Bacteriophage-Insensitive Mutants of Antimicrobial-Resistant Salmonella Enterica are Altered in their Tetracycline Resistance and Virulence in Caco-2 Intestinal Cells

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. General Characterization of S. Agona BIMs
2.2. Lipopolysaccharide Mutations in BIMs
2.3. Antibiotic Sensitivity of BIMs
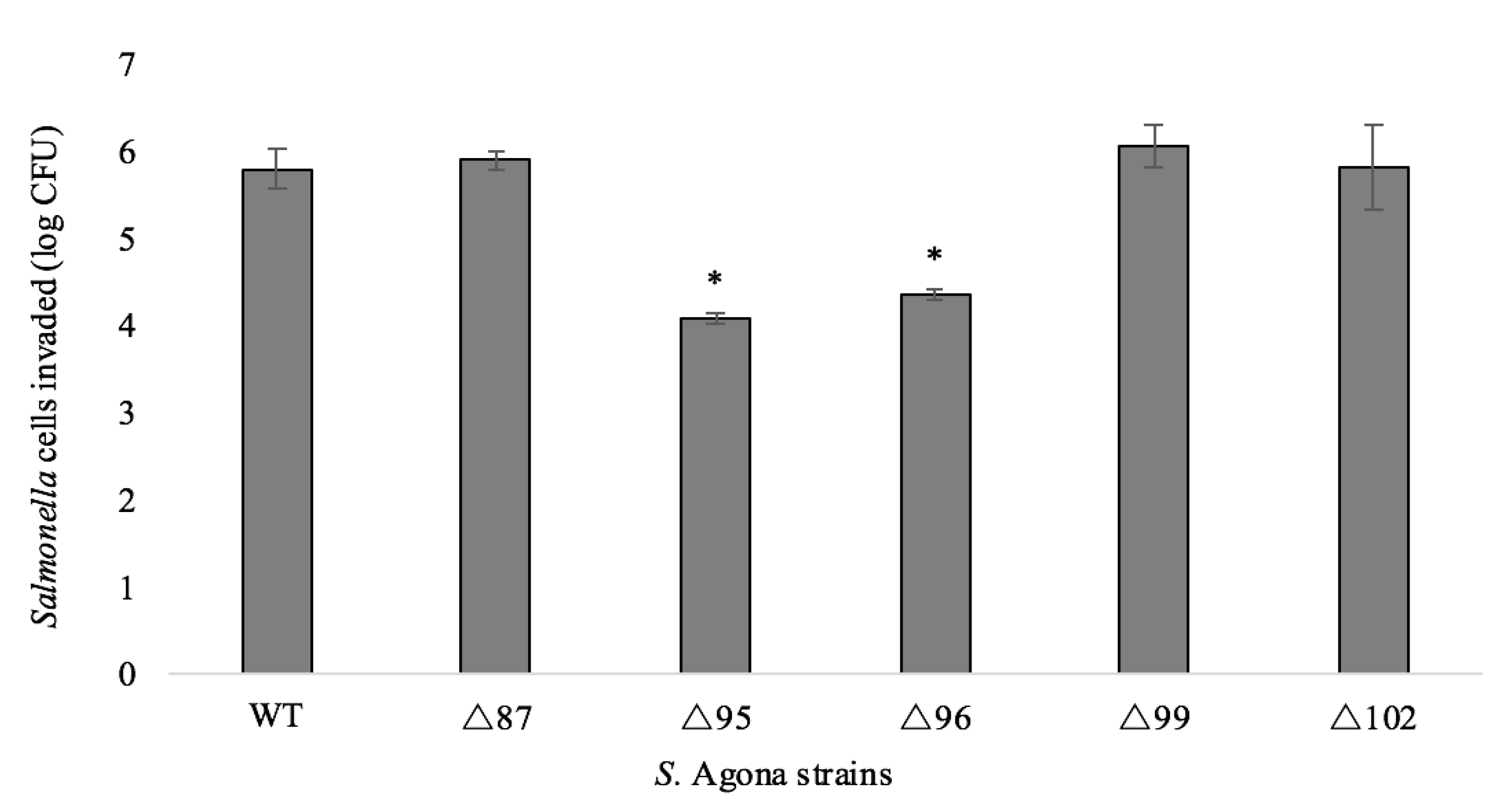
2.4. Adhesion and Invasion Assays
3. Materials and Methods
3.1. Bacterial Maintenance & Growth Conditions
3.2. Isolation of BIMs of S. Agona FSL S5-517
3.3. Minimum-Inhibitory Concentration Assays
3.4. Caco-2 Cell Maintenance and Differentiation
3.5. Adhesion & Gentamicin Protection Assays
3.6. DNA Preparation, Sequencing and Variant Calling
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Majowicz, S.; Musto, J.; Scallan, E.; Angulo, F.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. International Collaboration on Enteric Disease ‘Burden of Illness’ Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, T.T.; Feasey, N.A.; Gordon, M.A.; Keddy, K.H.; Angulo, F.J.; Crump, J.A. Global burden of invasive nontyphoidal Salmonella disease, 2010. Emerg. Infect. Dis. 2015, 21, 941–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, colonization, and virulence differences among serovars. Sci. World J. 2014, 2015, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reports of Selected Salmonella Outbreak Investigations. Available online: https://www.cdc.gov/salmonella/outbreaks.html (accessed on 30 August 2019).
- Fong, K.; LaBossiere, B.; Switt, A.I.M.; Delaquis, P.; Goodridge, L.; Levesque, R.C.; Danyluk, M.D.; Wang, S. Characterization of four novel bacteriophages isolated from British Columbia for control of non-typhoidal Salmonella in vitro and on sprouting alfalfa seeds. Front. Microbiol. 2017, 8, 2193. [Google Scholar] [CrossRef]
- Global Priority List of Antibiotic-Resistant Bacteria to Guide Research Discovery and Development of New Antibiotics. Available online: http://www.who.int/entity/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf? ua=1 (accessed on 2 September 2019).
- Mattila, S.; Ruotsalainen, P.; Jalasvuori, M. On-demand isolation of bacteriophages against drug-resistant bacteria for personalized phage therapy. Front. Microbiol. 2015, 6, 1271. [Google Scholar] [CrossRef] [Green Version]
- Donadio, S.; Maffioli, S.; Monciardini, P.; Sosio, M.; Jabes, D. Antibiotic discovery in the twenty-first century: Current trends and future perspectives. J. Antibiot. 2010, 63, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, P.; Martens, E. Antibiotics in late clinical development. Biochem. Pharmacol. 2017, 133, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Weber-Dabrowska, B.; Jończyk-Matysiak, E.; Zaczek, M.; Łobocka, M.; Łusiak-Szelachowska, M.; Górski, A. Bacteriophage procurement for therapeutic purposes. Front. Microbiol. 2016, 7, 1177. [Google Scholar] [CrossRef]
