Assessing the Effects of Redox Modifier MnTnBuOE-2-PyP 5+ on Cognition and Hippocampal Physiology Following Doxorubicin, Cyclophosphamide, and Paclitaxel Treatment

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Morris Water Maze (MWM)
2.1.1. Distance Moved
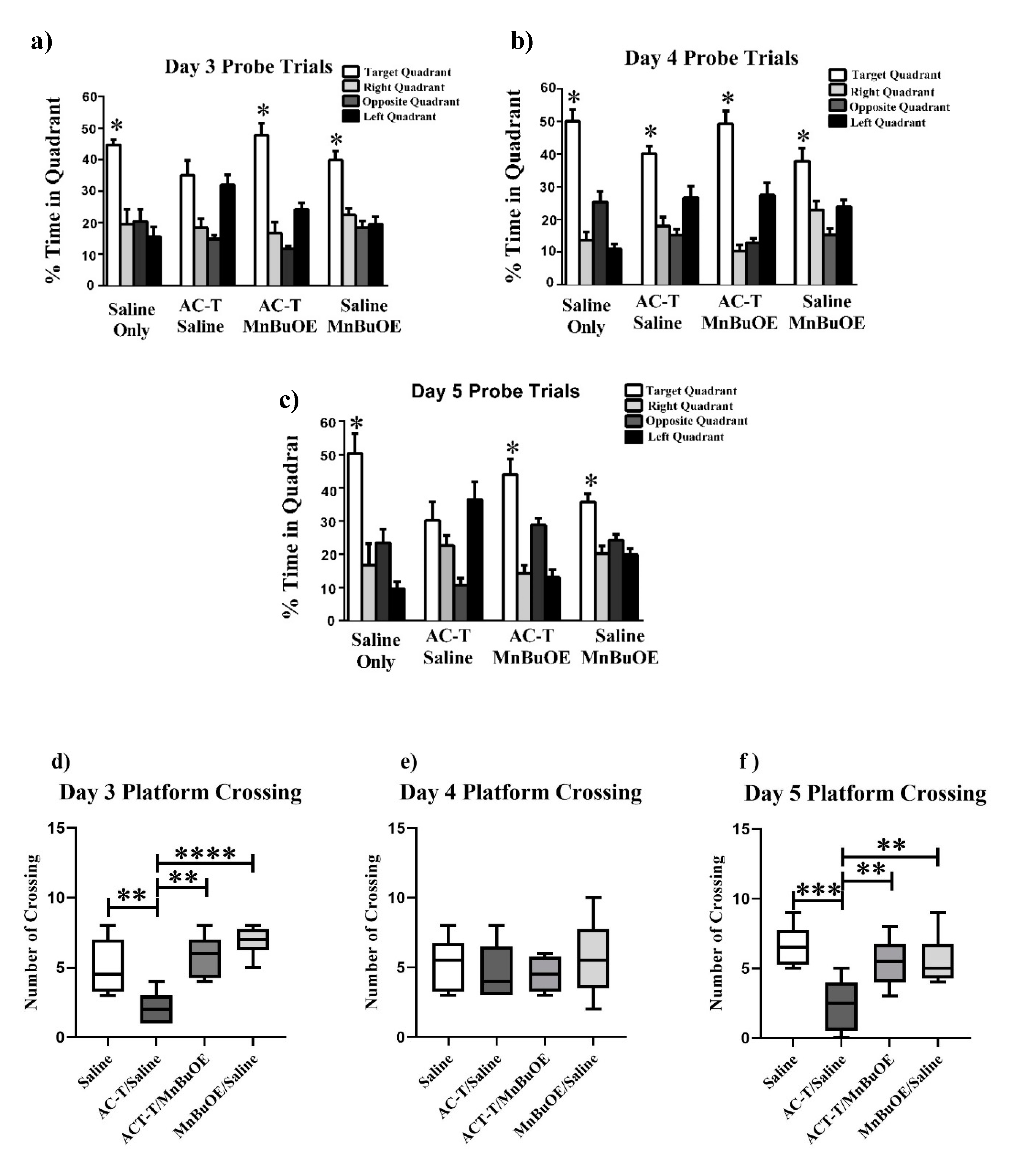
2.1.2. Probe Trials
2.2. Dendritic Morphology
2.2.1. Dentate Gyrus (DG)
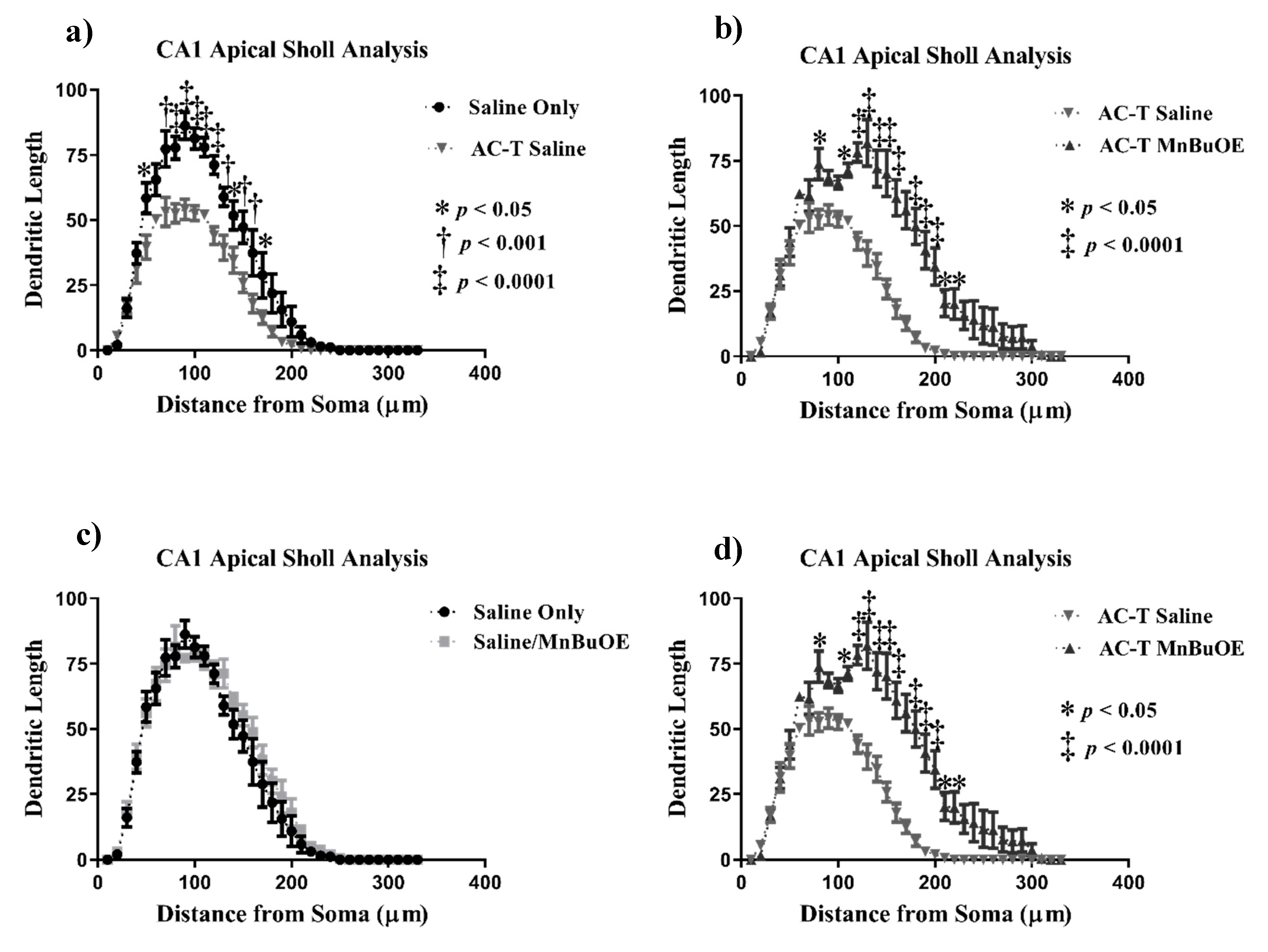
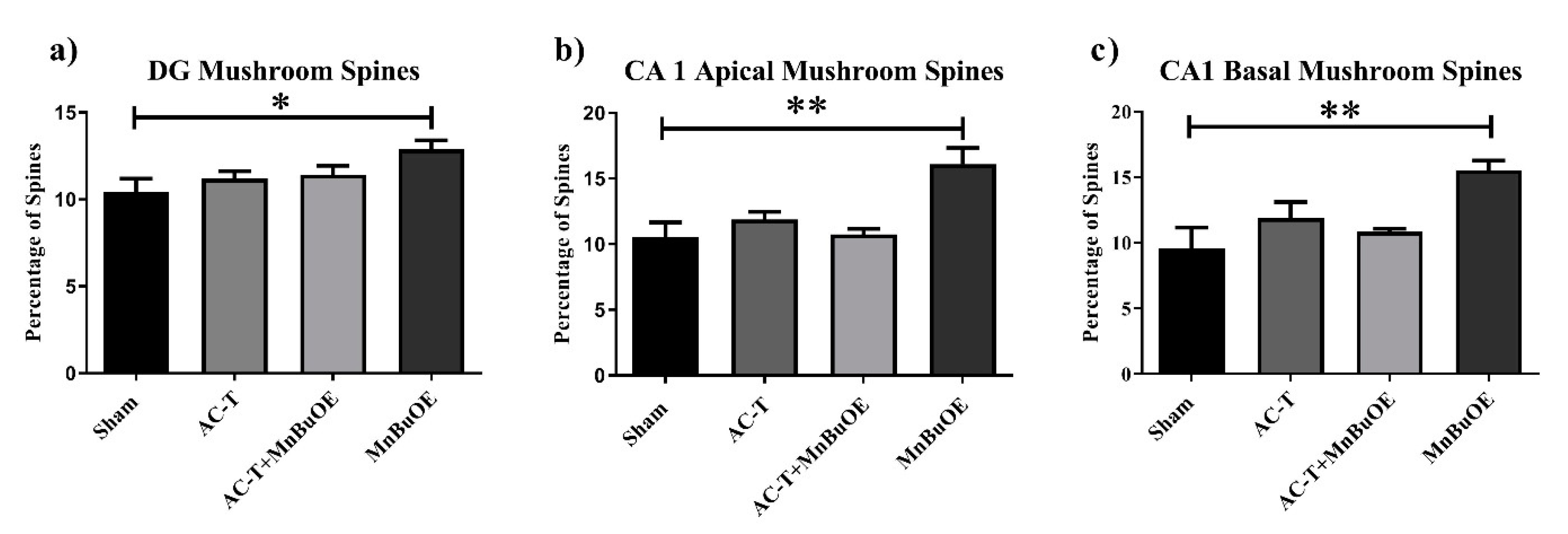
2.2.2. CA1 Pyramidal Neurons
2.3. Proteomics
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemotherapy Paradigm
4.3. Water Maze
4.4. Tissue Collection
4.5. Golgi Staining
4.6. Dendritic Morphology
4.7. Dendritic Spine Morphology
4.8. Proteomics
4.8.1. Tissue Preparation
4.8.2. GeLC-MS/MS Analysis
4.8.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| TLA | Three letter acronym |
| LD | linear dichroism |
| AC-T | doxorubicin, cyclophosphamide, and paclitaxel |
| CA 1 | Cornu Ammonis 1 |
| DG | Dentate Gyrus |
| CICI | Chemotherapy Induced Cognitive Impairment |
| BC | Breast Cancer |
| ROS | Reactive Oxygen Species |
| BBB | Blood Brain Barrier |
| TNFα | tumor necrosis factor alpha |
| NOS | nitric oxide synthase |
| IPA | Ingenuity Pathway Analysis |
| NMDA | N-methyl-D-aspartate |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
References
- Silberfarb, P.M. Chemotherapy and cognitive defects in cancer patients. Annu. Rev. Med. 1983, 34, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Davis, B.D.; Fernandez, F.; Adams, F.; Holmes, V.; Levy, J.K.; Lewis, D.; Neidhart, J. Diagnosis of dementia in cancer patients. Cognitive impairment in these patients can go unrecognized. Psychosomatics 1987, 28, 175–179. [Google Scholar] [CrossRef]
- Argyriou, A.A.; Assimakopoulos, K.; Iconomou, G.; Giannakopoulou, F.; Kalofonos, H.P. Either called “chemobrain” or “chemofog,” the long-term chemotherapy-induced cognitive decline in cancer survivors is real. J. Pain Symptom Manage. 2011, 41, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Hede, K. Chemobrain is real but may need new name. J. Natl. Cancer Inst. 2008, 100, 162–163. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA A Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.S.U.; Ansari, J.; Spooner, D.; Hussain, S.A. Chemotherapy for breast cancer (Review). Oncol. Rep. 2010, 24, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Coombes, R.C.; Bliss, J.M.; Wils, J.; Morvan, F.; Espié, M.; Amadori, D.; Gambrosier, P.; Richards, M.; Aapro, M.; Villar-Grimalt, A.; et al. Adjuvant cyclophosphamide, methotrexate, and fluorouracil versus fluorouracil, epirubicin, and cyclophosphamide chemotherapy in premenopausal women with axillary node-positive operable breast cancer: Results of a randomized trial. The International Collaborative Cancer Group. J. Clin. Oncol. 1996, 14, 35–45. [Google Scholar] [CrossRef]
