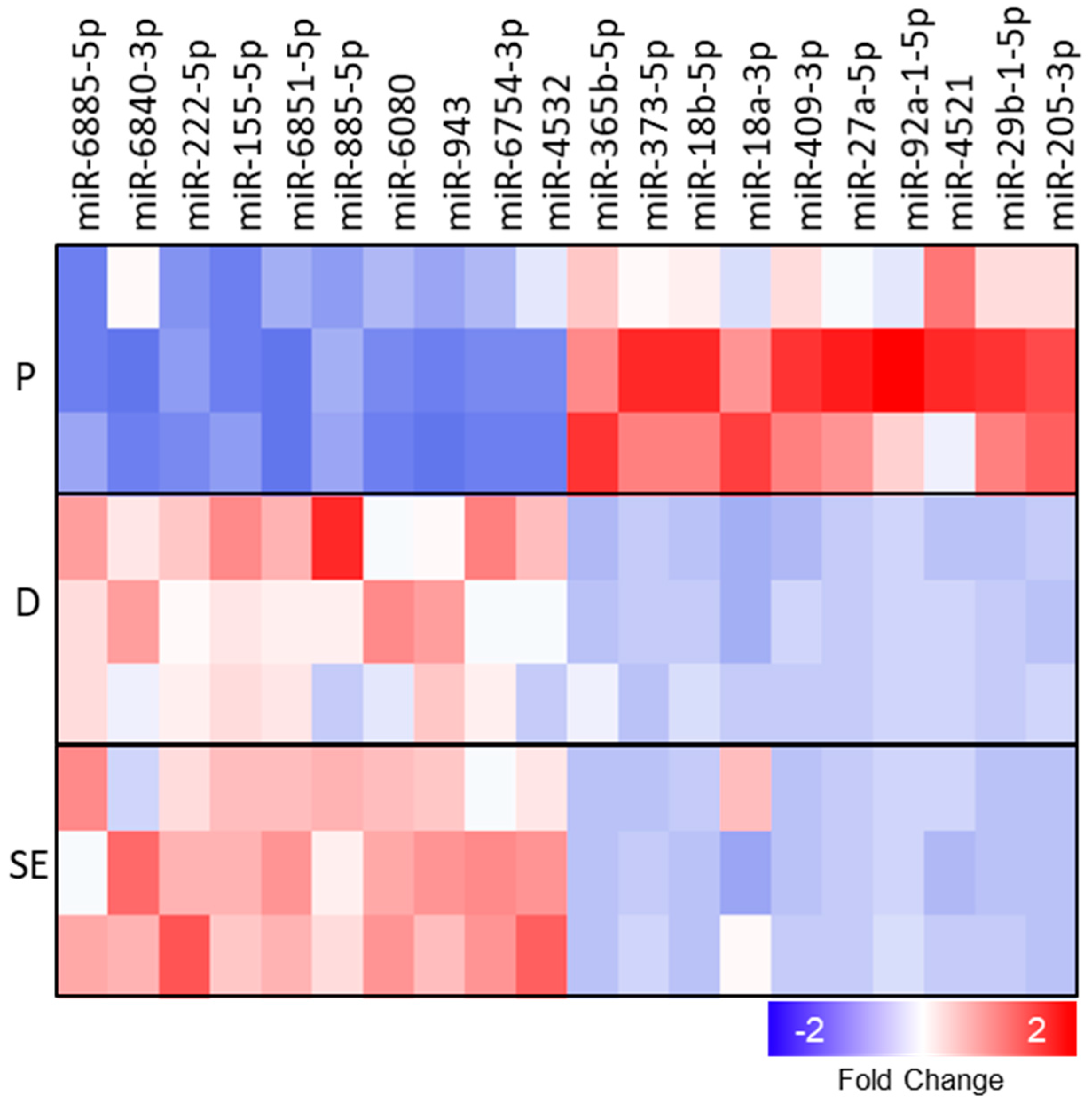
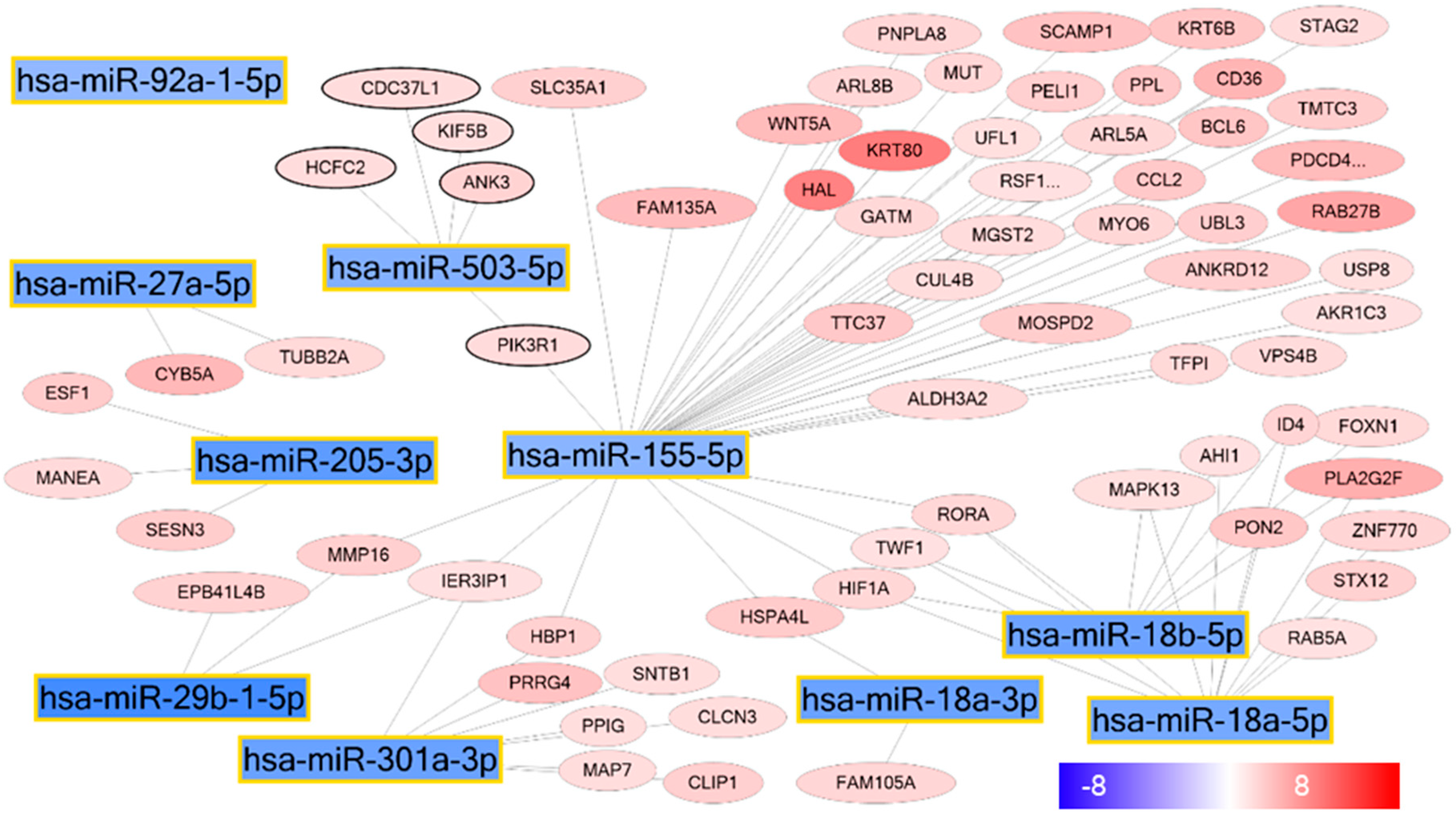
2.2. miRNA–mRNA Interaction Networks During KC Differentiation
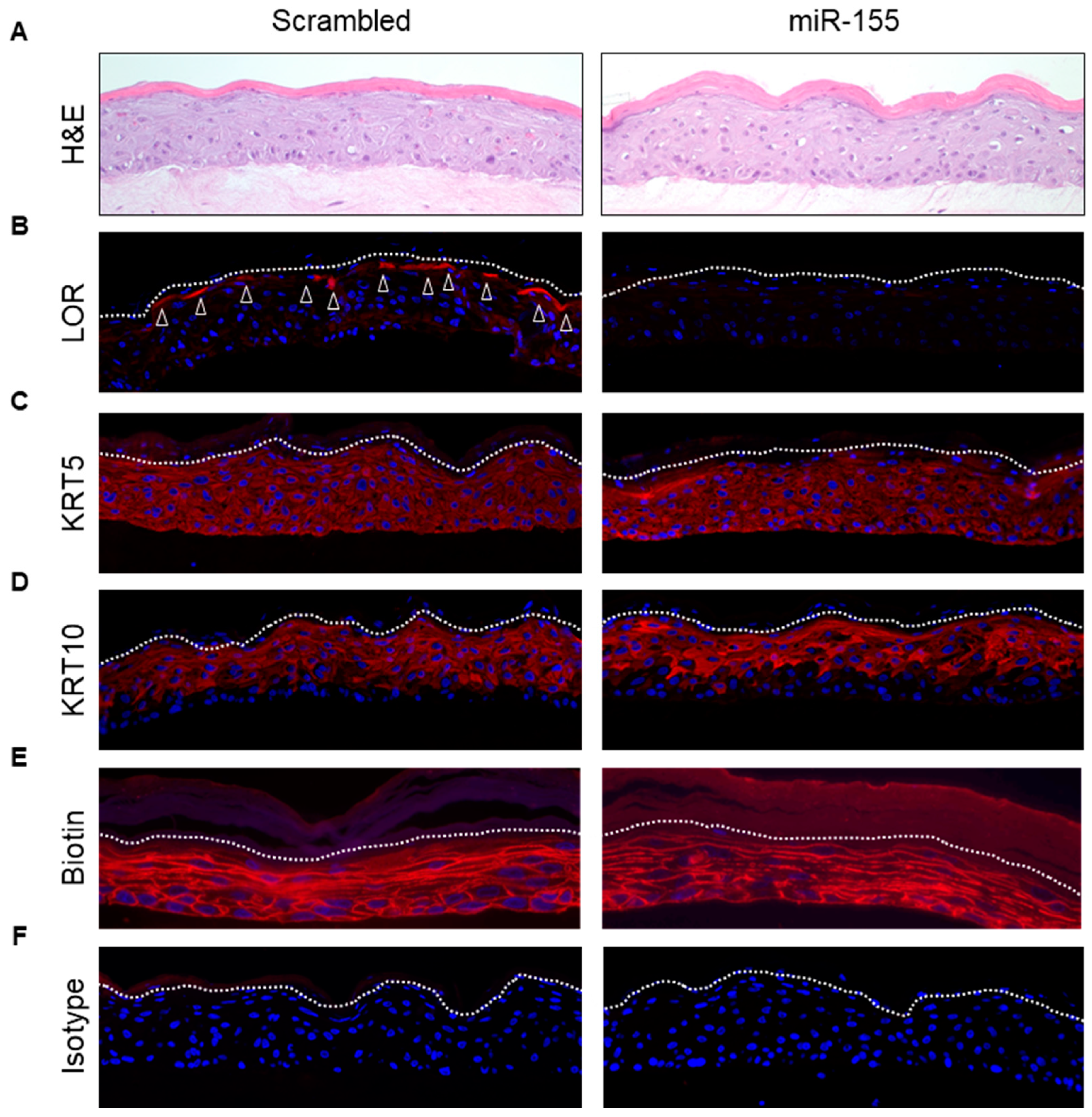
2.3. Over-Expression of miR-155 in a Human Skin Model Affects KC Differentiation
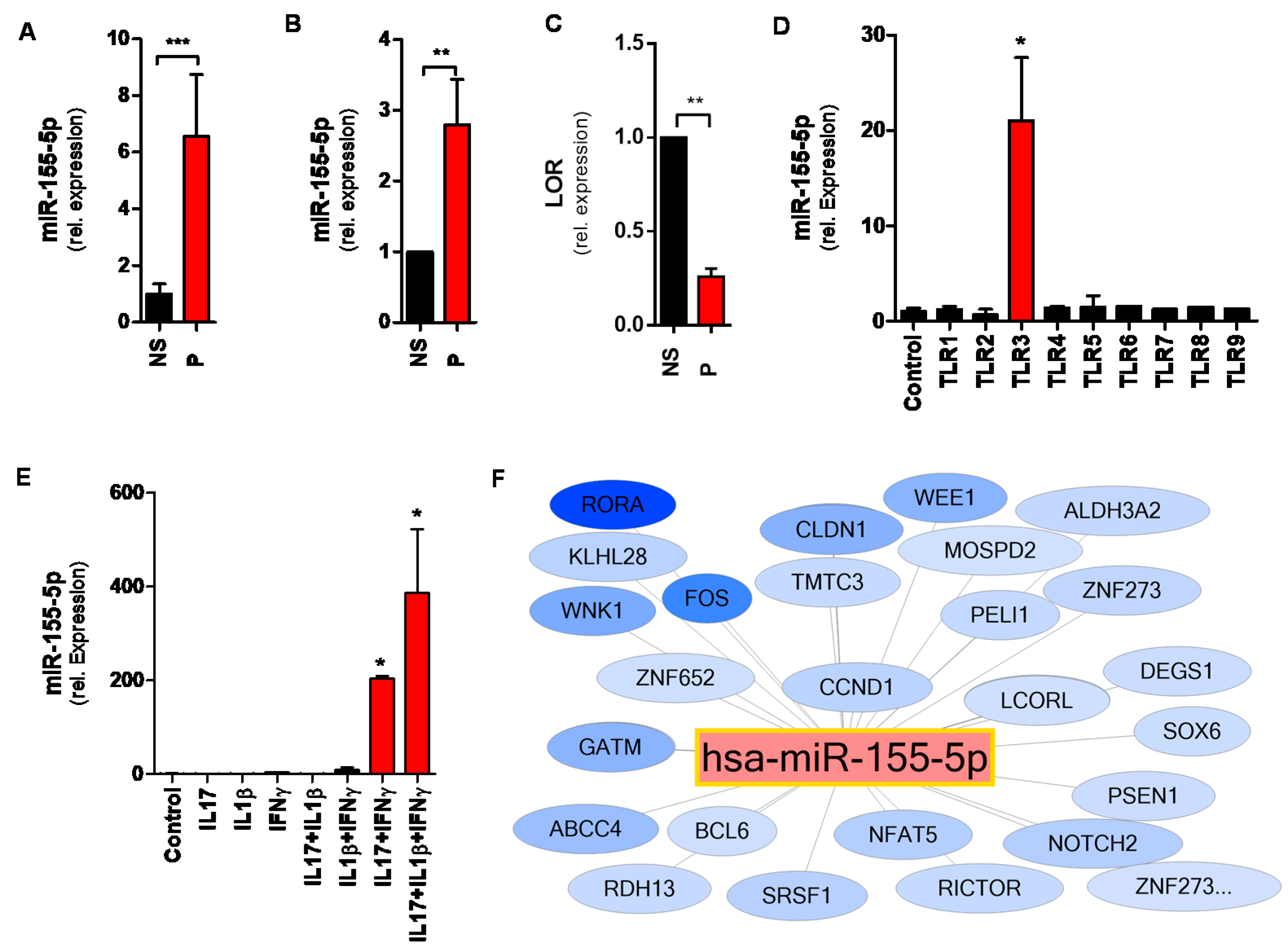
2.4. MiR-155 Is Upregulated in the Epidermis of Psoriatic Skin Lesions
2.5. TLR3 and Inflammatory Cytokines Upregulate miR-155 in Human Keratinocytes
2.6. The miR-155 Network in Psoriasis
3. Discussion
4. Materials and Methods
4.1. Psoriasis Samples
4.2. Cell culture and Preparation of Organotypic Skin Cultures
4.3. RNA Extraction and cDNA Synthesis
4.4. Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis of miR-155
4.5. qPCR of mRNA
4.6. miR-155 Mimics Transfection
4.7. mRNA and miRNA Microarrays
4.8. Microarray Expression Data Analysis
4.9. Functional Annotation Clustering and Pathway Analysis
4.10. Immunofluorescence Staining
4.11. Analysis of Epidermal Permeability
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALDH3A2 | Aldehyde Dehydrogenase 3 Family Member A2 |
| ANOVA | Analysis of variance |
| CLDN1 | Claudin-1 |
| DEGS1 | Delta 4-Desaturase, Sphingolipid 1 |
| dKC | Differentiated keratinocytes |
| ECM | Extracellular matrix |
| FOS | Fos Proto-Oncogene, AP-1 Transcription Factor Subunit |
| GO | Gene ontology |
| HAL | histidine ammonia-lyase |
| IL-1β | Interleukin-1β |
| IL-17 | Interleukin-17 |
| INF-γ | Interferon gamma |
| IVN | Involucrin |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| KC | Keratinocyte |
| KRT5 | Keratin 5 |
| KRT10 | Keratin 10 |
| KRT80 | Keratin 80 |
| LCORL | Ligand Dependent Nuclear Receptor Corepressor Like |
| LOR | Loricrin |
| miRNA | microRNA |
| mTOR | mammalian target of Rapamycin |
| NFAT | Nuclear factor of activated T-cells |
| NOTH2 | Neurogenic Locus Notch Homolog Protein 2 |
| NS | Normal skin |
| PCA | Principal component analysis |
| pKC | Proliferating keratinocyte |
| PRR | Pattern recognition receptor |
| RICTOR | RPTOR Independent Companion Of MTOR Complex 2 |
| RORA | Retinoic acid receptor related orphan receptor A |
