Immunological Pattern in IgA Nephropathy

Abstract
:1. Introduction
2. Results
2.1. Immunophenotype of Leukocyte Subpopulations
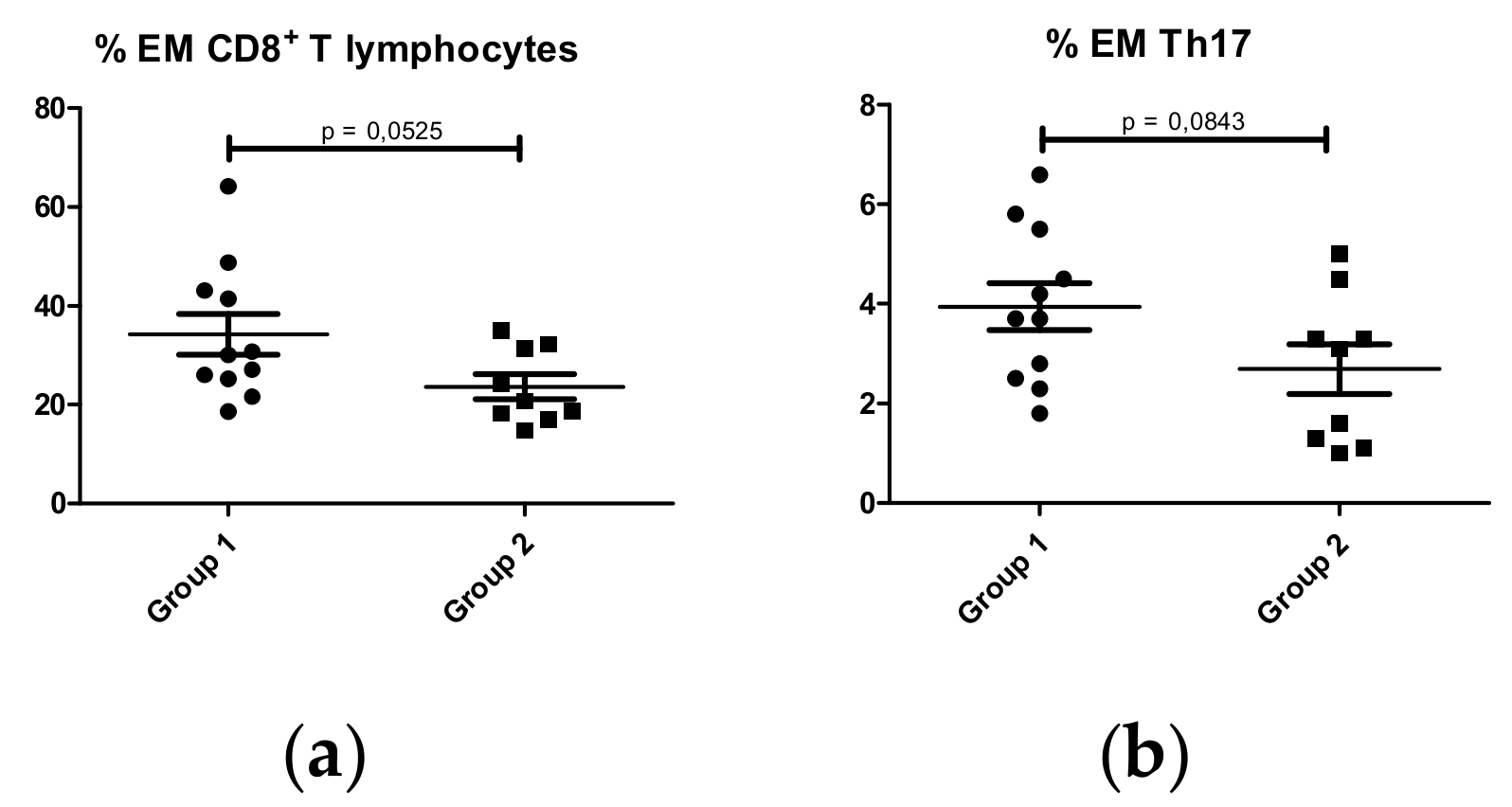
2.1.1. T lymphocyte Subpopulations
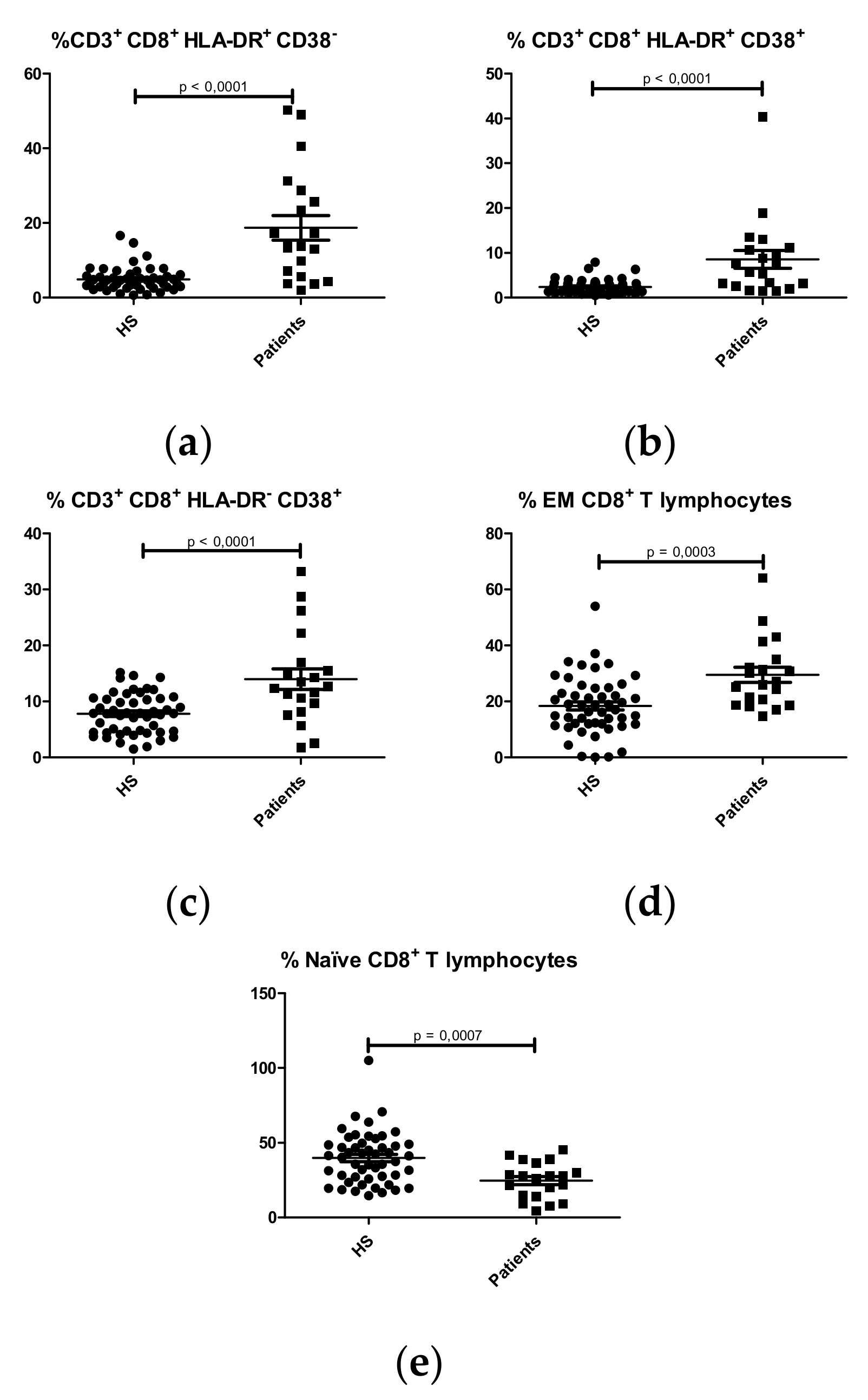
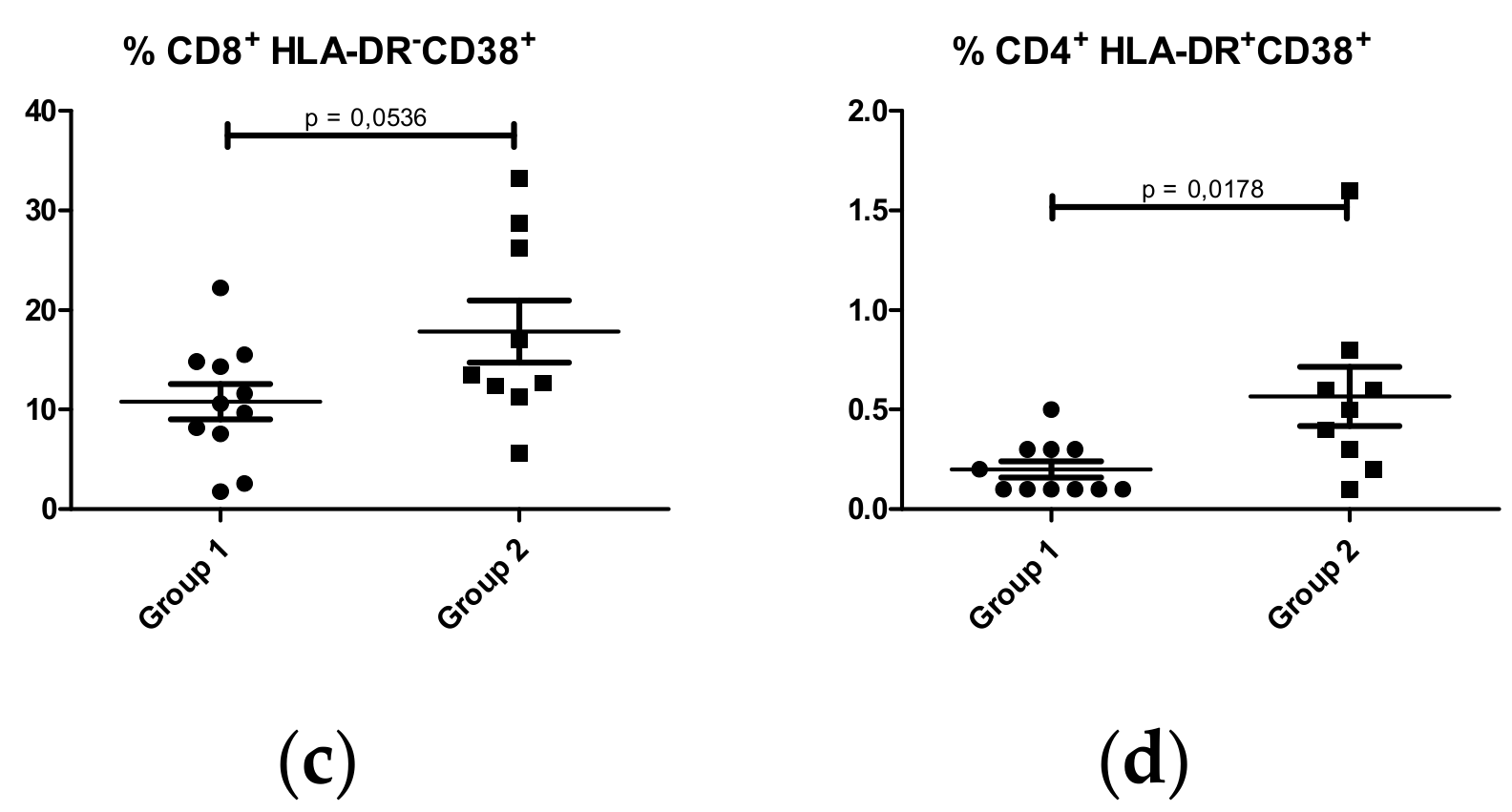
CD8+ T Lymphocytes
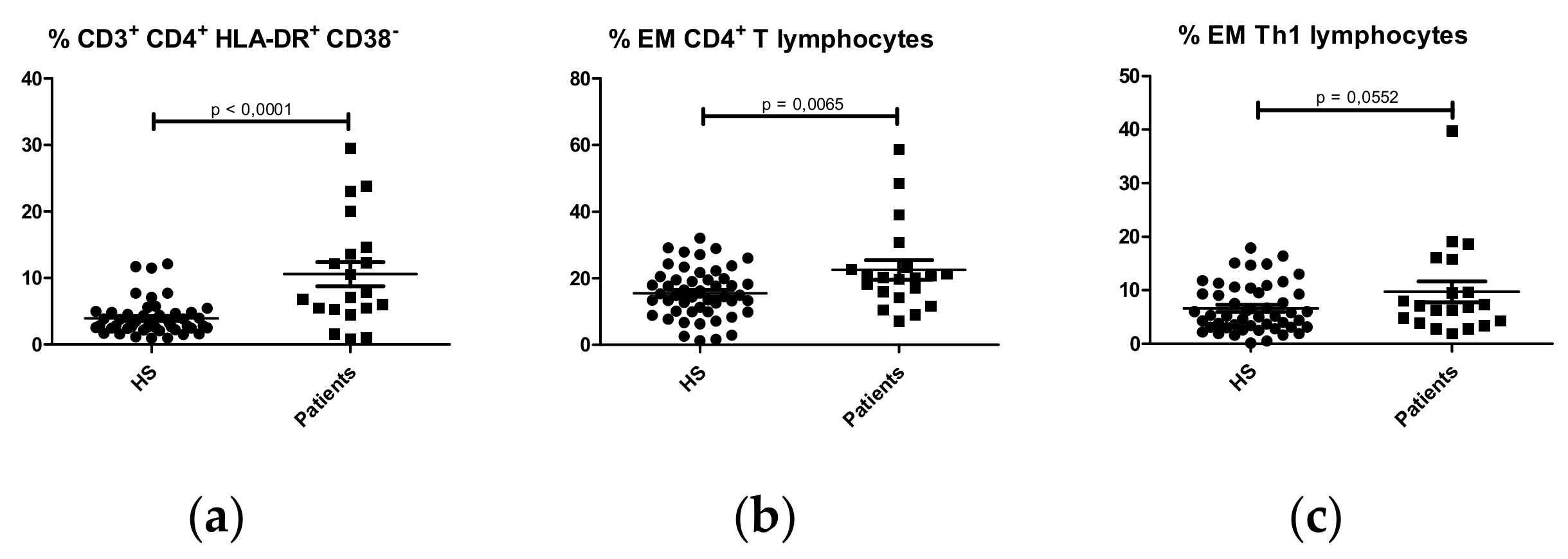
CD4+ T lymphocytes
2.1.2. B lymphocyte Subpopulations
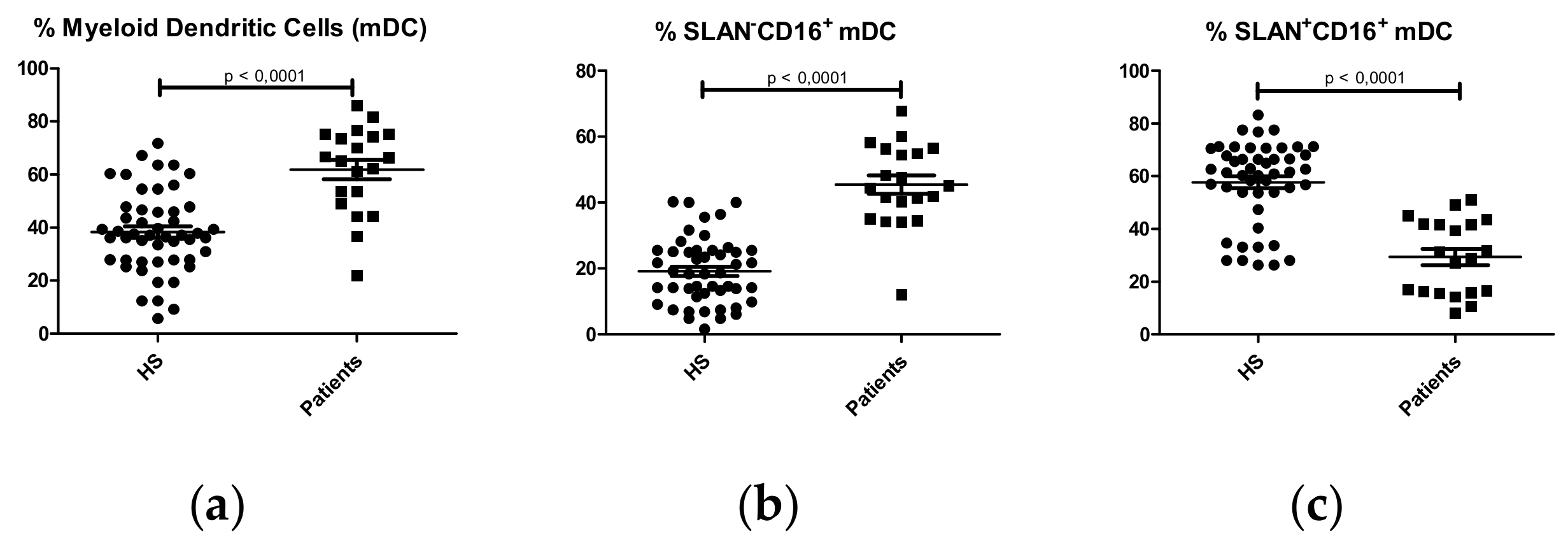
2.1.3. Dendritic Cell Subpopulations
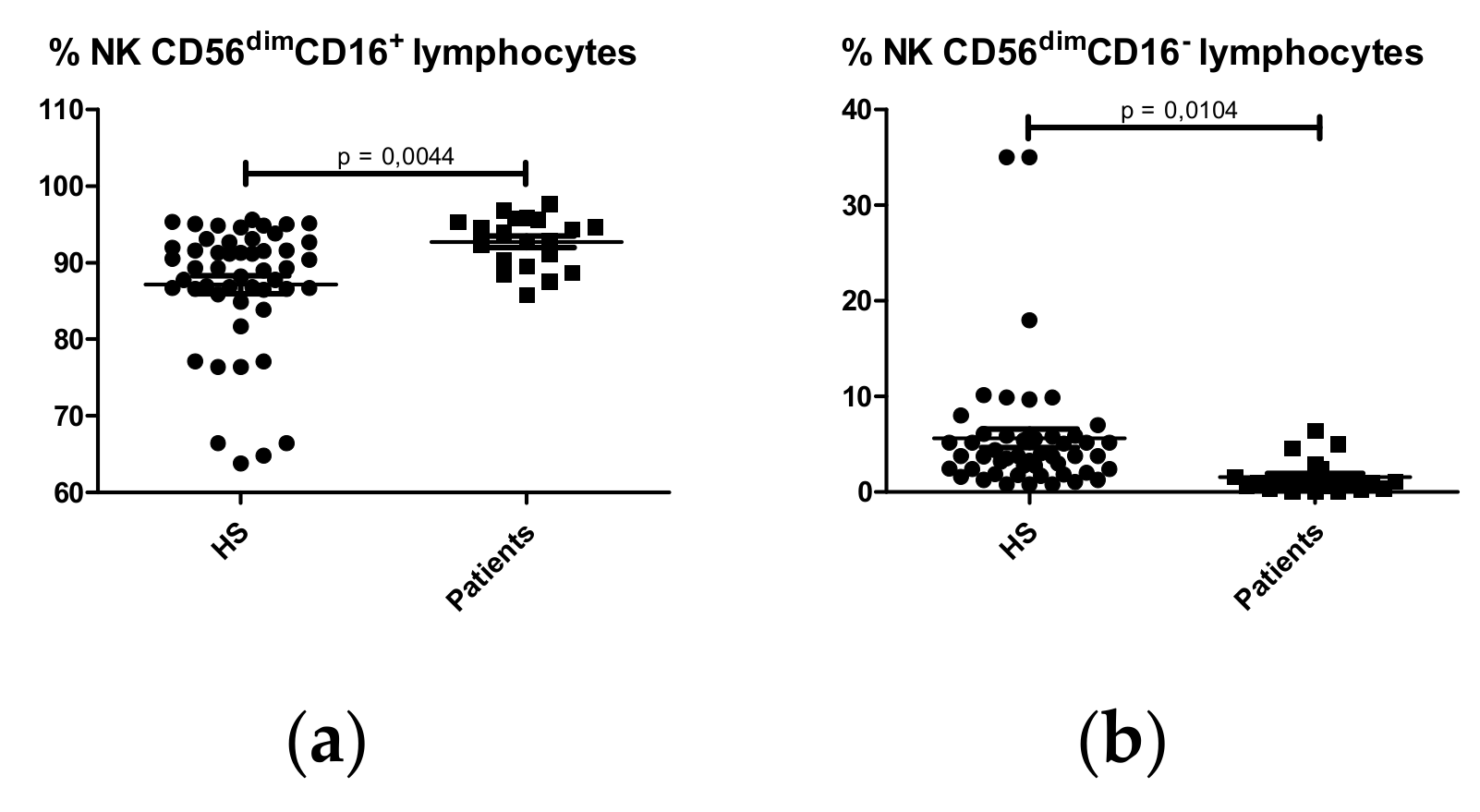
2.1.4. NK Cell Subpopulations
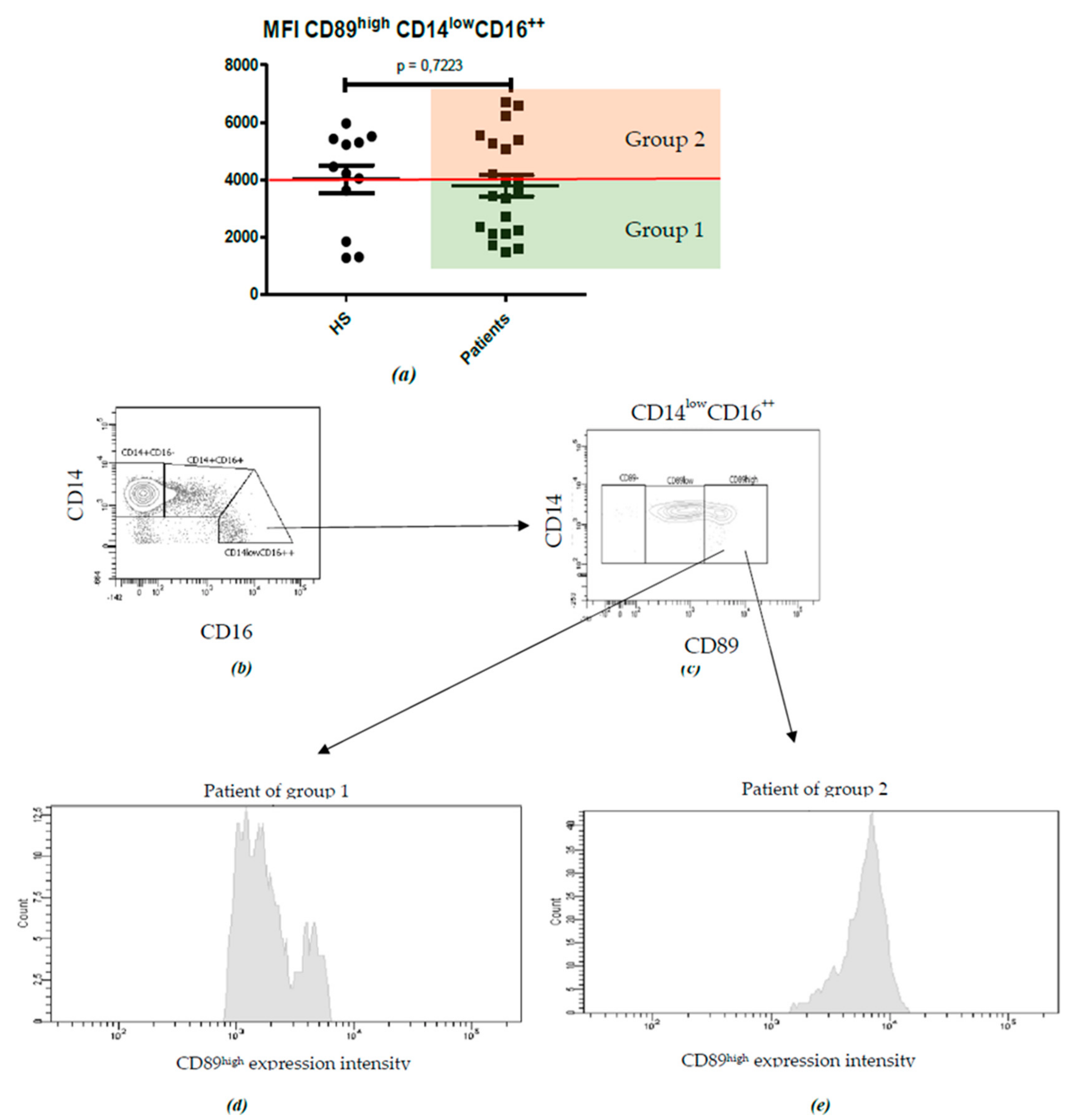
2.2. Characterization of Monocyte Subpopulations
2.3. Serum Levels of Galactose Deficient IgA1
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Immunological Characterization and Flow Cytometric Analysis
4.3. Serum Levels of Galactose Deficient IgA1
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boyd, J.K.; Cheung, C.K.; Molyneux, K.; Feehally, J.; Barratt, J. An update on the pathogenesis and treatment of IgA nephropathy. Kidney Int. 2012, 81, 833–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barratt, J.; Feehally, J. IgA nephropathy. J. Am. Soc. Nephrol. 2005, 16, 2088–2097. [Google Scholar] [CrossRef] [PubMed]