- Endersen, L.; O’Mahony, J.; Hill, C.; Ross, R.P.; McAuliffe, O.; Coffey, A. Phage therapy in the food industry. Ann. Rev. Food Sci. Technol. 2014, 5, 327–349. [Google Scholar] [CrossRef]
- Filippov, A.A.; Sergueev, K.V.; He, Y.; Huang, X.Z.; Gnade, B.T.; Mueller, A.J.; Nikolich, M.P. Bacteriophage-resistant mutants in Yersinia pestis: Identification of phage receptors and attenuation for mice. PLoS ONE 2011, 6, e25486. [Google Scholar] [CrossRef] [Green Version]
- O’Flynn, G.; Coffey, A.; Fitzgerald, G.; Ross, R.P. Salmonella enterica phage-resistant mutant colonies display an unusual phenotype in the presence of phage Felix 01. Lett. Appl. Microbiol. 2007, 45, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Seed, K.D. Battling phages: How bacteria defend against viral attack. PLoS Pathog. 2015, 11, e1004847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avrani, S.; Wurtzel, O.; Sharon, I.; Sorek, R.; Lindell, D. Genomic island variability facilitates Prochlorococcus-virus coexistence. Nature 2011, 474, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Darby, C. Yersinia pestis is viable with endotoxin composed of only lipid A. J. Bacteriol. 2005, 187, 6599–6600. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef]
- Ferenci, T. Trade-off mechanisms shaping the diversity of bacteria. Trends Microbiol. 2016, 24, 209–223. [Google Scholar] [CrossRef]
- Kim, M.; Kim, S.; Park, B.; Ryu, S. Core lipopolysaccharide-specific phage SSU5 as an auxiliary component of a phage cocktail for Salmonella biocontrol. Appl. Environ. Microbiol. 2014, 80, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Lin, H.; Mi, Z.; Xing, S.; Tong, Y.; Wang, J. Screening of polyvalent phage- resistant Escherichia coli strains based on phage receptor analysis. Front. Microbiol. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Park, M.; Lee, J.H.; Shin, H.; Kim, M.; Choi, J.; Kang, D.H.; Ryu, S. Characterization and comparative genomic analysis of a novel bacteriophage, SFP10, simultaneously inhibiting both Salmonella enterica and Escherichia coli O157: H7. Appl. Environ. Microbiol. 2012, 78, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Lee, J.H.; Kim, H.; Choi, Y.; Heu, S.; Ryu, S. Receptor diversity and host interaction of bacteriophages infecting Salmonella enterica serovar Typhimurium. PLoS ONE 2012, 7, e43392. [Google Scholar] [CrossRef]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS). Available online: http://publications.gc.ca/collections/collection_2018/aspc-phac/HP2-4- 2016-eng.pdf (accessed on 10 September 2019).