| SE | Skin equivalent |
| SOX6 | SRY-Box Transcription Factor 6 |
| TLR | Toll-like receptor |
| TNFα | Tumor necrosis factor alpha |
| VEGF | Vascular endothelial growth factor |
| WEE1 | WEE1 G2 Checkpoint Kinase |
| ZNF273 | Zinc Finger Protein 273 |
| ZNF652 | Zinc Finger Protein 652 |
References
- Fuchs, E.; Raghavan, S. Getting under the skin of epidermal morphogenesis. Nat. Rev. Genet. 2002, 3, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Blumenberg, M. SKINOMICS: Transcriptional profiling in dermatology and skin biology. Curr. Genom. 2012, 13, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckhart, L.; Lippens, S.; Tschachler, E.; Declercq, W. Cell death by cornification. Biochim. Biophys. Acta 2013, 1833, 3471–3480. [Google Scholar] [CrossRef]
- Perdigoto, C.N.; Valdes, V.J.; Bardot, E.S.; Ezhkova, E. Epigenetic regulation of epidermal differentiation. Cold Spring. Harb. Perspect. Med. 2014, 4, a015263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Pajares, V.; Yan, K.; Zarnegar, B.J.; Jameson, K.L.; Khavari, P.A. Genetic pathways in disorders of epidermal differentiation. Trends Genet 2013, 29, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Blumenberg, M. In Skinomics, transcriptional profiling approaches to molecular and structural biology of epidermis. Semin. Cutan. Med. Surg. 2019, 2019, E12–E18. [Google Scholar] [CrossRef]
- Blanpain, C.; Fuchs, E. Epidermal homeostasis: A balancing act of stem cells in the skin. Nat. Rev. Mol. Cell Biol. 2009, 10, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Kular, L.; Vij, M.; Herter, E.K.; Li, X.; Wang, A.; Chu, T.; Toma, M.-A.; Zhang, L.; Liapi, E. Human skin long noncoding RNA WAKMAR1 regulates wound healing by enhancing keratinocyte migration. Proc. Natl. Acad. Sci. USA 2019, 116, 9443–9452. [Google Scholar] [CrossRef] [Green Version]
- Terlecki-Zaniewicz, L.; Pils, V.; Bobbili, M.R.; Lämmermann, I.; Perrotta, I.; Grillenberger, T.; Schwestka, J.; Weiß, K.; Pum, D.; Arcalis, E. Extracellular vesicles in human skin: Cross-talk from senescent fibroblasts to keratinocytes by miRNAs. J. Investig. Dermatol. 2019, 139, 2425–2436. [Google Scholar] [CrossRef] [Green Version]