- Henderson, I.C.; Berry, D.A.; Demetri, G.D.; Cirrincione, C.T.; Goldstein, L.J.; Martino, S.; Ingle, J.N.; Cooper, M.R.; Hayes, D.F.; Tkaczuk, K.H.; et al. Improved outcomes from adding sequential Paclitaxel but not from escalating Doxorubicin dose in an adjuvant chemotherapy regimen for patients with node-positive primary breast cancer. J. Clin. Oncol. 2003, 21, 976–983. [Google Scholar] [CrossRef]
- Chen, Y.; Jungsuwadee, P.; Vore, M.; Butterfield, D.A.; St Clair, D.K. Collateral damage in cancer chemotherapy: Oxidative stress in nontargeted tissues. Mol. Interv. 2007, 7, 147–156. [Google Scholar] [CrossRef]
- Daneman, R.; Prat, A. The Blood–Brain Barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gawad, H.M.A.; Abd El-Gawad, H.M.; El-Sawalhi, M.M. Nitric oxide and oxidative stress in brain and heart of normal rats treated with doxorubicin: Role of aminoguanidine. J. Biochem. Mol. Toxicol. 2004, 18, 69–77. [Google Scholar] [CrossRef]
- Joshi, P.; Vig, P.J.S.; Veerisetty, V.; Cameron, J.A.; Sekhon, B.S.; Desaiah, D. Increase in Brain Nitric Oxide Synthase Activity in Daunorubicin-treated Rats. Pharmacol. Toxicol. 1996, 78, 99–103. [Google Scholar] [CrossRef]
- Oboh, G.; Akomolafe, T.L.; Adefegha, S.A.; Adetuyi, A.O. Inhibition of cyclophosphamide-induced oxidative stress in rat brain by polar and non-polar extracts of Annatto (Bixa orellana) seeds. Exp. Toxicol. Pathol. 2011, 63, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Nafees, S.; Rashid, S.; Ali, N.; Hasan, S.K.; Sultana, S. Rutin ameliorates cyclophosphamide induced oxidative stress and inflammation in Wistar rats: Role of NFκB/MAPK pathway. Chem. Biol. Interact. 2015, 231, 98–107. [Google Scholar] [CrossRef]
- Ince, S.; Kucukkurt, I.; Demirel, H.H.; Acaroz, D.A.; Akbel, E.; Cigerci, I.H. Protective effects of boron on cyclophosphamide induced lipid peroxidation and genotoxicity in rats. Chemosphere 2014, 108, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Mapuskar, K.A.; Anderson, C.M.; Spitz, D.R.; Batinic-Haberle, I.; Allen, B.G.E.; Oberley-Deegan, R. Utilizing Superoxide Dismutase Mimetics to Enhance Radiation Therapy Response While Protecting Normal Tissues. Semin. Radiat. Oncol. 