- Yang, Z.; Yin, S.; Li, G.; Wang, J.; Huang, G.; Jiang, B.; Peng, Y. Global transcriptomic analysis of the interactions between phage φabp1 and extensively drug-resistant Acinetobacter baumannii. MSystems 2019, 4, e00068-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amram, E.; Mikula, I.; Schnee, C.; Ayling, R.D.; Nicholas, R.A.J.; Rosales, R.S.; Harrus, S.; Lysnyansky, I. 16S rRNA gene mutations associated with decreased susceptibility to tetracycline in Mycoplasma bovis. Antimicrob. Agents Chemother. 2015, 59, 796–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vale, P.F.; Lafforgue, G.; Gatchitch, F.; Gardan, R.; Moineau, S.; Gandon, S. Costs of CRISPR-Cas-mediated resistance in Streptococcus thermophilus. Proc. R. Soc. B 2015, 282, 1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasu, K.; Nagaraja, V. Diverse functions of restriction-modification systems in addition to cellular defense. Microbiol. Mol. Rev. 2013, 77, 53–72. [Google Scholar] [CrossRef] [Green Version]
- Blasdel, B.G.; Ceyssens, P.J.; Chevallereau, A.; Debarbieux, L.; Lavigne, R. Comparative transcriptomics reveals a conserved bacterial adaptive phage response (BAPR) to viral predation. BioRxiv 2018, 2018, 248849. [Google Scholar]
- Sacher, J.; Flint, A.; Butcher, J.; Blasdel, B.; Reynolds, H.; Lavigne, R.; Szymanski, C. Transcriptomic analysis of the Campylobacter jejuni response to T4-like phage NCTC 12673 infection. Viruses 2018, 10, 332. [Google Scholar] [CrossRef] [Green Version]
- Moroni, O.; Kheadr, E.; Boutin, Y.; Lacroix, C.; Fliss, I. Inactivation of adhesion and invasion of food-borne Listeria monocytogenes by bacteriocin-producing Bifidobacterium strains of human origin. Appl. Environ. Microbiol. 2006, 72, 6894–6901. [Google Scholar] [CrossRef] [Green Version]
- Santander, J.; Vasquez, J.I.; Segovia, C.; Santos, L.; Turra, G.; Huber, K. Complete genome sequence of the Salmonella enterica serovar Enteritidis bacteriophages fSE1C and fSE4C isolated from food matrices. Stand. Genome Sci. 2017, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Aballay, A.; Ausubel, F.M. Programmed cell death mediated by ced-3 and ced-4 protects Caenorhabditis elegans from Salmonella Typhimurium-mediated killing. Proc. Natl. Acad. Sci. USA 2001, 98, 2735–2739. [Google Scholar] [CrossRef] [Green Version]
- Hoare, A.; Bittner, M.; Carter, J.; Alvarez, S.; Zaldívar, M.; Bravo, D.; Contreras, I. The outer core lipopolysaccharide of Salmonella enterica serovar Typhi is required for bacterial entry into epithelial cells. Infect. Immun. 2006, 74, 1555–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, M.; Bryan, R.; Rajan, S.; Scheffler, L.; Brunnert, S.; Tang, H.; Prince, A. Role of flagella in pathogenesis of Pseudomonas aeruginosa pulmonary infection. Infect. Immun. 