Impact of Targeted Deletion of the Circadian Clock Gene Bmal1 in Excitatory Forebrain Neurons on Adult Neurogenesis and Olfactory Function

 ,
, 
Abstract
:1. Introduction
2. Results
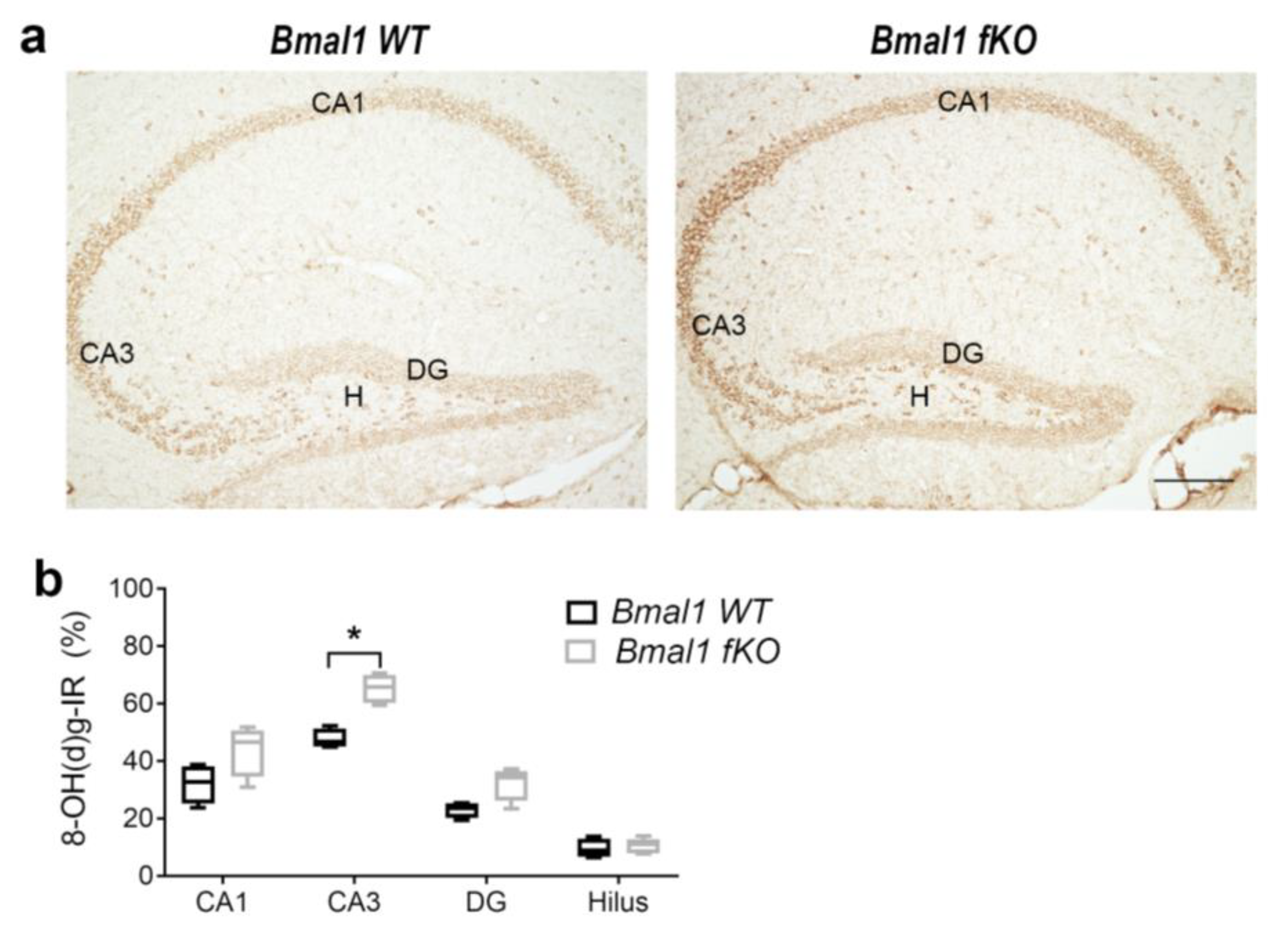
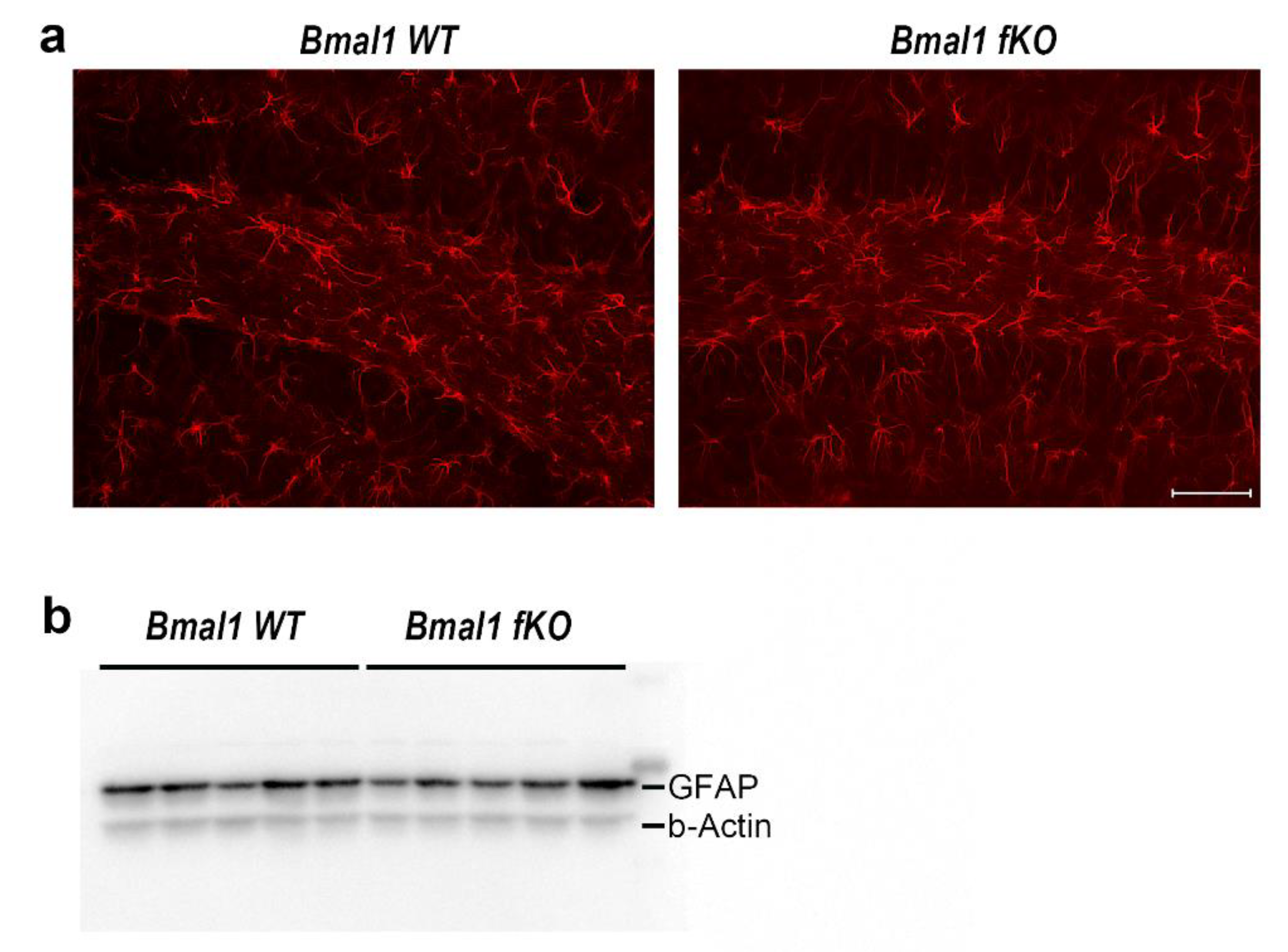
2.1. Forebrain Specific Bmal1 Deletion Leads to Subregional Increases in Oxidative Stress but not to Astrocyte Activation
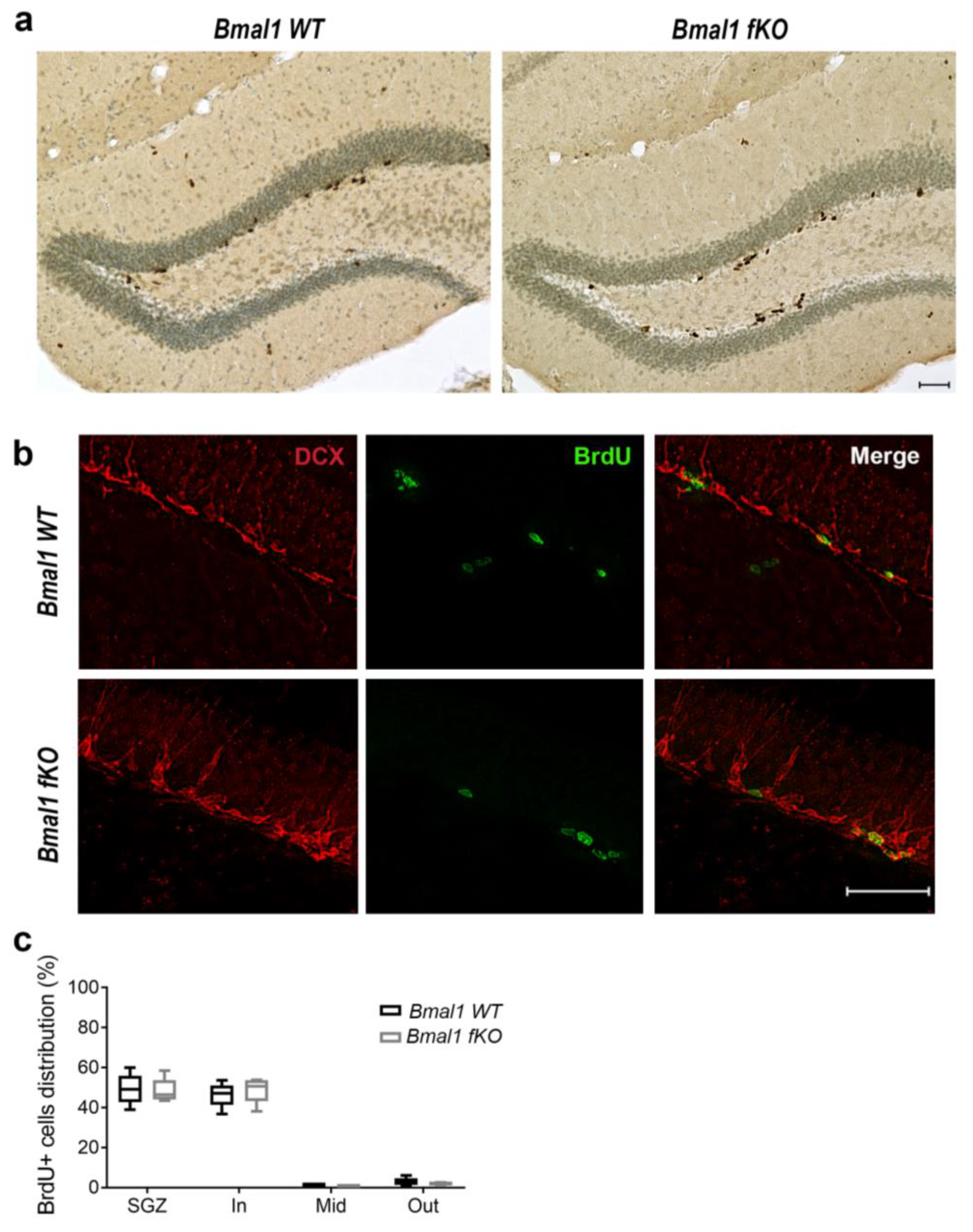
2.2. Forebrain Specific Bmal1 Deletion does not Affect Adult Neurogenesis in the Dentate Gyrus
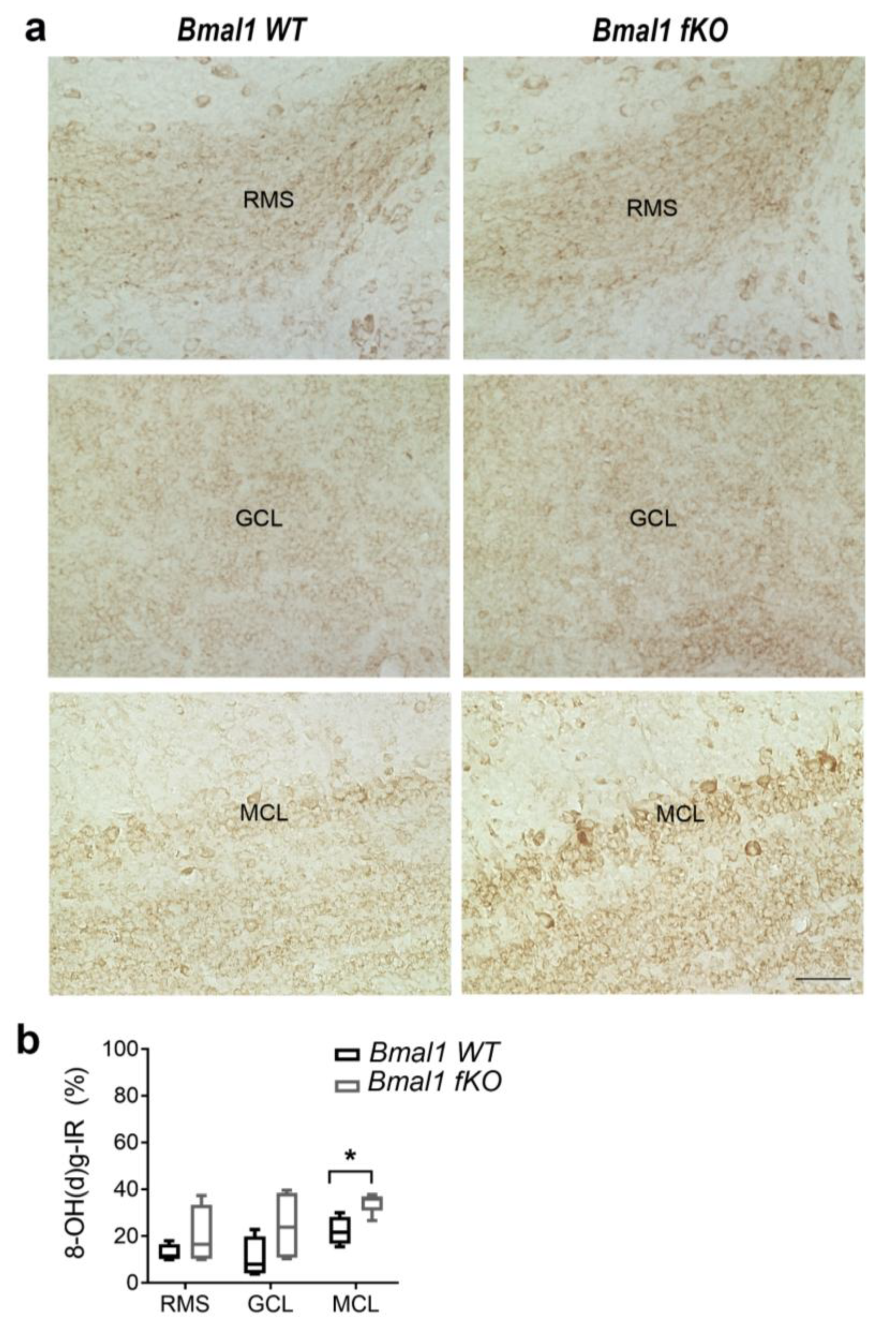
2.3. Forebrain Specific Bmal1 Deletion Affects Oxidative Stress in the Mitral Cell Layer (MCL) of the Olfactory Bulb (OB)
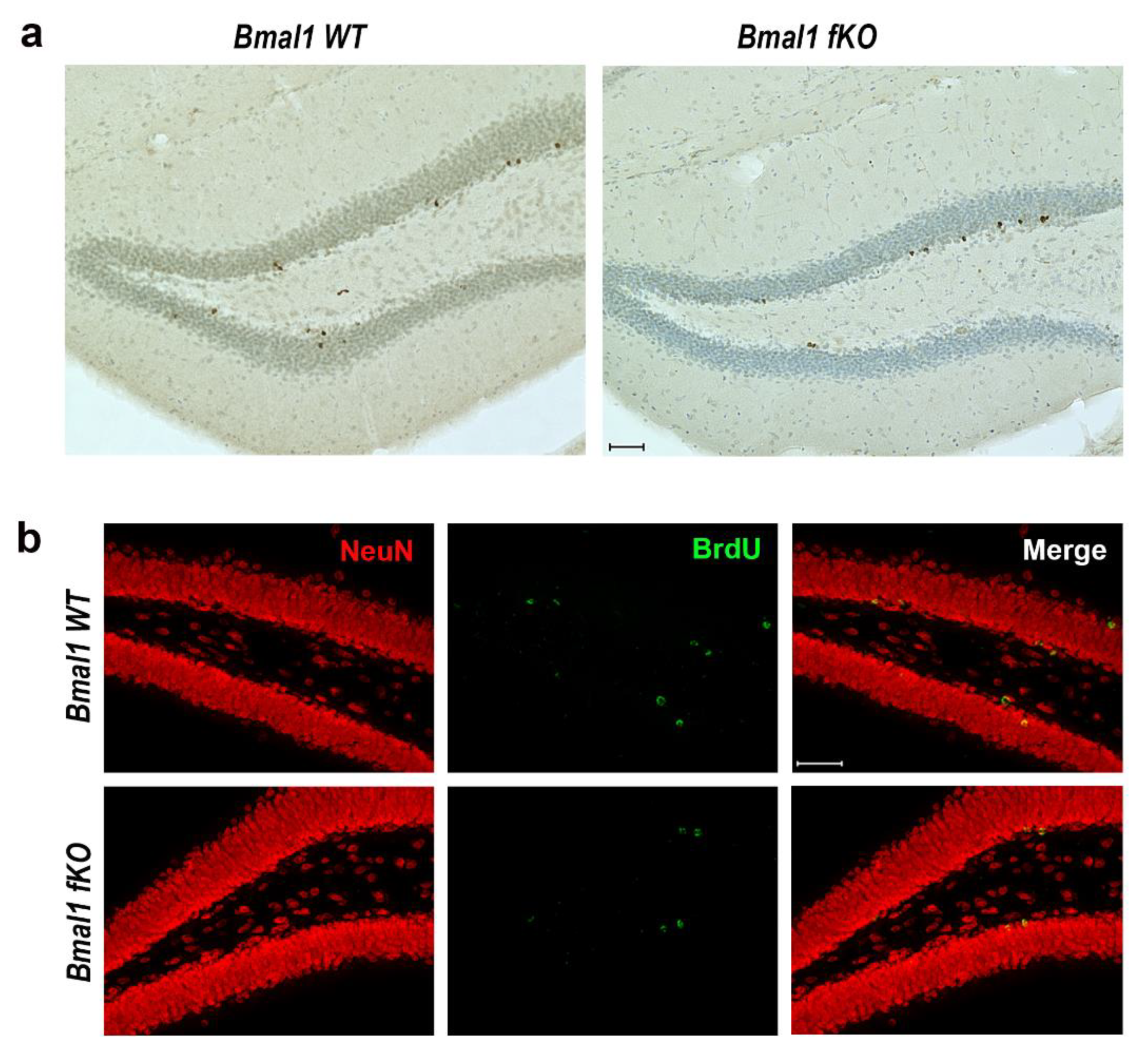
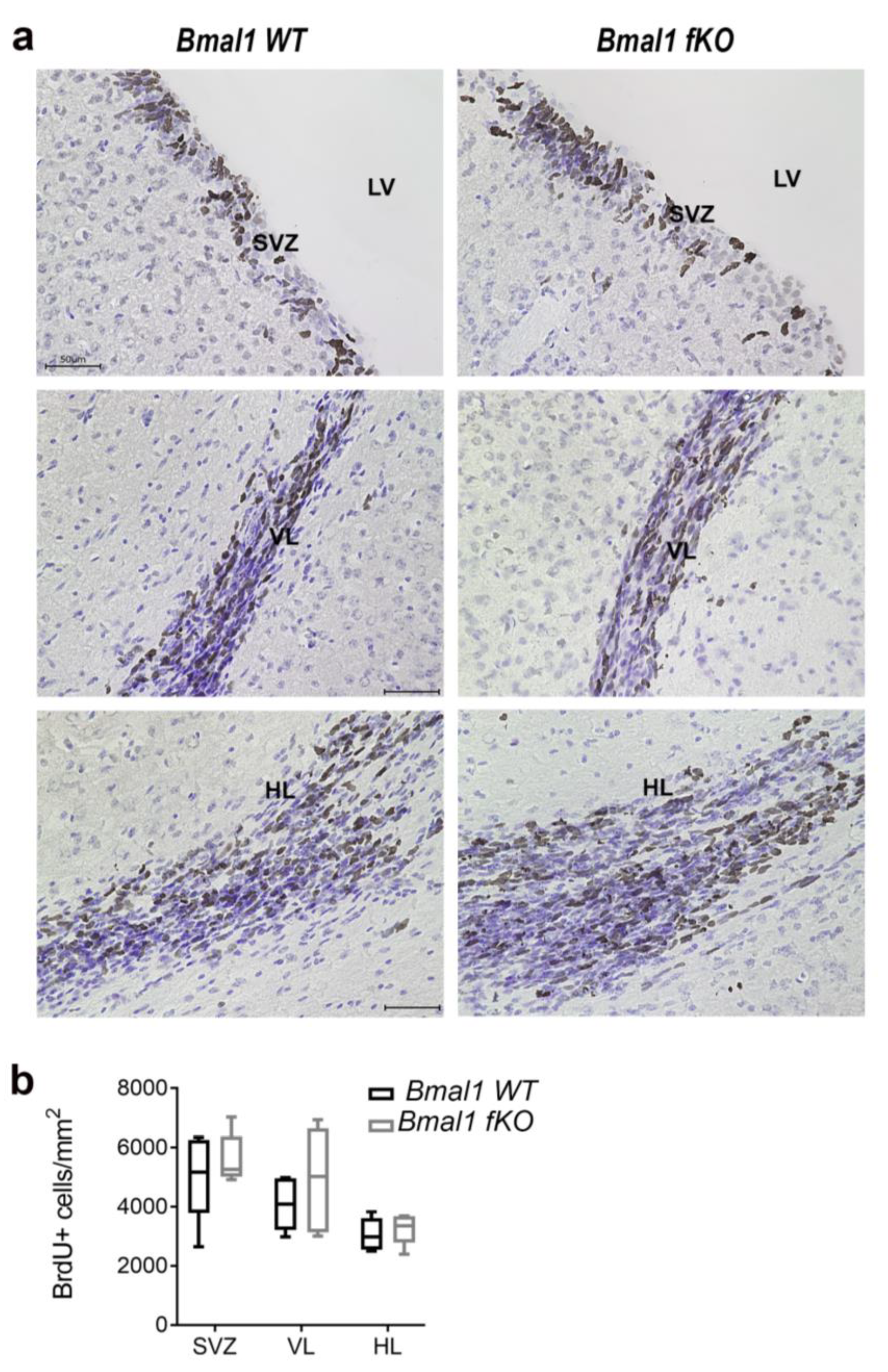
2.4. Forebrain Specific Bmal1 Deletion does not Affect Adult Neurogenesis in the Subventricular Zone and the Rostral Migratory Stream
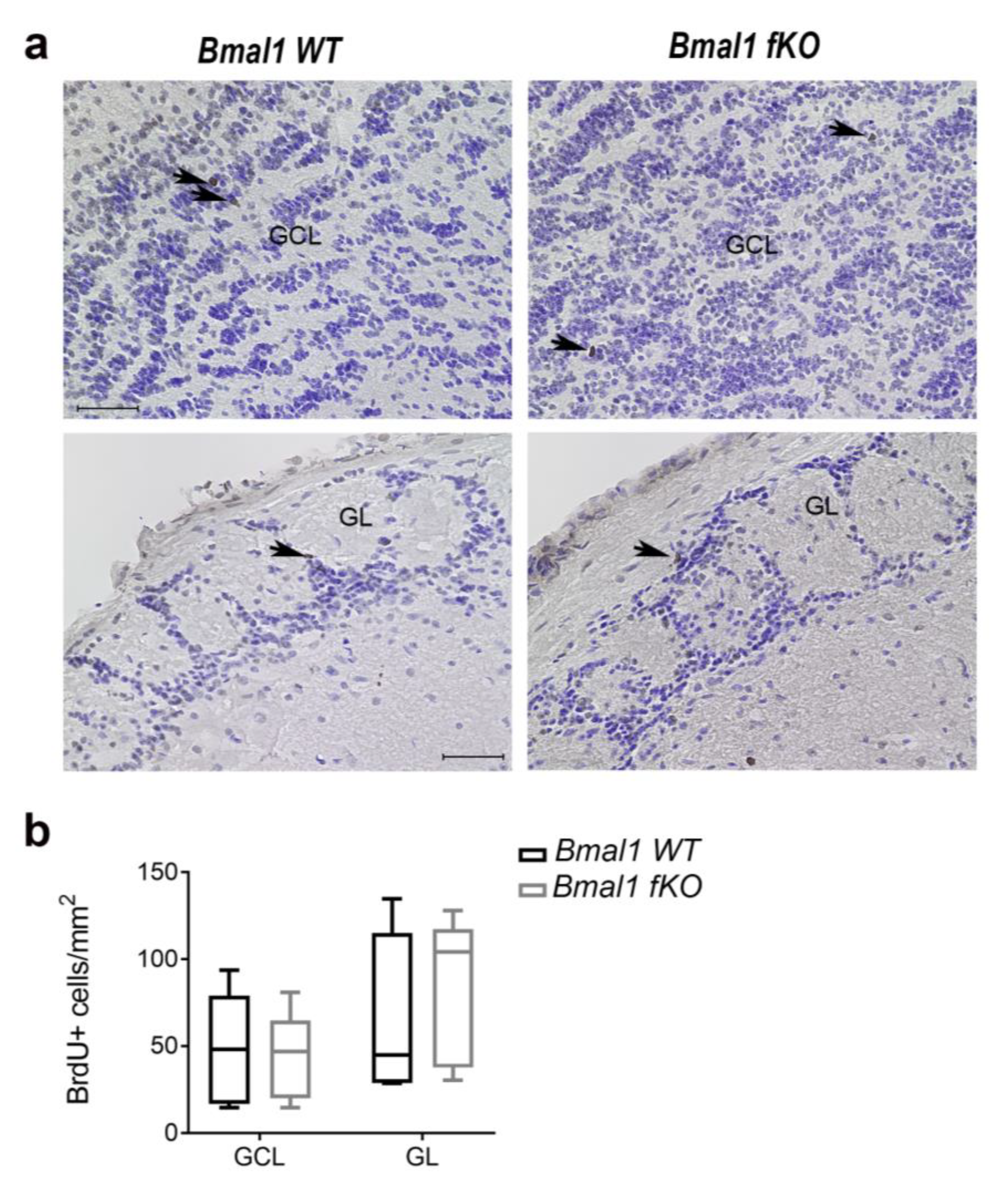
2.5. Forebrain Specific Bmal1 Deletion does not Affect Migration of NPCs to the OB
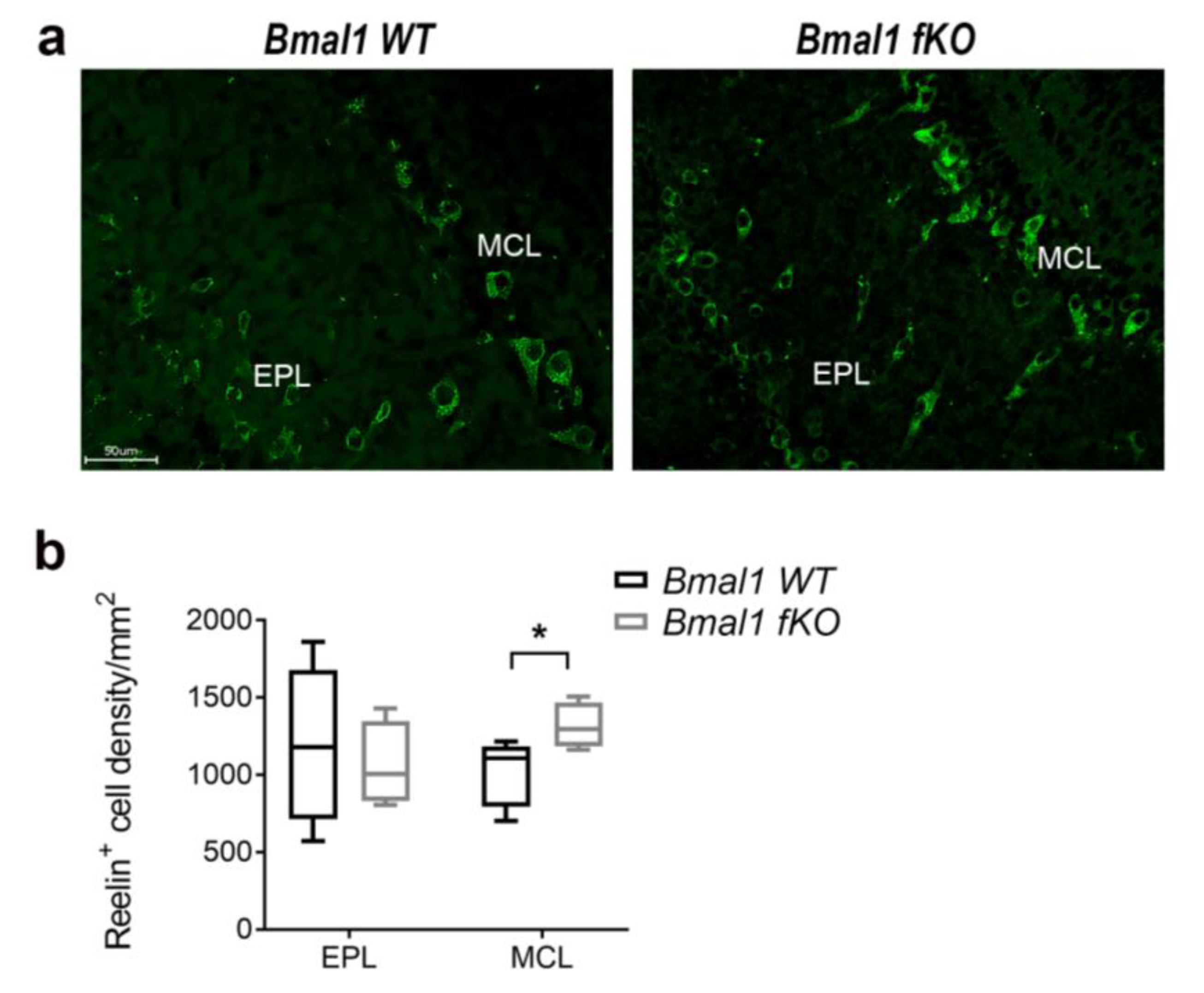
2.6. Forebrain Specific Bmal1 Deletion Affects Reelin Expression in OB
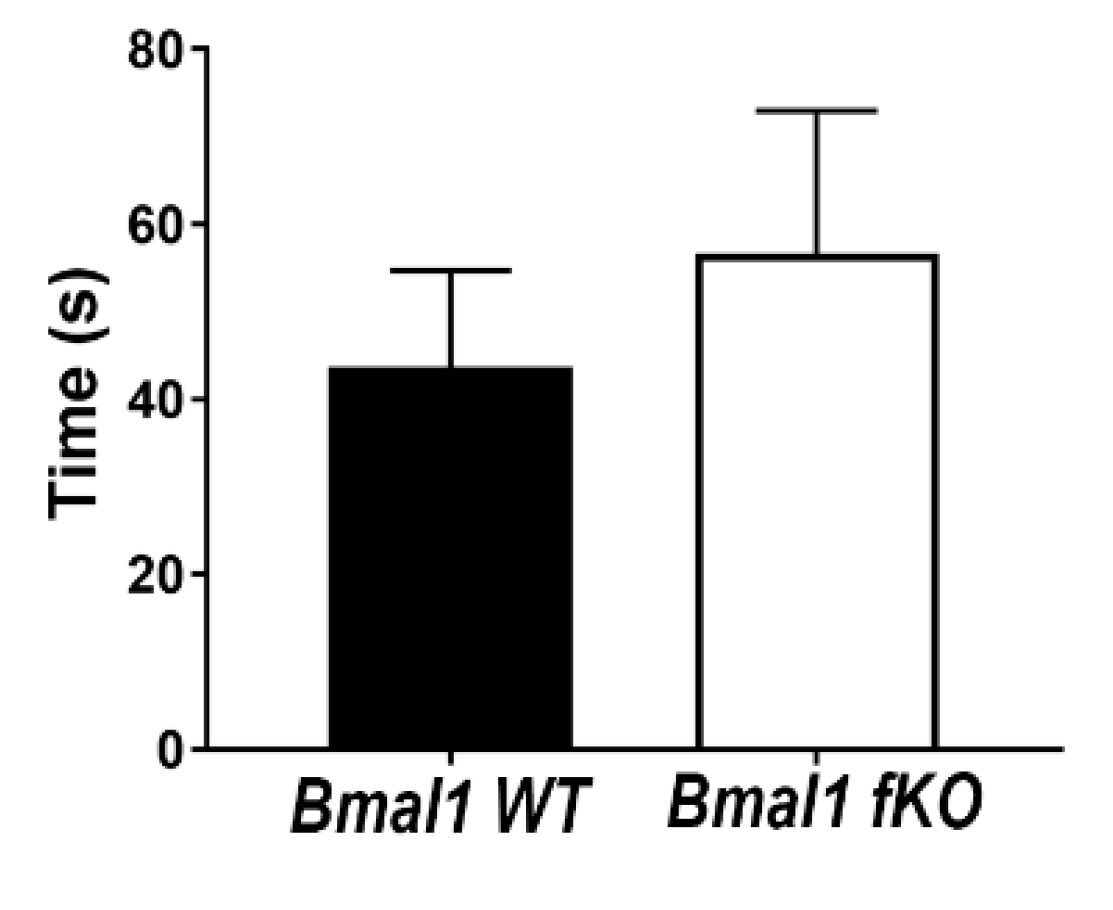
2.7. Olfactory Function Is Not Affected in Forebrain-Specific Bmal1-Deficient Mice
3. Discussion
4. Material and Methods
4.1. Experimental Animals
4.2. BrdU Assay
4.3. Tissue Processing
4.4. Immunohistochemistry
4.5. Immunofluorescence
4.6. Image Acquisition and Analysis
4.7. Immunoblot
4.8. Behavioral Assays
