Calcium-Involved Action of Phytochemicals: Carotenoids and Monoterpenes in the Brain


Abstract
:1. Introduction
2. The Effect of Terpenes and Their Derivatives on Calcium-Engaged Signaling
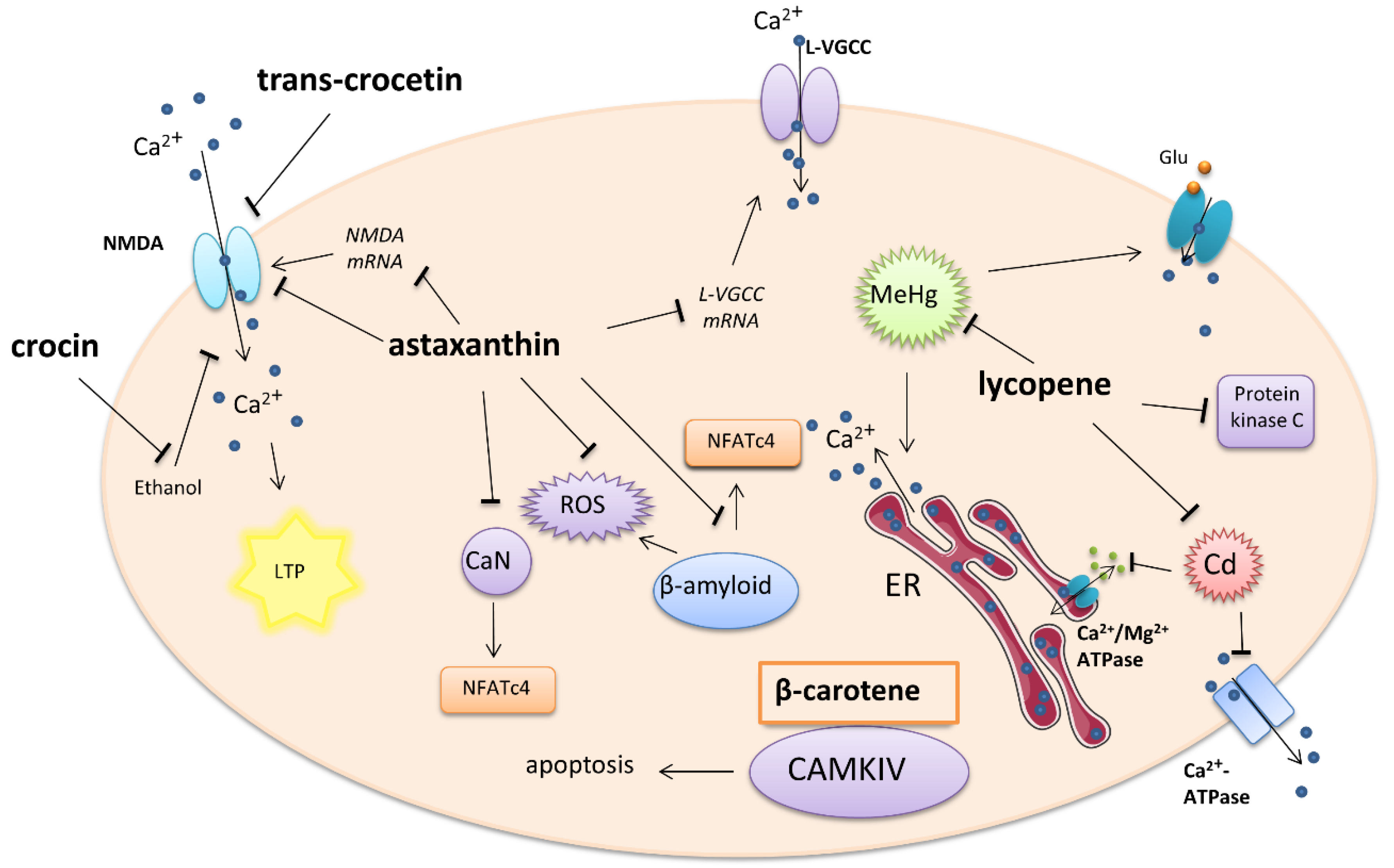
2.1. Carotenoids
2.1.1. Astaxanthin
2.1.2. Lycopene
2.1.3. β-Carotene
2.1.4. Crocin and Crocetin
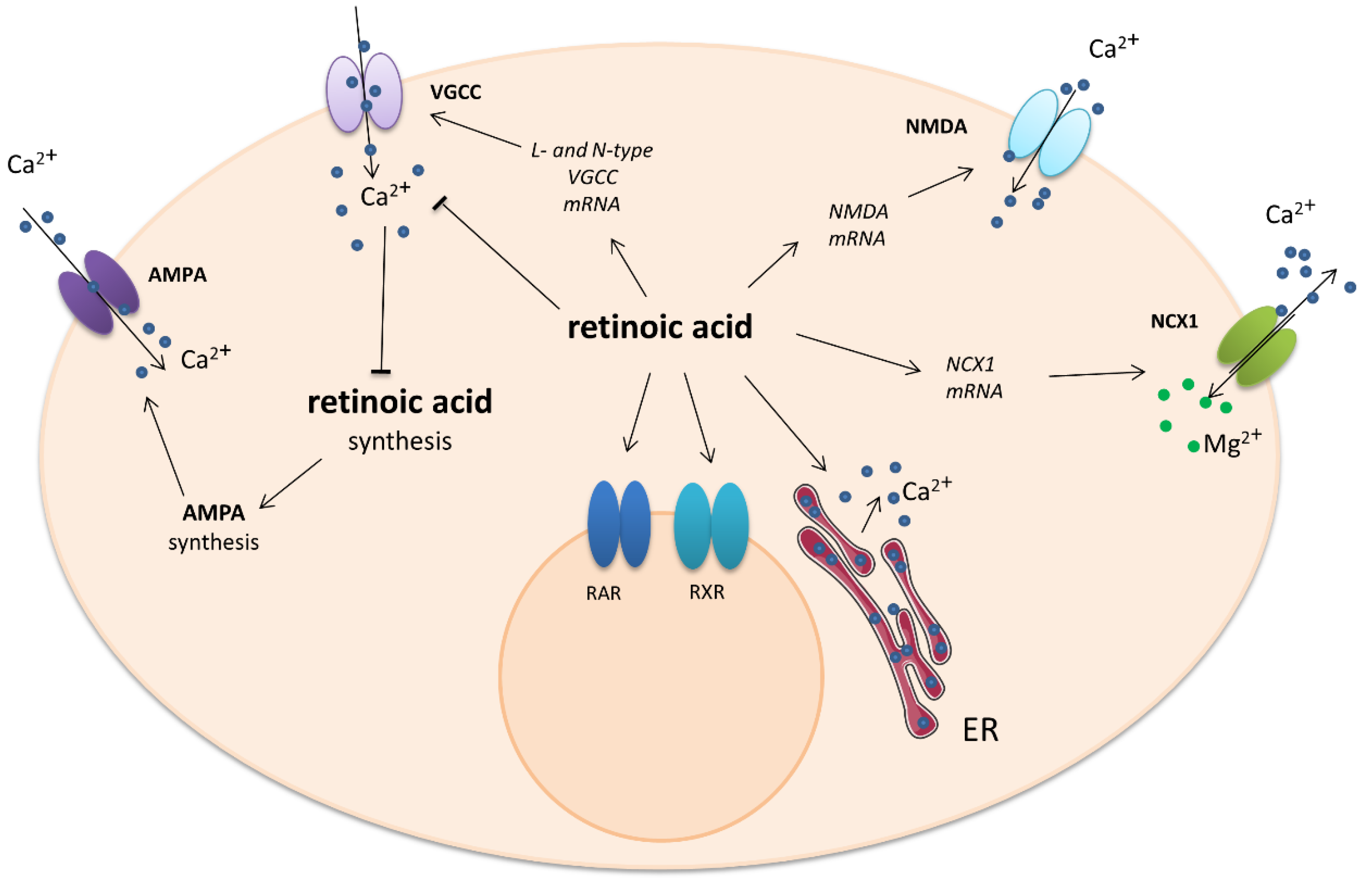
2.2. Retinoic Acid (Diterpene)
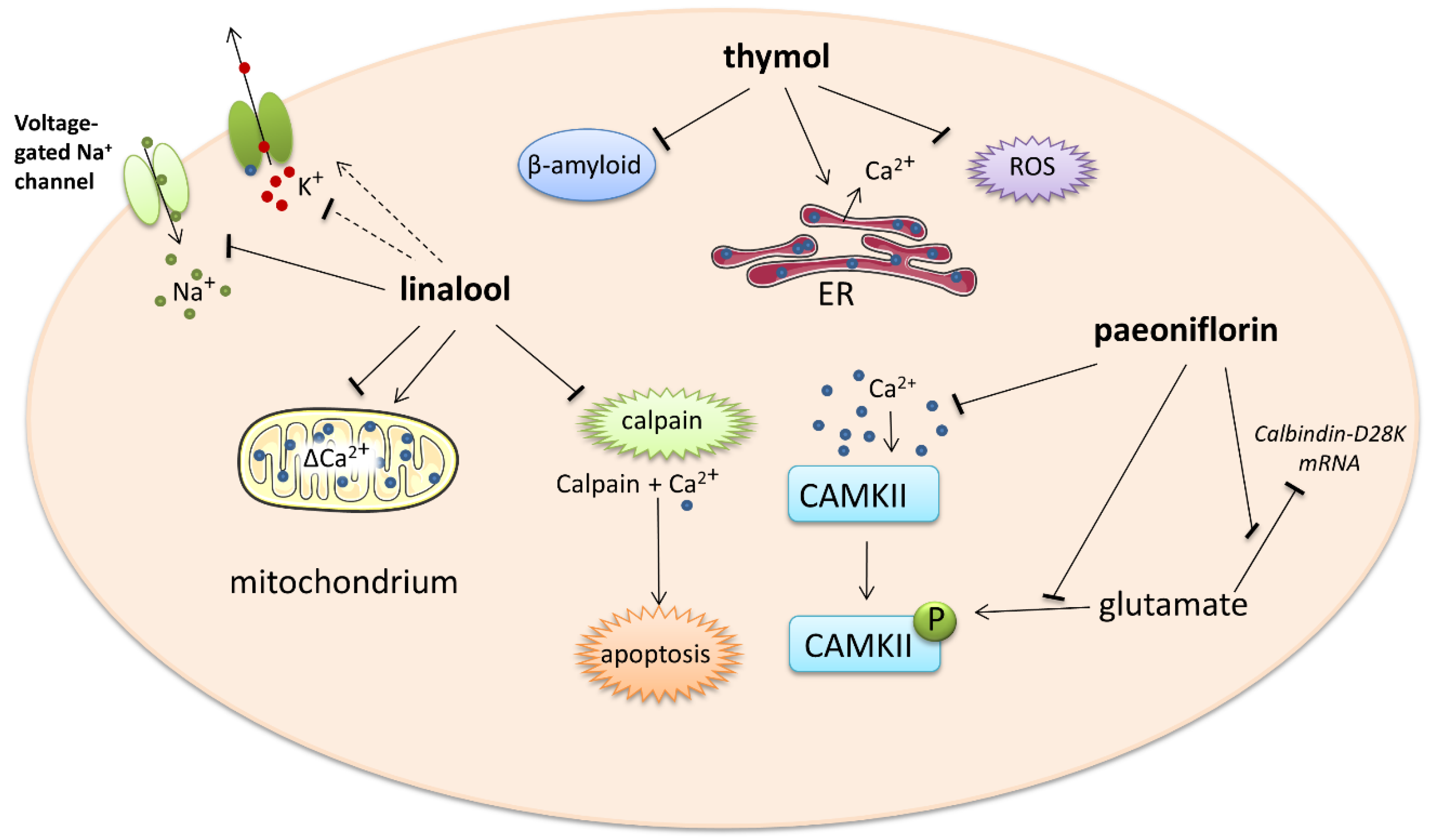
2.3. Monoterpenes
2.3.1. Linalool
2.3.2. Thymol
2.3.3. Paeoniflorin
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| NMDAR | N-methyl-D-aspartate receptor |
| AMPAR | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor |
| VGCC | Voltage-gated calcium channel |
| NCX | Sodium–Calcium Exchanger |
| RyR | Ryanodine receptor |
| PKC | Protein kinase C |
| CAMK | Calcium/calmodulin-dependent kinase type II or IV |
| AXT | Astaxanthin |
| RA | Retinoic acid |
| ATRA | All-trans-retinoic acids |
| RAR | Retinoic acid receptor |
| RXR | Retinoid X receptors |
References
- Charu, G.; Dhan, P. Phytonutrients as therapeutic agent. J. Complement. Integr. Med. 2014, 11, 151–156. [Google Scholar] [CrossRef]
- Liu, R.H. Health-promoting components of fruits and vegetables in the diet. Adv. Nutr. 2013, 4, 384–392. [Google Scholar] [CrossRef]
- Hussain, G.; Rasul, A.; Anwar, H.; Aziz, N.; Razzaq, A.; Wei, W.; Ali, M.; Li, J.; Li, X. Role of Plant Derived Alkaloids and Their Mechanism in Neurodegenerative Disorders. Int. J. Biol. Sci. 2018, 14, 341–357. [Google Scholar] [CrossRef] [Green Version]
- Bhullar, K.S.; Rupasinghe, H.P. Polyphenols: Multipotent therapeutic agents in neurodegenerative diseases. Oxid. Med. Cell Longev. 2013. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Rebas, E.; Rzajew, J.; Radzik, T.; Zylinska, L. Neuroprotective Polyphenols: A Modulatory Action on Neurotransmitter Pathways. Curr. Neuropharmacol. 2020. [Google Scholar] [CrossRef]
- Zündorf, G.; Reiser, G. Calcium Dysregulation and Homeostasis of Neural Calcium in the Molecular Mechanisms of Neurodegenerative Diseases Provide Multiple Targets for Neuroprotection. Antioxid. Redox Signal. 2011, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Pchitskaya, E.; Popugaeva, E.; Bezprozvanny, I. Calcium signaling and molecular mechanisms underlying neurodegenerative diseases. Cell Calcium. 2018, 70, 87–94. [Google Scholar] [CrossRef]
- Guo, H.; Camargo, L.M.; Yeboah, F.; Digan, M.E.; Niu, H.; Pan, Y.; Reiling, S.; Soler-Llavina, G.; Weihofen, W.A.; Wang, H.R.; et al. A NMDA-receptor calcium influx assay sensitive to stimulation by glutamate and glycine/D-serine. Sci. Rep. 2017, 7, 11608. [Google Scholar] [CrossRef] [Green Version]