1998, 66, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josenhans, C.; Suerbaum, S. The role of motility as a virulence factor in bacteria. Int. J. Med. Microbiol. 2002, 291, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Laanto, E.; Bamford, J.K.; Laakso, J.; Sundberg, L.R. Phage-driven loss of virulence in a fish pathogenic bacterium. PLoS ONE 2012, 7, e53157. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, Y.; Kita, M.; Kodama, T.; Imamura, S.; Ohno, T.; Sawai, N.; Graham, D.Y. Helicobacter pylori infection in mice: Role of outer membrane proteins in colonization and inflammation. Gastroenterology 2002, 123, 1992–2004. [Google Scholar] [CrossRef]
- Blondel, C.J.; Jiménez, J.C.; Leiva, L.E.; Alvarez, S.A.; Pinto, B.I.; Contreras, F.; Contreras, I. The type VI secretion system encoded in Salmonella pathogenicity island 19 is required for Salmonella enterica serotype Gallinarum survival within infected macrophages. Infect. Immun. 2013, 81, 1207–1220. [Google Scholar] [CrossRef] [Green Version]
- Sana, T.G.; Flaugnatti, N.; Lugo, K.A.; Lam, L.H.; Jacobson, A.; Monack, D.M. Salmonella Typhimurium utilizes a T6SS-mediated antibacterial weapon to establish in the host gut. Proc. Natl. Acad. Sci. USA 2016, 113, e5044–e5051. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, Z.; He, F.; Ruan, Z.; Jiang, Y.; Hua, X.; Yu, Y. The role of the type VI secretion system vgrG gene in the virulence and antimicrobial resistance of Acinetobacter baumanii ATCC 19606. PLoS ONE 2018, 13, e0192288. [Google Scholar]
- Basler, M. Type VI secretion system: Secretion by a contractile nanomachine. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20150021. [Google Scholar] [CrossRef] [Green Version]
- Pukatzki, S.; Ma, A.T.; Sturtevant, D.; Krastins, B.; Sarracino, D.; Nelson, W.C.; Heidelberg, F.; Mekalanos, J.J. Identification of a conserved bacterial protein secretion system in Vibrio cholera using the Dictyostelium host model system. Proc. Natl. Acad. Sci. USA 2006, 103, 1528–1533. [Google Scholar] [CrossRef] [Green Version]
- Schroll, C.; Huang, K.; Ahmed, S.; Kristensen, B.M.; Pors, S.E.; Jelsbak, L.; Lemire, S.; Thomsen, L.E.; Christensen, J.P.; Jensen, P.R.; et al. The SPI-19 encoded type-six secretion-systems (T6SS) of Salmonella enterica serovars Gallinarum and Dublin play different roles during infection. Vet. Microbiol. 2019, 230, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, B.; Dong, T.; Mekalanos, J. A view to a kill: The bacterial type VI secretion system. Cell Host Microbe 2014, 15, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, B.; Zhang, Y.; Wang, X.; Liu, M.; Zhang, T.; Tan, C. Characterization of multiple type-VI secretion system (T6SS) VgrG proteins in the pathogenicity and antibacterial activity of porcine extra-intestinal pathogenic Escherichia Coli. Virulence 2019, 10, 118–132. [Google Scholar] [CrossRef] [Green Version]