- Wagner, T.; Beer, L.; Gschwandtner, M.; Eckhart, L.; Kalinina, P.; Laggner, M.; Ellinger, A.; Gruber, R.; Kuchler, U.; Golabi, B. The differentiation-associated keratinocyte protein cornifelin contributes to cell-cell adhesion of epidermal and mucosal keratinocytes. J. Investig. Dermatol. 2019, 139, 2292–2301. [Google Scholar] [CrossRef]
- Finnegan, A.I.; Cho, R.; Luu, A.; Harirchian, P.; Lee, J.; Cheng, J.; Song, J.S. Single-cell transcriptomics reveals spatial and temporal turnover of keratinocyte differentiation regulators. Front. Genet. 2019, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Meisgen, F.; Pasquali, L.; Munkhammar, S.; Xia, P.; Ståhle, M.; Landén, N.X.; Pivarcsi, A.; Sonkoly, E. Next generation sequencing identifies the keratinocyte-specific miRNA signature of psoriasis. J. Investig. Dermatol. 2019, 139, 2547–2550. [Google Scholar] [CrossRef] [PubMed]
- Lena, A.; Shalom-Feuerstein, R.; di Val Cervo, P.R.; Aberdam, D.; Knight, R.; Melino, G.; Candi, E. miR-203 represses ‘stemness’ by repressing ΔNp63. Cell Death Differ. 2008, 15, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, H.; Li, C.; Zhang, X.; Zhou, P.; Xiao, X.; Zhang, W.; Wu, Y.; Zeng, R.; Wang, B. Nicastrin/miR-30a-3p/RAB31 Axis Regulates Keratinocyte Differentiation by Impairing EGFR Signaling in Familial Acne Inversa. J. Investig. Dermatol. 2019, 139, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, J.E.; Nguyen, G.H.; Fujita, M.; Florell, S.R.; Duffin, K.C.; Krueger, G.G.; O’Connell, R.M. microRNAs in Psoriasis. J. Investig. Dermatol. 2016, 136, 365–371. [Google Scholar] [CrossRef]
- Hildebrand, J.; Rütze, M.; Walz, N.; Gallinat, S.; Wenck, H.; Deppert, W.; Grundhoff, A.; Knott, A. A comprehensive analysis of microRNA expression during human keratinocyte differentiation in vitro and in vivo. J. Investig. Dermatol. 2011, 131, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Singhvi, G.; Manchanda, P.; Rapalli, V.K.; Dubey, S.K.; Gupta, G.; Dua, K. MicroRNAs as biological regulators in skin disorders. Biomed. Pharmacother. 2018, 108, 996–1004. [Google Scholar] [CrossRef]