- Working Group of the International IgA Nephropathy Network and the Renal Pathology Society; Roberts, I.S.D.; Cook, H.T.; Troyanov, S.; Alpers, C.E.; Amore, A.; Barratt, J.; Berthoux, F.; Bonsib, S.; Bruijn, J.A.; et al. The Oxford classification of IgA nephropathy: Pathology definitions, correlations, and reproducibility. Kidney Int. 2009, 76, 546–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magistroni, R.; D’Agati, V.D.; Appel, G.B.; Kiryluk, K. New developments in the genetics, pathogenesis, and therapy of IgA nephropathy. Kidney Int. 2015, 88, 974–989. [Google Scholar] [CrossRef] [Green Version]
- Coppo, R. Clinical and histological risk factors for progression of IgA nephropathy: an update in children, young and adult patients. J Nephrol. 2017, 30, 339–346. [Google Scholar] [CrossRef]
- Ali, G.G.; Zina, M.; Robert, J.W.; Catherine, V.B.; Susan, Y.W.; Richard, P.L.; Jiri, M.; Jam, N.; Bruce, A.J. Aberrant IgA1 Glycosylation Is Inherited in Familial and Sporadic IgA Nephropathy. J. Am. Soc. Nephrol. 2008, 19, 1008–1014. [Google Scholar] [CrossRef]
- Segarra, A. Avances en el conocimiento de la patogenia de la nefropatía IgA: ¿nuevas perspectivas para un futuro inmediato? Nefrologia 2010, 30, 501–507. [Google Scholar] [CrossRef]
- Segarra-medrano, A.; Carnicer-caceres, C.; Valtierra-carmeno, N. Estudio de las variables asociadas a la activación local del complemento en la nefropatía IgA idiopática. Nefrologí 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Monteiro, R.C. Role of IgA and IgA Fc receptors in inflammation. J. Clin. Immunol. 2010, 30, 1–9. [Google Scholar] [CrossRef]