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Birky, T.L.; Bray, M.S. Understanding circadian gene function: Animal models of tissue-specific circadian disruption. IUBMB Life 2014, 66, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Khapre, R.V.; Patel, S.A.; Kondratova, A.A.; Chaudhary, A.; Velingkaar, N.; Antoch, M.P.; Kondratov, R.V. Metabolic clock generates nutrient anticipation rhythms in mTOR signaling. Aging (Albany NY) 2014, 6, 675–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, C.H.; Takahashi, J.S. Molecular components of the mammalian circadian clock. Hum. Mol. Genet. 2006, 15, R271–R277. [Google Scholar] [CrossRef]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Chen, L.; Grant, G.R.; Paschos, G.; Song, W.L.; Musiek, E.S.; Lee, V.; McLoughlin, S.C.; Grosser, T.; Cotsarelis, G.; et al. Timing of expression of the core clock gene Bmal1 influences its effects on aging and survival. Sci. Trans. Med. 2016, 8, 324ra16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, M.; Wicht, H.; von Gall, C.; Korf, H.W. Owls and larks in mice. Front. Neurol. 2015, 6, 101. [Google Scholar] [CrossRef] [Green Version]
- Kondratova, A.A.; Dubrovsky, Y.V.; Antoch, M.P.; Kondratov, R.V. Circadian clock proteins control adaptation to novel environment and memory formation. Aging (Albany NY) 2010, 2, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Kondratov, R.V.; Kondratova, A.A.; Gorbacheva, V.Y.; Vykhovanets, O.V.; Antoch, M.P. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006, 20, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.A.; Schwarz-Herzke, B.; Stahr, A.; Prozorovski, T.; Aktas, O.; von Gall, C. Premature aging of the hippocampal neurogenic niche in adult Bmal1-deficient mice. Aging (Albany NY) 2015, 7, 435–449. [Google Scholar] [CrossRef]