- König, N.; Poluch, S.; Estabel, J.; Durand, M.; Drian, M.J. Synaptic and Non-synaptic AMPA Receptors Permeable to Calcium. Jpn. J. Pharmacol. 2001, 86, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Gleichmann, M.; Mattson, M.P. Neuronal Calcium Homeostasis and Dysregulation. Antioxid. Redox Signal. 2011, 14, 1261–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, D.; Ankarcrona, M. Beyond the critical point: An overview of excitotoxicity, calcium overload and the downstream consequences. Neurosci. Lett. 2018, 663, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Brini, M.; Carafoli, E. Calcium Pumps in Health and Disease. Physiol. Rev. 2009, 89, 1341–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niswender, C.M.; Conn, J. Metabotropic Glutamate Receptors: Physiology, Pharmacology, and Disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [Green Version]
- Kania, E.; Roest, G.; Vervliet, T.; Parys, J.B.; Bultynck, G. IP3 Receptor-Mediated Calcium Signaling and Its Role in Autophagy in Cancer. Front. Oncol. 2017, 7, 140. [Google Scholar] [CrossRef] [Green Version]
- Armita, S. Transient Receptor Potential (TRP) Channels. Subcell Biochem. 2018, 87, 141–165. [Google Scholar] [CrossRef]
- Fairless, R.; Williams, S.K.; Diem, R. Calcium-Binding Proteins as Determinants of Central Nervous System Neuronal Vulnerability to Disease. Int. J. Mol. Sci. 2019, 20, 2146. [Google Scholar] [CrossRef] [Green Version]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytologist. 2018, 220, 655–658. [Google Scholar] [CrossRef]
- Tetali, S.D. Terpenes and isoprenoids: A wealth of compounds for global use. Planta 2019, 249, 1–8. [Google Scholar] [CrossRef]
- Francis, X.; Cunningham, E.G. Elucidation of the Pathway to Astaxanthin in the Flowers of Adonis Aestivalis. Plant Cell 2011, 23, 3055–3069. [Google Scholar] [CrossRef] [Green Version]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, R.K.; Varadaraj, B.G. Crocin: An Overview. Indo Glob. J. Pharm. Sci. 2011, 1, 281–286. [Google Scholar]
- Sarkar, A.K. Emerging Trends of Bioscience Research; Educreation Publishing: New Delhi, India, 2019. [Google Scholar]
- Gilbert, C. What is vitamin A and why do we need it? Community Eye Health J. 2013, 26, 65. [Google Scholar]
- Sharma, S.; Anand, N. Approaches to Design and Synthesis of Antiparasitic Drugs; Elsevier Science: Amsterdam, The Netherlands, 1997. [Google Scholar]
- Zhang, L.; Wei, W. Anti-inflammatory and immunoregulatory effects of paeoniflorin and total glucosides of paeony. Pharm. Ther. 2019. [Google Scholar] [CrossRef]
- Cho, K.S.; Shin, M.; Kim, S.; Lee, S.B. Recent advances in studies on the therapeutic potential of dietary in neurodegenerative diseases. Oxid. Med. Cell Longev. 2018. [Google Scholar] [CrossRef]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [Green Version]
- Sudhahar, V.; Fukai, T. Studies on Pediatric Disorders; Chapter: Antioxidant Supplementation and Therapies; Tsukahara, H., Kaneko, K., Eds.; Springer: New York, NY, USA, 2014; pp. 183–209. [Google Scholar]