- Hoffjan, S.; Stemmler, S. On the role of the epidermal differentiation complex in ichthyosis vulgaris, atopic dermatitis and psoriasis. Br. J. Dermatol. 2007, 157, 441–449. [Google Scholar] [CrossRef]
- Guinea-Viniegra, J.; Jiménez, M.; Schonthaler, H.B.; Navarro, R.; Delgado, Y.; Concha-Garzón, M.J.; Tschachler, E.; Obad, S.; Daudén, E.; Wagner, E.F. Targeting miR-21 to treat psoriasis. Sci. Transl. Med. 2014, 6, 225re1. [Google Scholar] [CrossRef]
- Timis, T.L.; Orasan, R.I. Understanding psoriasis: Role of miRNAs. Biomed. Rep. 2018, 9, 367–374. [Google Scholar]
- Ma, L.; Xue, H.B.; Wang, F.; Shu, C.M.; Zhang, J.H. MicroRNA-155 may be involved in the pathogenesis of atopic dermatitis by modulating the differentiation and function of T helper type 17 (Th17) cells. Clin. Exp. Immunol. 2015, 181, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebane, A.; Akdis, C.A. MicroRNAs: Essential players in the regulation of inflammation. J. Allergy Clin. Immunol. 2013, 132, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Van Gele, M.; Bracke, S.; Medeiros, A.K.D.A.; Lambert, J. Exploring the feasibility of whole blood to identify systemic miRNA biomarkers for patients with moderate-to-severe psoriasis. Eur. J. Dermatol. 2016, 26, 195–198. [Google Scholar] [CrossRef]
- García–Rodríguez, S.; Arias–Santiago, S.; Blasco-Morente, G.; Orgaz-Molina, J.; Rosal-Vela, A.; Navarro, P.; Magro-Checa, C.; Martínez-López, A.; Ruiz, J.C.; Raya, E. Increased expression of microRNA-155 in peripheral blood mononuclear cells from psoriasis patients is related to disease activity. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Moura, J.; Sørensen, A.; Leal, E.C.; Svendsen, R.; Carvalho, L.; Willemoes, R.J.; Jørgensen, P.T.; Jenssen, H.; Wengel, J.; Dalgaard, L.T. MicroRNA-155 inhibition restores Fibroblast Growth Factor 7 expression in diabetic skin and decreases wound inflammation. Sci. Rep. 2019, 9, 5836. [Google Scholar] [CrossRef]