- Ali, A.A.H.; Schwarz-Herzke, B.; Mir, S.; Sahlender, B.; Victor, M.; Gorg, B.; Schmuck, M.; Dach, K.; Fritsche, E.; Kremer, A.; et al. Deficiency of the clock gene Bmal1 affects neural progenitor cell migration. Brain Struct. Funct. 2019, 224, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Snider, K.H.; Dziema, H.; Aten, S.; Loeser, J.; Norona, F.E.; Hoyt, K.; Obrietan, K. Modulation of learning and memory by the targeted deletion of the circadian clock gene Bmal1 in forebrain circuits. Behav. Brain Res. 2016, 308, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Aimone, J.B.; Deng, W.; Gage, F.H. Resolving new memories: A critical look at the dentate gyrus, adult neurogenesis, and pattern separation. Neuron 2011, 70, 589–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shors, T.J.; Miesegaes, G.; Beylin, A.; Zhao, M.; Rydel, T.; Gould, E. Neurogenesis in the adult is involved in the formation of trace memories. Nature 2001, 410, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.L.; Song, H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aimone, J.B.; Wiles, J.; Gage, F.H. Computational influence of adult neurogenesis on memory encoding. Neuron 2009, 61, 187–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, J.S.; Hong, N.S.; McDonald, R.J.; Wojtowicz, J.M. A role for adult neurogenesis in spatial long-term memory. Neuroscience 2005, 130, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Anacker, C.; Hen, R. Adult hippocampal neurogenesis and cognitive flexibility-linking memory and mood. Nat. Rev. Neurosci. 2017, 18, 335–346. [Google Scholar] [CrossRef]