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids Functionality, Sources, and Processing by Supercritical Technology: A Review. J. Chem. 2016. [Google Scholar] [CrossRef] [Green Version]
- Mohammadzadeh-Honarvar, N.; Saedisomeolia, A.; Abdolahi, M.; Shayeganrad, A.; Taheri-Sangsari, G.; Hassanzadeh-Rad, B. Molecular Anti-inflammatory Mechanisms of Retinoids and Carotenoids in Alzheimer’s Disease: A Review of Current Evidence. J. Mol. Neurosci. 2017, 61, 289–304. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H. Multiple Mechanisms of Anti-Cancer Effects Exerted by Astaxanthin. Mar. Drugs 2015, 13, 4310–4330. [Google Scholar] [CrossRef] [Green Version]
- Grimmig, B.; Kim, S.H.; Nash, K.; Bickford, P.C.; Douglas, S.R. Neuroprotective mechanisms of astaxanthin: A potential therapeutic role in preserving cognitive function in age and neurodegeneration. GeroScience 2017, 39, 19–32. [Google Scholar] [CrossRef]
- Wu, T.H.; Liao, J.H.; Hou, W.C.; Huang, F.Y.; Maher, T.J.; Hu, C.C. Astaxanthin protects against oxidative stress and calcium-induced porcine lens protein degradation. J. Agric. Food Chem. 2006, 54, 2418–2423. [Google Scholar] [CrossRef] [PubMed]
- Altunrende, M.E.; Gezen-Ak, D.; Atasoy, I.L.; Candas, E.; Dursun, E. The Role of Astaxanthin on Transcriptional Regulation of NMDA receptors Voltage Sensitive Calcium Channels and Calcium Binding Proteins in Primary Cortical Neurons. Arch. Neuropsychiatry 2018, 55, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.J.; Yuan, H.; Kang, J.Q.; Tan, F.C.K.; Traynelis, S.F.; Low, C.M. Distinct roles of GRIN2A and GRIN2B variants in neurological conditions. F1000Research 2019, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.; Sharma, D.; Sharma, M.; Sharma, N.; Bidve, P.; Prajapati, N.; Kalia, K.; Tiwari, V. Astaxanthin ameliorates behavioral and biochemical alterations in in-vitro and in-vivo model of neuropathic pain. Neurosci. Lett. 2018, 674, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Zhang, X.; Huang, B.; Zhu, Y.; Chen, X. Astaxanthin suppresses MPP(+)-induced oxidative damage in PC12 cells through a Sp1/NR1 signaling pathway. Mar. Drugs 2013, 11, 1019–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cline, E.N.; Bicca, M.A.; Viola, K.L.; Klein, W.L. The Amyloid-Oligomer Hypothesis: Beginning of the Third Decade. J. Alzheimer’s Dis. 2018, 64, S567–S610. [Google Scholar] [CrossRef] [Green Version]
- Lobos, P.; Bruna, B.; Cordova, A.; Barattini, P.; Galáz, J.L.; Adasme, T.; Hidalgo, C.; Muñoz, P.; Paula-Lima, A. Astaxanthin Protects Primary Hippocampal Neurons against Noxious Effects of Aβ-Oligomers. Neural. Plast. 2016. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.Y.; Lu, C.W.; Wang, S.J. Astaxanthin inhibits glutamate release in rat cerebral cortex nerve terminals via suppression of voltage-dependent Ca(2+) entry and mitogen-activated protein kinase signaling pathway. J. Agric. Food Chem. 2010, 58, 8271–8278. [Google Scholar] [CrossRef]