- Xu, L.; Leng, H.; Shi, X.; Ji, J.; Fu, J. MiR-155 promotes cell proliferation and inhibits apoptosis by PTEN signaling pathway in the psoriasis. Biomed. Pharmacother. 2017, 90, 524–530. [Google Scholar] [CrossRef]
- Hou, R.; Liu, R.; Zhao, X.; Jia, Y.; An, P.; Hao, Z.; Li, J.; Li, X.; Yin, G.; Zhang, K. Increased miR-155-5p expression in dermal mesenchymal stem cells of psoriatic patients: Comparing the microRNA expression profile by microarray. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Soonthornchai, W.; Tangtanatakul, P.; Meephansan, J.; Ruchusatsawat, K.; Reantragoon, R.; Hirankarn, N.; Wongpiyabovorn, J. Down-regulation of miR-155 after treatment with narrow-band UVB and methotrexate associates with apoptosis of keratinocytes in psoriasis. Asian Pac. J. Allergy Immunol. 2019. [Google Scholar] [CrossRef]
- De Koning, H.D.; Rodijk-Olthuis, D.; van Vlijmen-Willems, I.M.; Joosten, L.A.; Netea, M.G.; Schalkwijk, J.; Zeeuwen, P.L. A comprehensive analysis of pattern recognition receptors in normal and inflamed human epidermis: Upregulation of dectin-1 in psoriasis. J. Investig. Dermatol. 2010, 130, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.J.; Sen, G.L.; Ward, N.L.; Johnston, A.; Chun, K.; Chen, Y.; Adase, C.; Sanford, J.A.; Gao, N.; Chensee, M.; et al. Antimicrobial Peptide LL37 and MAVS Signaling Drive Interferon-beta Production by Epidermal Keratinocytes during Skin Injury. Immunity 2016, 45, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Pivarcsi, A.; Nagy, I.; Koreck, A.; Kis, K.; Kenderessy-Szabo, A.; Szell, M.; Dobozy, A.; Kemeny, L. Microbial compounds induce the expression of pro-inflammatory cytokines, chemokines and human β-defensin-2 in vaginal epithelial cells. Microbes Infect. 2005, 7, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Brodin, P.; Wei, T.; Meisgen, F.; Eidsmo, L.; Nagy, N.; Kemeny, L.; Ståhle, M.; Sonkoly, E.; Pivarcsi, A. MiR-125b, a microRNA downregulated in psoriasis, modulates keratinocyte proliferation by targeting FGFR2. J. Investig. Dermatol. 2011, 131, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; Poy, M.N.; Stoffel, M.; Fuchs, E. A skin microRNA promotes differentiation by repressing ‘stemness’. Nature 2008, 452, 225–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonkoly, E.; Wei, T.; Janson, P.C.