- Gritti, A.; Bonfanti, L.; Doetsch, F.; Caille, I.; Alvarez-Buylla, A.; Lim, D.A.; Galli, R.; Verdugo, J.M.; Herrera, D.G.; Vescovi, A.L. Multipotent neural stem cells reside into the rostral extension and olfactory bulb of adult rodents. J. Neurosci. 2002, 22, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Doetsch, F.; Alvarez-Buylla, A. Network of tangential pathways for neuronal migration in adult mammalian brain. Proc. Natl. Acad. Sci. USA 1996, 93, 14895–14900. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.M.; Qu, T.; Kriho, V.; Lacor, P.; Smalheiser, N.; Pappas, G.D.; Guidotti, A.; Costa, E.; Sugaya, K. Reelin function in neural stem cell biology. Proc. Natl. Acad. Sci. USA 2002, 99, 4020–4025. [Google Scholar] [CrossRef] [Green Version]
- Panzanelli, P.; Bardy, C.; Nissant, A.; Pallotto, M.; Sassoe-Pognetto, M.; Lledo, P.M.; Fritschy, J.M. Early synapse formation in developing interneurons of the adult olfactory bulb. J. Neurosci. 2009, 29, 15039–15052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livneh, Y.; Adam, Y.; Mizrahi, A. Odor processing by adult-born neurons. Neuron 2014, 81, 1097–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellwig, S.; Hack, I.; Zucker, B.; Brunne, B.; Junghans, D. Reelin together with ApoER2 regulates interneuron migration in the olfactory bulb. PLoS ONE 2012, 7, e50646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama-Yamamoto, A.; Yamamoto, T.; Miki, A.; Terashima, T. Changes in reelin expression in the mouse olfactory bulb after chemical lesion to the olfactory epithelium. Eur. J. Neurosci. 2005, 21, 2586–2592. [Google Scholar] [CrossRef] [PubMed]
- Musiek, E.S.; Lim, M.M.; Yang, G.; Bauer, A.Q.; Qi, L.; Lee, Y.; Roh, J.H.; Ortiz-Gonzalez, X.; Dearborn, J.T.; Culver, J.P.; et al. Circadian clock proteins regulate neuronal redox homeostasis and neurodegeneration. J. Clin. Investig. 2013, 123, 5389–5400. [Google Scholar] [CrossRef]