- Cascales, E.; Cambillau, C. Structural biology of type VI secretion systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 1102–1111. [Google Scholar] [CrossRef] [Green Version]
- Bingle, L.; Bailey, C.; Pallen, M. Type VI secretion: A beginner’s guide. Curr. Opin. Microbiol. 2008, 11, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Danneels, B.; Pinto-Carbó, M.; Carlier, A. Patterns of nucleotide deletion and insertion inferred from bacterial pseudogenes. Genome Biol. Evol. 2018, 10, 1792–1802. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.E.; Wernegreen, J.J. Sequence context of indel mutations and their effect on protein evolution in a bacterial endosymbiont. Genome Biol. Evol. 2013, 5, 599–605. [Google Scholar] [CrossRef] [Green Version]
- Hosseinidoust, Z.; Van De Ven, T.G.; Tufenkji, N. Evolution of Pseudomonas aeruginosa virulence as a result of phage predation. Appl. Environ. Microbiol. 2013, 79, 6110–6116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sistrom, M.; Park, D.; O’Brien, H.E.; Wang, Z.; Guttman, D.S.; Townsend, J.P.; Turner, P.E. Genomic and gene-expression comparisons among phage-resistant type-IV pilus mutants of Pseudomonas syringae pathovar phaseolicola. PLoS ONE 2015, 10, e0144514. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Cunha, A. Bacteriophages with potential to inactivate Salmonella Typhimurium: Use of single phage suspensions and phage cocktails. Virus Res. 2016, 220, 179–192. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Prot. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Bolton, A.J.; Osborne, M.P.; Stephen, J. Comparative study of the invasiveness of Salmonella serotypes Typhimurium, Choleraesuis and Dublin for Caco-2 cells, HEp-2 cells and rabbit ileal epithelia. J. Med. Microbiol. 2000, 49, 503–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, F.; Ueda, F.; Ochiai, Y.; Mochizuki, M.; Shoji, H.; Ogawa-Goto, K.; Sata, T.; Ogasawara, K.; Fujima, A.; Hondo, R. Invasion assay of Listeria monocytogenes using Vero and Caco-2 cells. J. Microbiol. Methods 2006, 66, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Tritt, A.; Eisen, J.A.; Facciotti, M.T.; Darling, A.E. An integrated pipeline for de novo assembly of microbial genomes. PLoS ONE 2012, 7, e42304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; Van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297. [Google Scholar] [CrossRef] [Green Version]
- Garrison, E.; Marth, G. Haplotype-Based Variant Detection from Short-Read Sequencing. Available online: https://arxiv.org/abs/1207.3907 (accessed on 12 February 2020).