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A. MicroRNAs: Novel regulators involved in the pathogenesis of psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toulza, E.; Mattiuzzo, N.R.; Galliano, M.-F.; Jonca, N.; Dossat, C.; Jacob, D.; de Daruvar, A.; Wincker, P.; Serre, G.; Guerrin, M. Large-scale identification of human genes implicated in epidermal barrier function. Genome Biol. 2007, 8, R107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhou, L.; Dai, J. Retinoic acid receptor-related orphan receptor RORα regulates differentiation and survival of keratinocytes during hypoxia. J. Cell. Physiol. 2018, 233, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Barresi, C.; Stremnitzer, C.; Mlitz, V.; Kezic, S.; Kammeyer, A.; Ghannadan, M.; Posa-Markaryan, K.; Selden, C.; Tschachler, E.; Eckhart, L. Increased sensitivity of histidinemic mice to UVB radiation suggests a crucial role of endogenous urocanic acid in photoprotection. J. Investig. Dermatol. 2011, 131, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmayr, M.; André, E.; Conquet, F.; Rondi-Reig, L.; Delhaye-Bouchaud, N.; Auclair, N.; Daniel, H.; Crépel, F.; Mariani, J.; Sotelo, C.; et al. staggerer phenotype in retinoid-related orphan receptor alpha-deficient mice. Proc. Natl. Acad. Sci. USA 1998, 95, 3960–3965. [Google Scholar] [CrossRef] [Green Version]
- Mehrel, T.; Hohl, D.; Rothnagel, J.A.; Longley, M.A.; Bundman, D.; Cheng, C.; Lichti, U.; Bisher, M.E.; Steven, A.C.; Steinert, P.M.; et al. Identification of a major keratinocyte cell envelope protein, loricrin. Cell 1990, 61, 1103–1112. [Google Scholar] [CrossRef]
- Hohl, D.; Mehrel, T.; Lichti, U.; Turner, M.L.; Roop, D.R.; Steinert, P.M. Characterization of human loricrin. Structure and function of a new class of epidermal cell envelope proteins. J. Biol. Chem. 1991, 266, 6626–6636. [Google Scholar]