- Pfeffer, M.; Muller, C.M.; Mordel, J.; Meissl, H.; Ansari, N.; Deller, T.; Korf, H.W.; von Gall, C. The mammalian molecular clockwork controls rhythmic expression of its own input pathway components. J. Neurosci. 2009, 29, 6114–6123. [Google Scholar] [CrossRef]
- Maury, E. Off the Clock: From Circadian Disruption to Metabolic Disease. Int. J. Mol. Sci. 2019, 20, 1597. [Google Scholar] [CrossRef] [Green Version]
- Bayer, K.U.; Lohler, J.; Schulman, H.; Harbers, K. Developmental expression of the CaM kinase II isoforms: Ubiquitous gamma- and delta-CaM kinase II are the early isoforms and most abundant in the developing nervous system. Mol. Brain Res. 1999, 70, 147–154. [Google Scholar] [CrossRef]
- Deisseroth, K.; Singla, S.; Toda, H.; Monje, M.; Palmer, T.D.; Malenka, R.C. Excitation-neurogenesis coupling in adult neural stem/progenitor cells. Neuron 2004, 42, 535–552. [Google Scholar] [CrossRef] [Green Version]
- Gould, E.; Tanapat, P.; Rydel, T.; Hastings, N. Regulation of hippocampal neurogenesis in adulthood. Biol. Psychiatry 2000, 48, 715–720. [Google Scholar] [CrossRef]
- Duncan, M.J.; Prochot, J.R.; Cook, D.H.; Tyler Smith, J.; Franklin, K.M. Influence of aging on Bmal1 and Per2 expression in extra-SCN oscillators in hamster brain. Brain Res. 2013, 1491, 44–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesner, R.P. Behavioral functions of the CA3 subregion of the hippocampus. Learn. Mem. 2007, 14, 771–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evola, M.; Hall, A.; Wall, T.; Young, A.; Grammas, P. Oxidative stress impairs learning and memory in apoE knockout mice. Pharmacol. Biochem. Behav. 