- Novak, J.; Julian, B.A.; Mestecky, J.; Renfrow, M.B. Glycosylation of IgA1 and pathogenesis of IgA nephropathy. Semin Immunopathol. 2012, 34, 365–382. [Google Scholar] [CrossRef]
- Oortwijn, B.D.; Roos, A.; Royle, L.; van Gijlswijk-Janssen, D.J.; Faber-Krol, M.C.; Eijgenraam, J.W.; Dwek, R.A.; Daha, W.R.; Rudd, P.M.; van Kooten, C. Differential Glycosylation of Polymeric and Monomeric IgA: A Possible Role in Glomerular Inflammation in IgA Nephropathy. J. Am. Soc. Nephrol. 2006, 17, 3529–3539. [Google Scholar] [CrossRef] [PubMed]
- Moura, I.C.; Arcos-Fajardo, M.; Sadaka, C.; Leroy, V.; Benhamou, M.; Novak, J.; Vrtovsnik, F.; Haddad, E.; Chintalacharuvu, K.R.; Monteiro, R.C. Glycosylation and Size of IgA1 Are Essential for Interaction with Mesangial Transferrin Receptor in IgA Nephropathy. J. Am. Soc. Nephrol. 2004, 15, 622–634. [Google Scholar] [CrossRef] [Green Version]
- Moura, I.C.; Centelles, M.N.; Arcos-Fajardo, M.; Malheiros, D.M.; Collawn, J.F.; Cooper, M.D.; Monteiro, R.C. Identification of the transferrin receptor as a novel immunoglobulin (Ig)A1 receptor and its enhanced expression on mesangial cells in Iga nephropathy. J. Exp. Med. 2001, 194, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Boyette, L.B.; MacEdo, C.; Hadi, K.; Elinoff, B.D.; Walters, J.T.; Ramaswami, B.; Chalasani, G.; Taboas, J.M.; Lakkis, F.G.; Metes, D.M. Phenotype, function, and differentiation potential of human monocyte subsets. PLoS ONE 2017, 12, e0176460. [Google Scholar] [CrossRef]