2019, 29, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Spasojević, I.; Chen, Y.; Noel, T.J.; Fan, P.; Zhang, L.; Rebouças, J.S.; St Clair, D.K.; Batinić-Haberle, I. Pharmacokinetics of the potent redox-modulating manganese porphyrin, MnTE-2-PyP(5+), in plasma and major organs of B6C3F1 mice. Free Radic. Biol. Med. 2008, 45, 943–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajic, Z.; Tovmasyan, A.; Spasojevic, I.; Sheng, H.; Lu, M.; Li, A.M.; Gralla, E.B.; Warner, D.S.; Benov, L.; Batinic-Haberle, I. A new SOD mimic, Mn(III) ortho N-butoxyethylpyridylporphyrin, combines superb potency and lipophilicity with low toxicity. Free Radic. Biol. Med. 2012, 52, 1828–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batinic-Haberle, I.; Tovmasyan, A.; Spasojevic, I. An educational overview of the chemistry, biochemistry and therapeutic aspects of Mn porphyrins – From superoxide dismutation to H2O2-driven pathways. Redox Biol. 2015, 5, 43–65. [Google Scholar] [CrossRef] [Green Version]
- Kosmacek, E.A.; Chatterjee, A.; Tong, Q.; Lin, C.; Oberley-Deegan, R.E. MnTnBuOE-2-PyP protects normal colorectal fibroblasts from radiation damage and simultaneously enhances radio/chemotherapeutic killing of colorectal cancer cells. Oncotarget 2016, 7, 34532–34545. [Google Scholar] [CrossRef] [PubMed]
- Alexander, T.C.; Kiffer, F.; Groves, T.; Anderson, J.; Wang, J.; Hayar, A.; Chen, M.T.; Rodriguez, A.; Allen, A.R. Effects of thioTEPA chemotherapy on cognition and motor coordination. Synapse 2019, 73, e22085. [Google Scholar] [CrossRef] [PubMed]
- Groves, T.R.; Farris, R.; Anderson, J.E.; Alexander, T.C.; Kiffer, F.; Carter, G.; Wang, J.; Boerma, M.; Allen, A.R. 5-Fluorouracil chemotherapy upregulates cytokines and alters hippocampal dendritic complexity in aged mice. Behav. Brain Res. 2017, 316, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichenbaum, H. The hippocampus and declarative memory: Cognitive mechanisms and neural codes. Behav. Brain Res. 2001, 127, 199–207. [Google Scholar] [CrossRef]
- Bird, C.M.; Burgess, N. The hippocampus and memory: Insights from spatial processing. Nat. Rev. Neurosci. 2008, 9, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Philpot, R.M.; Ficken, M.; Wecker, L. Doxorubicin and cyclophosphamide lead to long-lasting impairment of spatial memory in female, but not male mice. Behav. Brain Res. 2016, 307, 165–175. [Google Scholar] [CrossRef]
- El-Agamy, S.E.; Abdel-Aziz, A.K.; Wahdan, S.; Esmat, A.; Azab, S.S. Astaxanthin Ameliorates Doxorubicin-Induced Cognitive Impairment (Chemobrain) in Experimental Rat Model: Impact on Oxidative, Inflammatory, and Apoptotic Machineries. Mol. Neurobiol. 2018, 55, 5727–5740. [Google Scholar] [CrossRef]
- Kang, S.; Lee, S.; Kim, J.; Kim, J.-C.; Kim, S.-H.; Son, Y.; Shin, T.; Youn, B.; Kim, J.-S.; Wang, H.; et al. Chronic Treatment with Combined Chemotherapeutic Agents Affects Hippocampal Micromorphometry and Function in Mice, Independently of Neuroinflammation. Exp. Neurobiol. 2018, 27, 419–436. [Google Scholar] [CrossRef]