- Momeni, H.R. Role of Calpain in Apoptosis. Cell J. 2011, 13, 65–72. [Google Scholar]
- Yildiz-Unal, A.; Korulu, S.; Karabay, A. Neuroprotective strategies against calpain-mediated neurodegeneration. Neuropsychiatr. Dis. Treat. 2015, 11, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Reese, C.L.; Taglialatela, G. A Role for Calcineurin in Alzheimers Disease. Curr. Neuropharmacol. 2011, 9, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.S.; Chang, C.L.; Lai, G.H. Reactive oxygen species scavenging activities in a chemiluminescence model and neuroprotection in rat pheochromocytoma cells by astaxanthin, beta-carotene, and canthaxanthin. Kaohsiung J. Med. Sci. 2013, 29, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Przybylska, S. Lycopene—A bioactive carotenoid offering multiple health benefits: A review. IJFST 2019. [Google Scholar] [CrossRef]
- Lu, C.W.; Hung, C.F.; Jean, W.H.; Lin, T.Y.; Huang, S.K.; Wang, S.J. Lycopene depresses glutamate release through inhibition of voltage-dependent Ca2+ entry and protein kinase C in rat cerebrocortical nerve terminals. Can. J. Physiol. Pharmacol. 2018, 96, 479–484. [Google Scholar] [CrossRef] [Green Version]
- Roos, D.; Seeger, R.; Puntel, R.; Vargas Barbosa, N. Role of calcium and mitochondria in MeHg-mediated cytotoxicity. J. Biomed. Biotechnol. 2012, 2012, 8–10. [Google Scholar] [CrossRef] [Green Version]
- Qu, M.; Nan, X.; Gao, Z.; Guo, B.; Liu, B.; Chen, Z. Protective effects of lycopene against methylmercury-induced neurotoxicity in cultured rat cerebellar granule neurons. Brain Res. 2013, 1540, 92–102. [Google Scholar] [CrossRef]
- Zhang, F.; Xing, S.; Li, Z. Antagonistic effects of lycopene on cadmium-induced hippocampal dysfunctions in autophagy, calcium homeostatis and redox. Oncotarget 2017, 8, 44720–44731. [Google Scholar] [CrossRef]
- Silalahi, J. Anticancer and health protective properties of citrus fruit components. Asia Pac. J. Clin. Nutr. 2002, 11, 79–84. [Google Scholar] [CrossRef]
- Naz, H.; Islam, A.; Ahmad, F.; Hassan, M. Calcium/calmodulin-dependent protein kinase IV: A multifunctional enzyme and potential therapeutic target. Prog. Biophys. Mol. Biol. 2016, 121, 54–65. [Google Scholar] [CrossRef]
- Naz, H.; Khan, P.; Tarique, M.; Rahman, S.; Meena, A.; Ahamad, S.; Luqman, S.; Islam, A.; Ahmad, F.; Hassan, M.I. Binding studies and biological evaluation of β-carotene as a potential inhibitor of human calcium/calmodulin-dependent protein kinase IV. Int. J. Biol. Macromol. 2017, 96, 161–170. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Barrow, C. Crocin, a Mechanistic Treatise. In Bioactive Compounds from Plant Origin: Extraction, Applications, and Potential Health Benefits; CRC Press: New York, NY, USA, 2019. [Google Scholar]