- Sprangers, S.; Vries, T.J.; de Everts, V. Monocyte Heterogeneity: Consequences for Monocyte-Derived Immune Cells. J. Immunol. Res. 2016, 2016, 1475435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, S.N.; Serino, G.; Sallustio, F.; Blasi, A.; Rossini, M.; Pesce, F.; Schena, F.P. Altered monocyte expression and expansion of non-classical monocyte subset in IgA nephropathy patients. Nephrol. Dial. Transplant. 2015, 30, 1122–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthelot, L.; Robert, T.; Vuiblet, V.; Tabary, T.; Braconnier, A.; Dramé, M.; Toupance, O.; Rieu, P.; Monteiro, R.C.; Touré, F. Recurrent IgA nephropathy is predicted by altered glycosylated IgA, autoantibodies and soluble CD89 complexes. Kidney Int. 2015, 88, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Launay, P.; Grossetête, B.; Arcos-Fajardo, M.; Gaudin, E.; Torres, S.P.; Beaudoin, L.; de Serre, N.P.-M.; Lehuen, A.; Monteiroa, R.C. Fcalpha receptor (CD89) mediates the development of immunoglobulin A (IgA) nephropathy (Berger’s disease). Evidence for pathogenic soluble receptor-Iga complexes in patients and CD89 transgenic mice. J. Exp. Med. 2000, 191, 1999–2009. [Google Scholar] [CrossRef] [Green Version]