- Rochefort, N.L.; Konnerth, A. Dendritic spines: From structure to in vivo function. EMBO Rep. 2012, 13, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Bellot, A.; Guivernau, B.; Tajes, M.; Bosch-Morató, M.; Valls-Comamala, V.; Muñoz, F.J. The structure and function of actin cytoskeleton in mature glutamatergic dendritic spines. Brain Res. 2014, 1573, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bourne, J.; Harris, K.M. Do thin spines learn to be mushroom spines that remember? Curr. Opin. Neurobiol. 2007, 17, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Okamoto, K.-I.; Hayashi, Y.; Sheng, M. The importance of dendritic mitochondria in the morphogenesis and plasticity of spines and synapses. Cell 2004, 119, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Berry, M.J. Selenium and selenoproteins in the brain and brain diseases. J. Neurochem. 2004, 86, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Byrns, C.N.; Pitts, M.W.; Gilman, C.A.; Hashimoto, A.C.; Berry, M.J. Mice lacking selenoprotein P and selenocysteine lyase exhibit severe neurological dysfunction, neurodegeneration, and audiogenic seizures. J. Biol. Chem. 2014, 289, 9662–9674. [Google Scholar] [CrossRef] [Green Version]
- Nystuen, A.M.; Yang, A.W. The inositol metabolism pathway as a target for neuroprotective strategies. Neural Regen. Res. 2015, 10, 1928–1929. [Google Scholar] [CrossRef]
- Silvério, R.; Laviano, A.; Rossi Fanelli, F.; Seelaender, M. l-carnitine and cancer cachexia: Clinical and experimental aspects. J. Cachexia Sarcopenia Muscle 2011, 2, 37–44. [Google Scholar] [CrossRef]
- Pin, F.; Barreto, R.; Couch, M.E.; Bonetto, A.; O’Connell, T.M. Cachexia induced by cancer and chemotherapy yield distinct perturbations to energy metabolism. J. Cachexia Sarcopenia Muscle 2019, 10, 140–154. [Google Scholar] [CrossRef] [Green Version]
- Williams, K. Interactions of polyamines with ion channels. Biochem. J. 1997, 326, 943. [Google Scholar] [CrossRef] [Green Version]
- Saunders, L.R.; Verdin, E. Sirtuins: Critical regulators at the crossroads between cancer and aging. Oncogene 2007, 26, 5489–5504. [Google Scholar] [CrossRef] [Green Version]
- Donmez, G. The neurobiology of sirtuins and their role in neurodegeneration. Trends Pharmacol. Sci. 2012, 33, 494–501. [Google Scholar] [CrossRef]