- Yorgun, M.A.; Rashid, K.; Aslanidis, A.; Bresgen, C.; Dannhausen, K.; Langmann, T. Crocin, a plant-derived carotenoid, modulates microglial reactivity. Biochem. Biophys. Rep. 2017, 12, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Berger, F.; Hensel, A.; Nieber, K. Saffron extract and trans-crocetin inhibit glutamatergic synaptic transmission in rat cortical brain slices. Neuroscience 2011, 180, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Sugiura, M.; Shoyama, Y.; Saito, H. Crocin antagonizes ethanol inhibition of NMDA receptor-mediated responses in rat hippocampal neurons. Brain Res. 1998, 787, 132–138. [Google Scholar] [CrossRef]
- Lechtenberg, M.; Schepmann, D.; Niehues, M.; Hellenbrand, N.; Wünsch, B.; Hensel, A. Quality and functionality of saffron: Quality control, species assortment and affinity of extract and isolated saffron compounds to NMDA and sigma1 (sigma-1) receptors. Planta Med. 2008, 74, 764–772. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Li, R.; Chen, G. Transcriptional Factors Mediating Retinoic Acid Signals in the Control of Energy Metabolism. Int. J. Mol. Sci. 2015, 16, 14210–14244. [Google Scholar] [CrossRef] [Green Version]
- Napoli, J.L.; Race, K.R. Biogenesis of retinoic acid from beta-carotene. Differences between the metabolism of beta-carotene and retinal. J. Biol. Chem. 1988, 263, 17372–17387. [Google Scholar]
- Vesprini, N.D.; Dawson, T.F.; Yuan, Y.; Bruce, D.; Spencer, G.E. Retinoic acid affects calcium signaling in adult molluscan neurons. Neurophysiol 2015, 113, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Hudecova, S.; Stefanik, P.; Macejova, D.; Brtko, J.; Krizanova, O. Retinoic Acid Increased Expression of the Na+/Ca2+ Exchanger in the Heart and Brain. Gen. Physiol. Biophys. 2004, 23, 417–422. [Google Scholar]
- De Hoog, E.; Lukewich, M.K.; Spencer, G.E. Retinoic acid inhibits neuronal voltage-gated calcium channels. Cell Calcium. 2018, 72, 51–61. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, X.; Chen, L.; Wei, H.; Chen, J.; Li, T.J. The change in retinoic acid receptor signaling induced by prenatal marginal vitamin A deficiency and its effects on learning and memory. Nutr. Biochem. 2017, 47, 75–85. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhang, Z.; Hintze, M.; Chen, L. Decrease in Calcium Concentration Triggers Neuronal Retinoic Acid Synthesis during Homeostatic Synaptic Plasticity. J. Neurosci. 2011, 31, 17764–17771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatanparast, J.; Bazleh, S.; Janahmadi, M. The effects of linalool on the excitability of central neurons of snail Caucasotachea atrolabiata. Comp. Biochem. Physiol. 2017, 192 Pt C, 33–39. [Google Scholar] [CrossRef]
- Elisabetsky, E.; Brum, L.F.; Souza, D.O. Anticonvulsant properties of linalool in glutamate-related seizure models. Phytomedicine 1999, 6, 107–113. [Google Scholar] [CrossRef]