- Working Group of the International IgA Nephropathy Network and the Renal Pathology Society; Cattran, D.C.; Coppo, R.; Cook, H.T.; Feehally, J.; Roberts, I.S.; Troyanov, S.; Alpers, C.E.; Amore, A.; Barratt, J.; et al. The Oxford classification of IgA nephropathy: rationale, clinicopathological correlations, and classification. Kidney Int. 2009, 76, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Coppo, R.; Troyanov, S.; Bellur, S.; Cattran, D.; Cook, H.T.; Feehally, J.; Roberts, I.S.D.; Morando, L.; Camilla, R.; Tesar, V.; et al. Validation of the Oxford classification of IgA nephropathy in cohorts with different presentations and treatments. Kidney Int. 2014, 86, 828–836. [Google Scholar] [CrossRef] [Green Version]
- Boltjes, A.; van Wijk, F. Human dendritic cell functional specialization in steady-state and inflammation. Front. Immunol. 2014, 5, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, E.; Touzot, M.; Bohineust, A.; Cappuccio, A.; Chiocchia, G.; Hosmalin, A.; Dalod, M.; Soumelis, V.; Amigorena, S. Human Inflammatory Dendritic Cells Induce Th17 Cell Differentiation. Immunity 2013, 38, 336–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Poli, A.; Michel, T.; Thérésine, M.; Andrès, E.; Hentges, F.; Zimmer, J. CD56bright natural killer (NK) cells: An important NK cell subset. Immunology 2009, 126, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Robert, T.; Berthelot, L.; Cambier, A.; Rondeau, E.; Monteiro, R.C. Molecular Insights into the Pathogenesis of IgA Nephropathy. Trends Mol. Med. 2015, 21, 762–775. [Google Scholar] [CrossRef]