- Den Bakker, H.C.; Allard, M.W.; Bopp, D.; Brown, E.W.; Fontana, J.; Iqbal, Z.; Wolfgang, W.J. Rapid whole-genome sequencing for surveillance of Salmonella enterica serovar Enteritidis. Emerg. Infect. Dis. 2014, 20, 1307–1313. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Affected Gene | Putative Function | Strand | nt Position a | Strain | Mutation | Sequence b | Impact on Polypeptide Synthesis |
|---|---|---|---|---|---|---|---|
| rfaL | O-antigen ligase | + | 425 | WT | 3 bp insertion | G------A | Insertion |
| Δ95 | GGATA | ||||||
| Δ96 | GGATA | ||||||
| + | 429 | WT | Substitution | G | None | ||
| Δ95 | G>A | ||||||
| Δ96 | G>A | ||||||
| + | 442 | WT | 4 bp insertion | T-------A | Frameshift | ||
| Δ95 | TGGGA | ||||||
| Δ96 | TGGGA | ||||||
| rfaY | Lipopolysaccharide core heptose (II) kinase | - | 514 | WT | 23 bp insertion | A---------------------------------------------- A | Frameshift |
| Δ95 | AGCGAAGCCCTAAACTTGTTAAAAA | ||||||
| Δ96 | AGCGAAGCCCTAAACTTGTTAAAAA | ||||||
| - | 538 | WT | Substitution | T | Asn --> Lys | ||
| Δ95 | T>G | ||||||
| Δ96 | T>G | ||||||
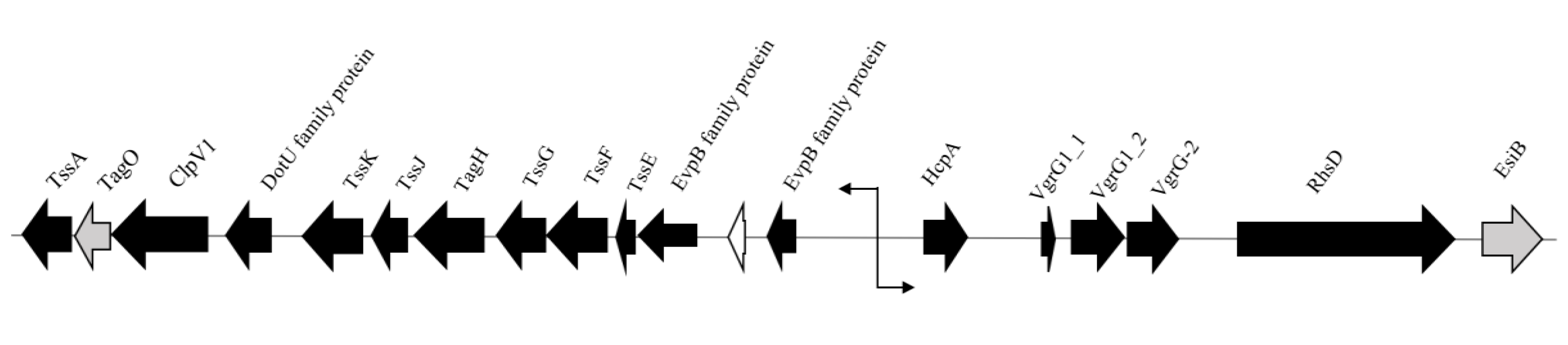
| vgrG1_2 | Actin cross-linking toxin, structural tip protein, involved in type VI secretion | + | 792 | WT | 6 bp insertion | T------------- | Frameshift |
| Δ87 | TCAAGGA | ||||||
| Δ95 | TCAAGGA | ||||||
| Δ96 | TCAAGGA | ||||||
| Δ99 | TCAAGGA | ||||||
| Δ102 | TCAAGGA | ||||||
| + | 798 | WT | Multi-nucleotide substitution | ATTTT | Leu --> Phe Phe --> Gly Tyr --> His | ||
| Δ87 | ATTT>CGGCC | ||||||
| Δ95 | ATTT>CGGCC | ||||||
| Δ96 | ATTT>CGGCC | ||||||
| Δ99 | ATTT>CGGCC | ||||||
| Δ102 | ATTT>CGGCC | ||||||
| + | 810 | WT | 4 bp insertion | A---------T | Frameshift | ||
| Δ95 | AGAACT | ||||||
| Δ96 | AGAACT | ||||||
| + | 816 | WT | 2 bp insertion | T----C | Frameshift | ||
| Δ95 | TCAC | ||||||
| Δ96 | TCAC | ||||||
| + | 820 | WT | Substitution | G | Gly --> Arg | ||
| Δ95 | G>C | ||||||
| Δ96 | G>C | ||||||
| + | 823 | WT | Substitution | G | Gly --> Stop codon | ||
| Δ95 | G>T | ||||||
| Δ96 | G>T | ||||||
| + | 825 | WT | Substitution | A | None | ||
| Δ95 | A>G | ||||||
| Δ96 | A>G | ||||||
| + | 828 | WT | 3 bp deletion | AGGA | Deletion | ||
| Δ95 | A------ | ||||||