- Sabogal-Guáqueta, A.M.; Hobbie, F.; Keerthi, A.; Oun, A.; Kortholt, A.; Boddeke, E.; Dolga, A. Linalool attenuates oxidative stress and mitochondrial dysfunction mediated by glutamate and NMDA toxicity. Biomed. Pharmacother. 2019, 118, 109295. [Google Scholar] [CrossRef]
- Corasaniti, M.T.; Maiuolo, J.; Maida, S.; Fratto, V.; Navarra, M.; Russo, R.; Amantea, D.; Morrone, L.A.; Bagetta, G. Cell signaling pathways in the mechanisms of neuroprotection afforded by bergamot essential oil against NMDA-induced cell death in vitro. Br. J. Pharmacol. 2007, 151, 518–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, H.; Azimullah, S.; Meeran, M.F.N.; Ansari, S.A. Neuroprotective effects of thymol, a dietary monoterpene against dopaminergic neurodegeneration in rotenone induced rat model of Parkinson’s disease. Int. J. Mol. Sci. 2019, 20, 1538. [Google Scholar] [CrossRef] [Green Version]
- Asadbegi, M.; Yaghmaei, P.; Salehi, I.; Komaki, A.; Ebrahim-Habibi, A. Investigation of thymol effect on learning and memory impairment induced by intrahippocampal injection of amyloid beta peptide in high fat diet- fed rats. Metab. Brain Dis. 2017, 32, 827–839. [Google Scholar] [CrossRef]
- Shen, A.Y.; Huang, M.H.; Wang, T.S.; Wu, H.M.; Kang, Y.F.; Chen, C.L. Thymol evoked Ca+ mobilization and ion currents in pituitary GH3 cells. Nat. Prod. Commun. 2009, 4, 749–752. [Google Scholar]
- Kostyuk, P.G.; Belan, P.V.; Tepikin, A.V. Free calcium transients and oscillations in nerve cells. Exp. Brain. Res. 1991, 83, 459–464. [Google Scholar] [CrossRef]
- Zolfaghari, Z.; Vatanparast, J. Thymol provokes burst of action potentials in neurons of snail Caucasotachea atrolabiata. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 228, 108654. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Zhong, X.M.; Feng, H.R.; Pan, A.J.; Li, Z.Y.; Huang, Z. Protective Effects of Paeoniflorin Against Glutamate-Induced Neurotoxicity in PC12 Cells via Antioxidant Mechanisms and Ca2+ Antagonism. Cell Mol. Neurobiol. 2010, 30, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Tan, Q.R.; Zhang, Z.J. Neuroprotective Effects of peoniflorin, But Not the Isomer Albiflorin, are Associated with the Suppression of Intracellular Calcium and Calcium/Calmodulin Protein Kinase II in PC12 Cells. J. Mol. Neurosci. 2013, 51, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Østerlie, M.; Bjerkeng, B.; Liaaen-Jensen, S. Plasma appearance and distribution of astaxanthin E/Z and R/S isomers in plasma lipoproteins of men after single dose administration of astaxanthin. J. Nutr. Biochem. 2000, 11, 482–490. [Google Scholar] [CrossRef]