- Novak, J.; Vu, H.L.; Novak, L.; Julian, B.A.; Mestecky, J.; Tomana, M. Interactions of human mesangial cells with IgA and IgA-containing immune complexes. Kidney Int. 2002, 62, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Novak, J.; Tomana, M.; Matousovic, K.; Brown, R.; Hall, S.; Novak, L.; Julian, B.A.; Wyatt, R.J.; Mestecky, J. IgA1-containing immune complexes in IgA nephropathy differentially affect proliferation of mesangial cells. Kidney Int. 2005, 67, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Matsuzaki, K.; Suzuki, H.; Okazaki, K.; Yanagawa, H.; Ieiri, N.; Sato, M.; Sato, T.; Taguma, Y.; Matsuoka, J.; et al. Serum levels of galactose-deficient immunoglobulin (Ig) A1 and related immune complex are associated with disease activity of IgA nephropathy. Clin. Exp. Nephrol. 2014, 18, 770–777. [Google Scholar] [CrossRef] [Green Version]
- Yanagawa, H.; Suzuki, H.; Suzuki, Y.; Kiryluk, K.; Gharavi, A.; Matsuoka, K.; Makita, Y.; Julian, B.A.; Novak, J.; Tomino, Y. A panel of serum biomarkers differentiates IgA nephropathy from other renal diseases. PLoS ONE 2014, 9, e98081. [Google Scholar] [CrossRef] [Green Version]
- Popadic, D.; Anegon, I.; Baeten, D.; Eibel, H.; Giese, T.; Marits, P.; Martinez-Caceres, E.; Mascart, F.; Nestle, F.; Pujol-Borrell, R.; et al. Predictive immunomonitoring—the COST ENTIRE initiative. Clin. Immunol. 2013, 147, 23–26. [Google Scholar] [CrossRef]