2010, 96, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Qvartskhava, N.; Lang, P.A.; Gorg, B.; Pozdeev, V.I.; Ortiz, M.P.; Lang, K.S.; Bidmon, H.J.; Lang, E.; Leibrock, C.B.; Herebian, D.; et al. Hyperammonemia in gene-targeted mice lacking functional hepatic glutamine synthetase. Proc. Natl. Acad. Sci. USA 2015, 112, 5521–5526. [Google Scholar] [CrossRef] [Green Version]
- Haussinger, D.; Sies, H. Hepatic encephalopathy: Clinical aspects and pathogenetic concept. Arch. Biochem. Biophys. 2013, 536, 97–100. [Google Scholar] [CrossRef]
- Blanchart, A.; De Carlos, J.A.; Lopez-Mascaraque, L. Time frame of mitral cell development in the mice olfactory bulb. J. Comp. Neurol. 2006, 496, 529–543. [Google Scholar] [CrossRef]
- Xiong, W.; Chen, W.R. Dynamic gating of spike propagation in the mitral cell lateral dendrites. Neuron 2002, 34, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Whitman, M.C.; Greer, C.A. Adult neurogenesis and the olfactory system. Prog. Neurobiol. 2009, 89, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Pujadas, L.; Gruart, A.; Bosch, C.; Delgado, L.; Teixeira, C.M.; Rossi, D.; de Lecea, L.; Martinez, A.; Delgado-Garcia, J.M.; Soriano, E. Reelin regulates postnatal neurogenesis and enhances spine hypertrophy and long-term potentiation. J. Neurosci. 2010, 30, 4636–4649. [Google Scholar] [CrossRef] [Green Version]
- Rideau Batista Novais, A.; Guiramand, J.; Cohen-Solal, C.; Crouzin, N.; de Jesus Ferreira, M.C.; Vignes, M.; Barbanel, G.; Cambonie, G. N-acetyl-cysteine prevents pyramidal cell disarray and reelin-immunoreactive neuron deficiency in CA3 after prenatal immune challenge in rats. Pediatr. Res. 2013, 73, 750–755. [Google Scholar] [CrossRef] [Green Version]