- Unlu, N.Z.; Bohn, T.; Francis, D.; Clinton, S.K.; Schwartz, S.J. Carotenoid Absorption in Humans Consuming Tomato Sauces Obtained from Tangerine or High-β-Carotene Varieties of Tomatoes. J. Agric. Food. Chem. 2007, 55, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Schweiggert, R.M.; Kopec, R.E.; Villalobos-Gutierrez, M.G.; Hogel, J.; Quesada, S.; Esquivel, P.; Schwartz, S.J.; Carle, R. Carotenoids are more bioavailable from papaya than from tomato and carrot in humans: A randomised cross-over study. Br. J. Nutr. 2014, 111, 490–498. [Google Scholar] [CrossRef] [Green Version]
- Riso, P.; Brusamolino, A.; Contino, D.; Martini, D.; Vendrame, S.; DelBo’, C.; Porrini, M. Lycopene absorption in humans after the intake of two different single-dose lycopene formulations. Pharmacol. Res. 2010, 62, 318–321. [Google Scholar] [CrossRef]
- Chryssanthi, G.C.; Lamari, F.N.; Georgakopoulos, C.D.; Cordopatis, P. A new validated SPE-HPLC method for monitoring crocetin in human plasma—Application after saffron tea consumption. J. Pharm. Biomed. Anal. 2011, 55, 563–568. [Google Scholar] [CrossRef]
- Almodóvar, P.; Briskey, D.; Rao, A.; Prodanov, M.; Inarejos-García, A.M. Bioaccessibility and Pharmacokinetics of a Commercial Saffron (Crocus sativus L.) Extract. Hindawi Evid. Based Complementary Altern. Med. 2020, 1–8. [Google Scholar] [CrossRef]
- Schmitt-Hoffmann, A.H.; Roos, B.; Sauer, J.; Schleimer, M.; Kovācs, P.; Stoeckel, K.; Maares, J. Influence of food on the pharmacokinetics of oral alitretinoin (9-cis retinoic acid). Clin. Exp. Dermatol. 2011, 36 (Suppl. 2), 18–23. [Google Scholar] [CrossRef]
- Jäger, W.R.; Buchbauer, G.; Jirovetz, L.; Fritzer, M. Percutaneous absorption of lavender oil from a massage oil. J. Soc. Cosmet. Chem. 1992, 43, 49–54. [Google Scholar]
- Kohlert, C.; Schindler, G.; März, R.W.; Abel, G.; Brinkhaus, B.; Derendorf, H.; Gräfe, E.U.; Veit, M. Systemic availability and pharmacokinetics of thymol in humans. J. Clin. Pharmacol. 2002, 42, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Sadakane, C.; Watanabe, J.; Fukutake, M.; Nisimura, H.; Maemura, K.; Kase, Y.; Kono, T. Pharmacokinetic Profiles of Active Components After Oral Administration of a Kampo Medicine, Shakuyakukanzoto, to Healthy Adult Japanese Volunteers. J. Pharm. Sci. 2015, 104, 3952–3959. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Class of Terpenes | Name of Compound | Plant Source | Structure | Affected Component of Calcium Signaling |
|---|---|---|---|---|
| Tetraterpenes -xantophylles | Astaxanthin | Species in the genus Adonis |  | ↓NMDAR expression ↑RYR2 expression ↓L-VGCC A1D expression ↓VGCC current ↑Calbindin expression ↑Parvalbumin expression ↓calpain activity ↓calcineurin activity ↓glutamate toxicity |
| Tetraterpene β-carotenes | Lycopene | Tomatoes, watermelons, pink grapefruits, apricots, pink guavas |  | ↓Cav2.1 activity ↓Cav2.2 activity ↓glutamate toxicity ↓ PKC activity ↑Ca/ATPase expression |
| β-carotene | Carrots, spinach, pumpkins, papayas, sweet potatoes, winter squash, mangoes, cantaloupes, red peppers |  | ↓CAMKIV activity | |