- Maecker, H.T. Standardizing immunophenotyping for the Human Immunology. Nat. Rev. Immunol. 2012, 12, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirant-Sánchez, B.; Hervás-García, J.V.; Teniente-Serra, A.; Brieva, L.; Moral-Torres, E.; Cano, A.; Munteis, E.; Mansilla, M.J.; Presas-Rodriguez, S.; Navarro-Barriuso, J.; et al. Predicting therapeutic response to fingolimod treatment in multiple sclerosis patients. CNS Neurosci. Ther. 2018, 24, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Yasutake, J.; Suzuki, Y.; Suzuki, H.; Hiura, N.; Yanagawa, H.; Makita, Y.; Kaneko, E.; Tomino, Y. Novel lectin-independent approach to detect galactose-deficient IgA1 in IgA nephropathy. Nephrol. Dial. Transplant. 2015, 30, 1315–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 (N = 13) CD89high MFI < 4000 | Group 2 (N = 9) CD89high MFI > 4000 | P Value | |
|---|---|---|---|
| Oxford | |||
| M1 | 13 (100%) | 9 (100%) | - |
| S1 | 10 (76.92%) | 2 (22.22%) | 0.0274 |
| E1 | 2 (15.38%) | 2 (22.22%) | 1.000 |
| T1-2 | 7 (53.84%) | 1 (11.11%) | 0.0669 |
| IgAN Patients (N = 22) | Normality Range | |
|---|---|---|
| Gender (Male), n (%) | 14 (63.63%) | - |
| Age, years | 50.63 | - |
| IgA, mg/dL | 368.55 ± 228.39 | 70–400 |
| C3, mg/dL | 116.41 ± 9.89 | 90-180 |
| C4, mg/dL | 27.78 ± 1.41 | 10–40 |
| eGFR, mL/min/1,73m2 | 53 ± 26.16 | >90 |
| Serum creatinine, mg/dL | 1.76 ± 3.03 | 0.72–1.18 |