| Δ96 | A------ | ||||||
| + | 831 | WT | Substitution | C | Asp --> Glu | ||
| Δ95 | C>G | ||||||
| Δ96 | C>G | ||||||
| + | 836 | WT | Substitution | C | Ala --> Glu | ||
| Δ95 | C>A | ||||||
| Δ96 | C>A |
| Strain | MIC (µg/mL) | ||
|---|---|---|---|
| SXT a | TET | SUF | |
| Wild type | 180/3500 | 128 | 2048 |
| ∆87 | 180/3500 | 128 | 2048 |
| ∆95 | 180/3500 | 128 | 2048 |
| ∆96 | 180/3500 | 76.8 | 2048 |
| ∆99 | 180/3500 | 128 | 2048 |
| ∆102 | 180/3500 | 128 | 2048 |
| Strain | Total Cells Adhered (log CFU ± SD) |
|---|---|
| WT | 6.22 ± 0.23 A |
| ∆87 | 6.65 ± 0.33 A |
| ∆95 | 6.28 ± 0.21 A |
| ∆96 | 6.01 ± 0.19 A |
| ∆99 | 6.48 ± 0.38 A |
| ∆99 | 6.56 ±0.44 A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fong, K.; Mu, K.; Rheault, J.-G.; Levesque, R.C.; Kitts, D.D.; Delaquis, P.; Goodridge, L.; Wang, S. Bacteriophage-Insensitive Mutants of Antimicrobial-Resistant Salmonella Enterica are Altered in their Tetracycline Resistance and Virulence in Caco-2 Intestinal Cells. Int. J. Mol. Sci. 2020, 21, 1883. https://doi.org/10.3390/ijms21051883
Fong K, Mu K, Rheault J-G, Levesque RC, Kitts DD, Delaquis P, Goodridge L, Wang S. Bacteriophage-Insensitive Mutants of Antimicrobial-Resistant Salmonella Enterica are Altered in their Tetracycline Resistance and Virulence in Caco-2 Intestinal Cells. International Journal of Molecular Sciences. 2020; 21(5):1883. https://doi.org/10.3390/ijms21051883
Chicago/Turabian StyleFong, Karen, Kaiwen Mu, Jean-Guillaume Rheault, Roger C. Levesque, David D. Kitts, Pascal Delaquis, Lawrence Goodridge, and Siyun Wang. 2020. "Bacteriophage-Insensitive Mutants of Antimicrobial-Resistant Salmonella Enterica are Altered in their Tetracycline Resistance and Virulence in Caco-2 Intestinal Cells" International Journal of Molecular Sciences 21, no. 5: 1883. https://doi.org/10.3390/ijms21051883
APA StyleFong, K., Mu, K., Rheault, J. -G., Levesque, R. C., Kitts, D. D., Delaquis, P., Goodridge, L., & Wang, S. (2020). Bacteriophage-Insensitive Mutants of Antimicrobial-Resistant Salmonella Enterica are Altered in their Tetracycline Resistance and Virulence in Caco-2 Intestinal Cells. International Journal of Molecular Sciences, 21(5), 1883. https://doi.org/10.3390/ijms21051883