| Crocin and crocetin | Crocus sativus, Gardenia |  | ↓NMDAR current ↓glutamate toxicity | |
| Diterpenes | Retinoic acid | Mangos, papayas, many of the squashes, carrots, sweet potatoes * |  | L-VGCC expression N-VGCC expression VGCC activity ↑NCX1 expression ↓AMPAR expression |
| Monoterpenes | Linalool | Lauraceae, Citrus fruits, Birch trees |  | ↓↑Ca-dependent K-channel PKC activity ↓calpain |
| Thymol | Thyme, Origanum, Ajowan |  | L-VGCC action T-VGCC action | |
| Paeoniflorin | Paeonia lactiflora |  | ↓glutamate toxicity ↑calbindin expression ↓CaMKII activity |
| Single Dose | Peak Level * | Time to Reach Peak Level | Elimination Half Time | Effective Concentration/dose ** | Target | |
|---|---|---|---|---|---|---|
| Astaxanthin | 100 mg | 1.3 µg/mL (2.17 µM) # | 7 h | 21 ± 11 h | 10–20 nM 100 nM 20 nM 0.5–10 µM | NMDAR subunits RAR VGCC Ca2+ level |
| Lycopene from tomato sauces | 4 mg or 13 mg | 30 nM or 0.2 µM | 5 h | > 6 h | 0.1–5 µM 0.5–10 µM 5 mg/kg ** | Cav2.2, Cav2.1 Glutamate excitotoxicity Ca2+-ATPase |
| Lycopene from papaya | 13 mg | 20–30 nM | 6 h | > 6 h | ||
| Lycopene supplement | 15 mg | 40 nM | 10 h | Lack of data | ||
| β-carotene from tomato sauces | 17 mg | 70 nM | 5 h | > 6 h | 18.6–186 µM | CaMKIV |
| β-carotene from papaya | 13 mg | 17–28 nM | 6 h | > 6 h | ||
| Crocetin | Tea from 200 mg saffron | 4 µM | 2 h | Lack of data | 1–50 µM | NMDAR |
| Crocetin-supplement | 25 µg | 400 ng/mL (1.2 µM) # | 1.5 h | 90 min | ||
| Retinoic acid-supplement | 40 mg | 83 ng/mL (0.28 µM) # | 3 h | 6 h | 10 µM | VGCC |
| Linalool-transdermally | 100–150 ng/mL (0.65–0.97 µM) # | 0.3 h | 1 h | 0.1–0.4 mM | K+ current | |
| Thymol-supplement | 1.08 mg | 93 ng/mL (0,62 µM) # | 1.9 h | 10 h | 1 mM | VGCC |
| Paeoniflorin | 5 mg | 6 ng/mL (12.5 nM) # | 3 h | 1.73 | 1–50 mM or 100 µM | ↓ Ca2+i Or CaMKII |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rzajew, J.; Radzik, T.; Rebas, E. Calcium-Involved Action of Phytochemicals: Carotenoids and Monoterpenes in the Brain. Int. J. Mol. Sci. 2020, 21, 1428. https://doi.org/10.3390/ijms21041428
Rzajew J, Radzik T, Rebas E. Calcium-Involved Action of Phytochemicals: Carotenoids and Monoterpenes in the Brain. International Journal of Molecular Sciences. 2020; 21(4):1428. https://doi.org/10.3390/ijms21041428
Chicago/Turabian StyleRzajew, Jowita, Tomasz Radzik, and Elzbieta Rebas. 2020. "Calcium-Involved Action of Phytochemicals: Carotenoids and Monoterpenes in the Brain" International Journal of Molecular Sciences 21, no. 4: 1428. https://doi.org/10.3390/ijms21041428
APA StyleRzajew, J., Radzik, T., & Rebas, E. (2020). Calcium-Involved Action of Phytochemicals: Carotenoids and Monoterpenes in the Brain. International Journal of Molecular Sciences, 21(4), 1428. https://doi.org/10.3390/ijms21041428