| Proteinuria, mg/g creatinine) | 1212.14 ± 1811.89 | 0–200 |
| Hematuria | 7 ± 15.90 | <3 |
| Oxford classification | ||
| M1 | 22 (100%) | - |
| S1 | 12 (54.54%) | - |
| E1 | 4 (18.18%) | - |
| T1-2 | 7 (31.82%) | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esteve Cols, C.; Graterol Torres, F.-A.; Quirant Sánchez, B.; Marco Rusiñol, H.; Navarro Díaz, M.I.; Ara del Rey, J.; Martínez Cáceres, E.M. Immunological Pattern in IgA Nephropathy. Int. J. Mol. Sci. 2020, 21, 1389. https://doi.org/10.3390/ijms21041389
Esteve Cols C, Graterol Torres F-A, Quirant Sánchez B, Marco Rusiñol H, Navarro Díaz MI, Ara del Rey J, Martínez Cáceres EM. Immunological Pattern in IgA Nephropathy. International Journal of Molecular Sciences. 2020; 21(4):1389. https://doi.org/10.3390/ijms21041389
Chicago/Turabian StyleEsteve Cols, Clara, Freddzia-Amanda Graterol Torres, Bibiana Quirant Sánchez, Helena Marco Rusiñol, Maruja Isabel Navarro Díaz, Jordi Ara del Rey, and Eva Mª Martínez Cáceres. 2020. "Immunological Pattern in IgA Nephropathy" International Journal of Molecular Sciences 21, no. 4: 1389. https://doi.org/10.3390/ijms21041389
APA StyleEsteve Cols, C., Graterol Torres, F. -A., Quirant Sánchez, B., Marco Rusiñol, H., Navarro Díaz, M. I., Ara del Rey, J., & Martínez Cáceres, E. M. (2020). Immunological Pattern in IgA Nephropathy. International Journal of Molecular Sciences, 21(4), 1389. https://doi.org/10.3390/ijms21041389