- Falah, M.; Rayan, M.; Rayan, A. A Novel Paclitaxel Conjugate with Higher Efficiency and Lower Toxicity: A New Drug Candidate for Cancer Treatment. Int J. Mol. Sci. 2019, 20, 4965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Allen, A.R.; Eilertson, K.; Sharma, S.; Baure, J.; Allen, B.; Leu, D.; Rosi, S.; Raber, J.; Huang, T.-T.; Fike, J.R. Delayed Administration of Alpha-Difluoromethylornithine Prevents Hippocampus-Dependent Cognitive Impairment after Single and Combined Injury in Mice. Radiat. Res. 2014, 182, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikolaenko, I.; Rao, L.M.; Roberts, R.C.; Kolb, B.; Jinnah, H.A. A Golgi study of neuronal architecture in a genetic mouse model for Lesch–Nyhan disease. Neurobiol. Dis. 2005, 20, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Groves, T.R.; Wang, J.; Boerma, M.; Allen, A.R. Assessment of Hippocampal Dendritic Complexity in Aged Mice Using the Golgi-Cox Method. J. Vis. Exp. 2017. [CrossRef]
- Sholl, D.A. Dendritic organization in the neurons of the visual and motor cortices of the cat. J. Anat. 1953, 87, 387–406. [Google Scholar]
- Kiffer, F.; Alexander, T.; Anderson, J.E.; Groves, T.; Wang, J.; Sridharan, V.; Boerma, M.; Allen, A.R. Late Effects of 16O-Particle Radiation on Female Social and Cognitive Behavior and Hippocampal Physiology. Radiat. Res. 2019, 191, 278. [Google Scholar] [CrossRef]
- Titus, A.D.J.; Shankaranarayana Rao, B.S.; Harsha, H.N.; Ramkumar, K.; Srikumar, B.N.; Singh, S.B.; Chattarji, S.; Raju, T.R. Hypobaric hypoxia-induced dendritic atrophy of hippocampal neurons is associated with cognitive impairment in adult rats. Neuroscience 2007, 145, 265–278. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A Statistical Model for Identifying Proteins by Tandem Mass Spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Description | Location in Cell | Type | Fold Change (log ratio) |
|---|---|---|---|---|
| Q8WUR0 | Protein C19orf12 homolog | ER, Cytosol, Mitochondrion | Transmembrane | –5.53 |
| P97291 | Cadherin-8 | cell membrane | Structural | –5.28 |
| Q544H8 | Zinc finger protein 161, isoform CRA_a | cytosol, nucleus, aggresome | Regulatory | –5.16 |
| Q9CPU2 | NADH dehydrogenase (ubiquinone) 1 beta subcomplex subunit 2 | mitochondrion | Accessory | –4.96 |
| F6RBX1 | Target of myb-like protein 2 (Fragment) | NA | Other | –4.79 |
| A0A0A0MQJ8 | Brain mitochondrial carrier protein 1 | Mitochondrion | Transporter | 4.31 |
| Q8BNZ1 | t-SNARE coiled-coil homology domain-containing protein | Golgi apparatus | Membrane | 4.51 |
| A0A140LJ31 | Putative lipoyltransferase 2 (Fragment) | Mitochondrion | Enzyme | 4.80 |
| I4DCY6 | Sigma non-opioid intracellular receptor 1 | ER | Transmembrane | 5.03 |
| Q3UJR8 | Basic transcription factor 3 | Cytosol | Regulatory | 5.34 |
| Protein | Description | Location in Cell | Type | Fold Change (log ratio) |
|---|---|---|---|---|
| Q9WTZ1 | RING-box protein 2 (Rbx2) | Nucleus, cytoplasm | Enzyme | –4.84 |
| Q8WUR0 | Protein C19orf12 homolog | ER, Cytosol, Mitochondrion | Transmembrane | –4.08 |