- Koch, P.J.; de Viragh, P.A.; Scharer, E.; Bundman, D.; Longley, M.A.; Bickenbach, J.; Kawachi, Y.; Suga, Y.; Zhou, Z.; Huber, M.; et al. Lessons from loricrin-deficient mice: Compensatory mechanisms maintaining skin barrier function in the absence of a major cornified envelope protein. J. Cell Biol. 2000, 151, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Ishitsuka, Y.; Roop, D.R. Loricrin Confers Photoprotective Function against UVB in Corneocytes. J. Investig. Dermatol. 2018, 138, 2684–2687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonkoly, E.; Janson, P.; Majuri, M.-L.; Savinko, T.; Fyhrquist, N.; Eidsmo, L.; Xu, N.; Meisgen, F.; Wei, T.; Bradley, M. MiR-155 is overexpressed in patients with atopic dermatitis and modulates T-cell proliferative responses by targeting cytotoxic T lymphocyte–associated antigen 4. J. Allergy Clin. Immunol. 2010, 126, 581–589.e20. [Google Scholar] [CrossRef]
- Šahmatova, L.; Tankov, S.; Prans, E.; Aab, A.; Hermann, H.; Reemann, P.; Pihlap, M.; Karelson, M.; Abram, K.; Kisand, K.; et al. MicroRNA-155 is dysregulated in the skin of patients with vitiligo and inhibits melanogenesis-associated genes in melanocytes and keratinocytes. Acta Dermato Venereologica 2016, 96, 742–748. [Google Scholar]
- Meisgen, F.; Xu Landén, N.; Bouez, C.; Zuccolo, M.; Gueniche, A.; Ståhle, M.; Sonkoly, E.; Breton, L.; Pivarcsi, A. Activation of toll-like receptors alters the microRNA expression profile of keratinocytes. Exp. Dermatol. 2014, 23, 281–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fry, L.; Baker, B. Triggering psoriasis: The role of infections and medications. Clin. Dermatol. 2007, 25, 606–615. [Google Scholar] [CrossRef]
- Wagner, T.; Gschwandtner, M.; Strajeriu, A.; Elbe-Burger, A.; Grillari, J.; Grillari-Voglauer, R.; Greiner, G.; Golabi, B.; Tschachler, E.; Mildner, M. Establishment of keratinocyte cell lines from human hair follicles. Sci. Rep. 2018, 8, 13434. [Google Scholar] [CrossRef]
- Mildner, M.; Jin, J.; Eckhart, L.; Kezic, S.; Gruber, F.; Barresi, C.; Stremnitzer, C.; Buchberger, M.; Mlitz, V.; Ballaun, C. Knockdown of filaggrin impairs diffusion barrier function and increases UV sensitivity in a human skin model. J. Investig. Dermatol. 2010, 130, 2286–2294. [Google Scholar] [CrossRef] [Green Version]