- Forero, D.A.; Casadesus, G.; Perry, G.; Arboleda, H. Synaptic dysfunction and oxidative stress in Alzheimer’s disease: Emerging mechanisms. J. Cell Mol. Med. 2006, 10, 796–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granados-Fuentes, D.; Ben-Josef, G.; Perry, G.; Wilson, D.A.; Sullivan-Wilson, A.; Herzog, E.D. Daily rhythms in olfactory discrimination depend on clock genes but not the suprachiasmatic nucleus. J. Biol. Rhythm. 2011, 26, 552–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portaluppi, F.; Smolensky, M.H.; Touitou, Y. Ethics and methods for biological rhythm research on animals and human beings. Chronobiol. Int. 2010, 27, 1911–1929. [Google Scholar] [CrossRef] [PubMed]
- Casanova, E.; Fehsenfeld, S.; Mantamadiotis, T.; Lemberger, T.; Greiner, E.; Stewart, A.F.; Schutz, G. A CamKIIalpha iCre BAC allows brain-specific gene inactivation. Genesis 2001, 31, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Gheusi, G.; Cremer, H.; McLean, H.; Chazal, G.; Vincent, J.; Lledo, P. Importance of newly generated neurons in the adult olfactory bulb for odor discrimination. Proc. Natl. Acad. Sci. USA 2000, 97, 1823–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicht, H.; Korf, H.W.; Ackermann, H.; Ekhart, D.; Fischer, C.; Pfeffer, M. Chronotypes and rhythm stability in mice. Chronobiol. Int. 2014, 31, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.; Rotermund, N.; Grzelka, K.; Benz, J.; Lohr, C.; Hirnet, D. Adenosine A1 Receptor-Mediated Attenuation of Reciprocal Dendro-Dendritic Inhibition in the Mouse Olfactory Bulb. Front. Cell Neurosci. 2017, 11, 435. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Crawley, J.N. Simple behavioral assessment of mouse olfaction. Curr. Protoc. Neurosci. 2009, 8, 24. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Total OB Volume (µm3) | GL (µm) | EPL (µm) | MCL (µm) | IPL (µm) |