| Q8BNZ1 | t-SNARE coiled-coil homology domain-containing protein | Golgi apparatus | Membrane | –4.03 |
| Q9D4I9 | RAB23, member RAS oncogene family, isoform CRA_a | Cytoskeleton, cytosol, plasma membrane | Enzyme | –3.92 |
| A0A0R4J0X8 | Rho guanine nucleotide exchange factor (GEF7), isoform CRA_a | Cytoplasm | Enzyme | 4.56 |
| A2BI12 | PC4 and SFRS1-interacting protein | Nucleus | Regulatory | 5.24 |
| Q8BHC4 | Dephospho-CoA kinase domain-containing protein | Mitochondrion | Enzyme | 5.26 |
| Q3U2K2 | Uncharacterized protein (Fragment) | Membrane | Transmembrane | 5.27 |
| Q3U506 | Uncharacterized protein | NA | NA | 5.80 |
| Network Rank | Network Description |
|---|---|
| Network 1 | Associated network functions: neurological disease, cell morphology, cell-to-cell signaling and interaction |
| Number of “focus molecules” contained in the network: 22 | |
| IPA p-score: 49 | |
| Network proteins: ACSL4, APOA4, ARL1, Akt, Ampa Receptor, CACNG2, CDH8, CDK5RAP3, CFAP20, Creb, ERK, ERK1/2, Filamin, INPPL1, ISOC1, Insulin, Jnk, LDL, LRPAP1, MAP3K4, Mapk, NFkB (complex), P38 MAPK, PI4KB, PON2, PSEN1, Pka, RTF1, S100B, SPRED2, SYNGAP1, TCF4, TMCO1, VAMP2, VHL | |
| Network 2 | Associated network functions: cell death and survival, cancer, cell-to-cell signaling and interaction |
| Number of “focus molecules” contained in the network: 16 | |
| IPA p-score: 33 | |
| Network proteins: ARHGAP17, ATP5MD, DUSP15, EGLN3, FANCD2, GPX3, GRB2, LIG1, LRRC40, LSM12, LSM4, LY6D, LYPLAL1, MLLT11, MRPS33, OSM, PDZD11, PIGK, PRAG1, PTEN, PTPN18, QRICH1, RAB30, RIOX1, SEPHS1, SHIP, TNF, TNFRSF10D, UAP1L1, UNC119B, WIPF2, YIF1B, ZBTB25, cerebroside 3-sulfate, glycosylphosphatidylinositol | |
| Network 3 | Associated network functions: behavior, nervous system development and function, cell-to-cell signaling and interaction |
| Number of “focus molecules” contained in the network: 15 | |
| IPA p-score: 30 | |
| Network proteins: ANKS1B, APP, Atp5e, CASC4, CFDP1, COPS7B, Cops2, DCAF5, DCAKD, DYNLT1, ERP44, FBXO21, H2AFJ, ITM2C, KCNA4, KIAA0586, MOCS2, NFYB, PDCD7, PNMA8B, RABL3, RWDD2A, RWDD2B, SCN1A, SRP19, SRP68, SRP9, SUN5, SV2B, TLL1, TMEM189, UBC, UBL3, UQCRHL, WRNIP1 | |
| Network 4 | Associated network functions: cellular development, cellular growth and proliferation, cell cycle |
| Number of “focus molecules” contained in the network: 12 | |
| IPA p-score: 23 | |
| Network proteins: ARG1, ATG101, BOD1L1, CDC7, CDK2, CDK5R1, CHN2, CPNE3, ERBB2, GLCE, GRPEL1, HMGA2, LRRC4, MRPL55, MYL3, NF2, NRBP1, NTNG1, PLAC8, POLR3E, RB1, RBMS3, RMDN1, SAMHD1, SLC25A14, SPP1, TAX1BP3, TGFB1, TMEM94, TP53RK, TSC22D2, TTC9B, VSIG4, ammonia, dTTP | |