- Beer, L.; Mlitz, V.; Gschwandtner, M.; Berger, T.; Narzt, M.S.; Gruber, F.; Brunner, P.M.; Tschachler, E.; Mildner, M. Bioinformatics approach for choosing the correct reference genes when studying gene expression in human keratinocytes. Exp. Dermatol. 2015, 24, 742–777. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic acids research 2002, 30, e36. [Google Scholar] [CrossRef]
- Beer, L.; Seemann, R.; Ristl, R.; Ellinger, A.; Kasiri, M.M.; Mitterbauer, A.; Zimmermann, M.; Gabriel, C.; Gyöngyösi, M.; Klepetko, W. High dose ionizing radiation regulates micro RNA and gene expression changes in human peripheral blood mononuclear cells. BMC Genom. 2014, 15, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beer, L.; Kalinina, P.; Köcher, M.; Laggner, M.; Jeitler, M.; Abbas Zadeh, S.; Copic, D.; Tschachler, E.; Mildner, M. miR-155 Contributes to Normal Keratinocyte Differentiation and Is Upregulated in the Epidermis of Psoriatic Skin Lesions. Int. J. Mol. Sci. 2020, 21, 9288. https://doi.org/10.3390/ijms21239288
Beer L, Kalinina P, Köcher M, Laggner M, Jeitler M, Abbas Zadeh S, Copic D, Tschachler E, Mildner M. miR-155 Contributes to Normal Keratinocyte Differentiation and Is Upregulated in the Epidermis of Psoriatic Skin Lesions. International Journal of Molecular Sciences. 2020; 21(23):9288. https://doi.org/10.3390/ijms21239288
Chicago/Turabian StyleBeer, Lucian, Polina Kalinina, Martin Köcher, Maria Laggner, Markus Jeitler, Salman Abbas Zadeh, Dragan Copic, Erwin Tschachler, and Michael Mildner. 2020. "miR-155 Contributes to Normal Keratinocyte Differentiation and Is Upregulated in the Epidermis of Psoriatic Skin Lesions" International Journal of Molecular Sciences 21, no. 23: 9288. https://doi.org/10.3390/ijms21239288
APA StyleBeer, L., Kalinina, P., Köcher, M., Laggner, M., Jeitler, M., Abbas Zadeh, S., Copic, D., Tschachler, E., & Mildner, M. (2020). miR-155 Contributes to Normal Keratinocyte Differentiation and Is Upregulated in the Epidermis of Psoriatic Skin Lesions. International Journal of Molecular Sciences, 21(23), 9288. https://doi.org/10.3390/ijms21239288