|---|---|---|---|---|---|
| Bmal1 WT | 10.2 + 0.52 | 100.3 + 10.98 | 161.6 + 2.7 | 29.07 + 1.5 | 32.08 + 1.5 |
| Bmal1 fKO | 10.04 + 0.42 | 118.3 + 19.8 | 165.5 + 19.6 | 34.28 + 3.9 | 35.3 + 4.5 |
| p value (t-test) | 0.8 | 0.4 | 0.8 | 0.2 | 0.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.A.H.; Tundo-Lavalle, F.; Hassan, S.A.; Pfeffer, M.; Stahr, A.; von Gall, C. Impact of Targeted Deletion of the Circadian Clock Gene Bmal1 in Excitatory Forebrain Neurons on Adult Neurogenesis and Olfactory Function. Int. J. Mol. Sci. 2020, 21, 1394. https://doi.org/10.3390/ijms21041394
Ali AAH, Tundo-Lavalle F, Hassan SA, Pfeffer M, Stahr A, von Gall C. Impact of Targeted Deletion of the Circadian Clock Gene Bmal1 in Excitatory Forebrain Neurons on Adult Neurogenesis and Olfactory Function. International Journal of Molecular Sciences. 2020; 21(4):1394. https://doi.org/10.3390/ijms21041394
Chicago/Turabian StyleAli, Amira A. H., Federica Tundo-Lavalle, Soha A. Hassan, Martina Pfeffer, Anna Stahr, and Charlotte von Gall. 2020. "Impact of Targeted Deletion of the Circadian Clock Gene Bmal1 in Excitatory Forebrain Neurons on Adult Neurogenesis and Olfactory Function" International Journal of Molecular Sciences 21, no. 4: 1394. https://doi.org/10.3390/ijms21041394
APA StyleAli, A. A. H., Tundo-Lavalle, F., Hassan, S. A., Pfeffer, M., Stahr, A., & von Gall, C. (2020). Impact of Targeted Deletion of the Circadian Clock Gene Bmal1 in Excitatory Forebrain Neurons on Adult Neurogenesis and Olfactory Function. International Journal of Molecular Sciences, 21(4), 1394. https://doi.org/10.3390/ijms21041394