| Network 5 | Associated network functions: cell morphology, nervous system development and function, tissue morphology |
| Number of “focus molecules” contained in the network: 10 | |
| IPA p-score: 18 | |
| Network proteins: AIG1, AP5Z1, BARX2, CHL1, Ck2, DBF4, DDX3Y, E2F1, EPGN, ESR1, FHL1, FOS, FUNDC2, FXYD6, GLP1R, Groucho, HOXA7, Hd-neuronal intranuclear inclusions, LEPROTL1, LMNA, Mta, N-cor, NUCKS1, PDCD5, PDLIM3, PSIP1, Rb-E2F transcription repression, SIN3A, SLC25A42, Sin3, TBL1XR1, TLE1, U2SURP, XK, ZFP36 |
| Pathway Name | p-Value | IPA Ratio |
|---|---|---|
| Molybdenum Cofactor Biosynthesis | 0.01348963 | 0.25 |
| Selenocysteine Biosynthesis II (Archaea and Eukaryotes) | 0.01995262 | 0.167 |
| Fatty Acid Activation | 0.04265795 | 0.0769 |
| Molecular Mechanisms of Cancer | 0.04365158 | 0.0102 |
| Superpathway of Inositol Phosphate Compounds | 0.04570882 | 0.0127 |
| Network Rank | Network Description |
|---|---|
| Network 1 | Associated network functions: cardiovascular disease, cardiovascular system development and function, cell morphology |
| Number of “focus molecules” contained in the network: 24 | |
| IPA p-score: 49 | |
| Network proteins: APRT, ARHGEF7, ATE1, ATG2A, Actin, BTF3, CDH8, CPEB2, CPT2, CSNK2A2, Cadherin, Ck2, Collagen type II, DCUN1D2, DUSP15, EIF2B2, ERK1/2, Filamin, Growth hormone, Hif1, INPPL1, ISOC1, N-cor, PCDH8, PDYN, PPTC7, RNF7, S100B, SLC39A7, SPRED2, THTPA, VCL, VHL, estrogen receptor, phosphatase | |
| Network 2 | Associated network functions: cell-to-cell signaling and interaction, nervous system development and function, infectious diseases |
| Number of “focus molecules” contained in the network: 24 | |
| IPA p-score: 49 | |
| Network proteins: 26s Proteasome, ANKS1B, APOA4, Akt, Ampa Receptor, CACNG2, COPS7B, DCAF5, DHRS7, ELMOD1, ERC1, ERP44, HDL, ITPR, IgG1, Ikb, MAP3K4, MHC Class II (complex), MYD88, NFkB (family), OSTF1, PDZD11, PSEN1, PSME1, Pro-inflammatory Cytokine, RBM8A, RILPL1, SERPINA1, SIGMAR1, SMG8, SYNGAP1, TBK1, TMEM263, TOM1L2, Ubiquitin | |
| Network 3 | Associated network functions: cell death and survival, skeletal and muscular disorders, behavior |
| number of “focus molecules” contained in the network: 17 | |
| IPA p-score: 31 | |
| Network proteins: AGA, AK2, Ap1, CDK5RAP3, CTSS, Creb, F Actin, FCGBP, FSH, GAS2L1, GCLM, IgG, Igm, Immunoglobulin, LDL, Lh, MTA3, Mapk, Mek, NFkB (complex), PARP, PARP1, PDCD6, PGAM2, PI3K (complex), PIP4P2, PPAT, PRKAB2, RGS8, RTF1, TCR, TNFAIP8, Vegf, caspase, cytochrome C | |
| Network 4 | Associated network functions: embryonic development, organ development, organismal development |
| Number of “focus molecules” contained in the network: 16 | |
| IPA p-score: 28 | |
| Network proteins: ADAM19, AP3S1, APRT, ATP5MD, Anti-inflammatory Cytokine, BOD1L1, CAMK2, CCDC85A, CDH4, CDH8, CLMN, CNRIP1, CPNE3, CREB1, DSC2, ESR2, FPR2, FUNDC2, GTF2F2, HABP2, IL4, ITPA, ITPK1, IgG4, JUNB, LMAN2, MOCS2, RAB30, RMDN1, SLC25A14, SPINT1, TGFB1, UBL3, YPEL5, ZBTB14 | |
| Network 5 | Associated network functions: RNA post-transcriptional modification, cell cycle, connective tissue disorders |
| Number of “focus molecules” contained in the network: 15 | |
| IPA p-score: 26 | |
| Network proteins: ALDH18A1, APEH, AQR, ARHGAP17, CDCA7L, CFAP300, CMTR1, COG1, CUL3, HNRNPUL2, INTS6, MLH1, MORC2, MRPS7, NTRK1, PABPN1, PAPSS1, POLR3B, PRPF6, PSIP1, RAMAC, RHOT1, RPAP1, SEPHS1, SLC25A23, SLFN11, SMNDC1, THOC2, TPRG1L, TRA2B, U2SURP, WDR90, WRNIP1, XAB2, XPNPEP3 |
| Pathway Name | p-Value | IPA Ratio |
|---|---|---|
| 3-phosphoinositide Degradation | 0.00278 | 0.0323 |
| Mitochondrial L-carnitine Shuttle Pathway | 0.00484 | 0.118 |
| Superpathway of Inositol Phosphate Compounds | 0.00778 | 0.0253 |
| Polyamine Regulation in Colon Cancer | 0.00806 | 0.0909 |
| Sirtuin Signaling Pathway | 0.00965 | 0.0205 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McElroy, T.; Brown, T.; Kiffer, F.; Wang, J.; Byrum, S.D.; Oberley-Deegan, R.E.; Allen, A.R. Assessing the Effects of Redox Modifier MnTnBuOE-2-PyP 5+ on Cognition and Hippocampal Physiology Following Doxorubicin, Cyclophosphamide, and Paclitaxel Treatment. Int. J. Mol. Sci. 2020, 21, 1867. https://doi.org/10.3390/ijms21051867
McElroy T, Brown T, Kiffer F, Wang J, Byrum SD, Oberley-Deegan RE, Allen AR. Assessing the Effects of Redox Modifier MnTnBuOE-2-PyP 5+ on Cognition and Hippocampal Physiology Following Doxorubicin, Cyclophosphamide, and Paclitaxel Treatment. International Journal of Molecular Sciences. 2020; 21(5):1867. https://doi.org/10.3390/ijms21051867
Chicago/Turabian StyleMcElroy, Taylor, Taurean Brown, Fred Kiffer, Jing Wang, Stephanie D. Byrum, Rebecca E. Oberley-Deegan, and Antiño R. Allen. 2020. "Assessing the Effects of Redox Modifier MnTnBuOE-2-PyP 5+ on Cognition and Hippocampal Physiology Following Doxorubicin, Cyclophosphamide, and Paclitaxel Treatment" International Journal of Molecular Sciences 21, no. 5: 1867. https://doi.org/10.3390/ijms21051867
APA StyleMcElroy, T., Brown, T., Kiffer, F., Wang, J., Byrum, S. D., Oberley-Deegan, R. E., & Allen, A. R. (2020). Assessing the Effects of Redox Modifier MnTnBuOE-2-PyP 5+ on Cognition and Hippocampal Physiology Following Doxorubicin, Cyclophosphamide, and Paclitaxel Treatment. International Journal of Molecular Sciences, 21(5), 1867. https://doi.org/10.3390/ijms21051867